粟酒裂殖酵母中Hsp60蛋白定位與表達水平的研究
2021-02-22 04:01:02王子璇商巾杰
南京師范大學學報(工程技術版) 2021年4期
王子璇,商巾杰
(南京師范大學生命科學學院,江蘇 南京 210023)
蛋白質的組裝被認為是多肽一級序列本身固有特性的唯一結果. 然而,在某些情況下,需要來自其他蛋白質分子的結構信息才能正確折疊并隨后組裝成寡聚物[1]. 這些“輔助”分子被稱為分子伴侶,其中一個亞家族是伴侶蛋白[2],包括10 kDa和60 kDa蛋白質. 分子伴侶在原核生物、葉綠體和線粒體中大量存在,為正常細胞生長所必需,且是應激誘導的,在熱休克條件下起到穩定或保護分解多肽的作用. 其中,Hsp60屬于一類60-65 kDa的分子伴侶,是由14-18個亞基組成的雙圓環形狀蛋白結構[3-4]. 一般來說,細胞中通過轉錄、翻譯后產生的多肽通過與Hsp10復合體相互作用,在Hsp60復合體的空腔中折疊成正確的蛋白結構,這一過程需要ATP的參與[5].
Hsp60屬Ⅰ類分子伴侶,又被稱為HSPD1或Cpn60,存在于線粒體中維持其蛋白質穩態. 其在健康和疾病中發揮著多重作用,尤其是作為一系列獲得性和遺傳性疾病的致病因子[6-8]. 因其有望給分子伴侶疾病的診斷和治療方法帶來新的發展,如各類癌癥、炎癥、自身免疫性疾病及一系列神經退行性疾病[8-11],Hsp60抑制劑和調節劑可能會作為新型抗癌藥物發揮功能[12],近年來國內外學者對Hsp60的研究興趣一直在穩步增長.
粟酒裂殖酵母(Schizosaccharomycespombe)作為模式生物,為分析高等真核生物的基因結構和功能提供了極好的模型系統. 相較于另一種模式生物——芽殖酵母(Saccharomycescerevisiae),裂殖酵母細胞中熱休克蛋白的轉錄調控與哺乳動物細胞中的轉錄調控模式更為相似[13]. 目前對裂殖酵母中熱休克蛋白的研究還不多. 本文對粟酒裂殖酵母中Hsp60蛋白進行了研究,經生物信息學分析研究發現,Hsp60定位于線粒體基質. Hsp60蛋白水平在高溫刺激時顯著增加,而生長時間對其蛋白表達無影響,這也為今后利用Hsp60作為線粒體蛋白表達的參照物提供了理論基礎.
1 材料與方法
1.1 實驗材料
1.1.1 菌株與培養基
粟酒裂殖酵母yHL6381(h+leu1-32his3-D1ura4-D18ade6-M210),由南京師范大學微生物所實驗室保存,培養基為YES(100 mL):3 g葡萄糖,0.5 g酵母粉,20 mg Adenine,20 mgL-histidine,20 mgL-leucine,20 mg Uracil.
1.1.2 實驗試劑
酵母粉購自OXOID;Lysing Enzymes from Trichoderma harzianum購自Sigma-Aldrich;Western Blot 中使用的抗體購自金斯瑞生物科技股份有限公司;其他常用試劑購自南京丁貝生物科技有限公司.
S buffer:sorbitol 25.48 g,HEPES 0.9532 g,1 mol/L MgCl250 μL. 加80 mL超純水完全溶解,用KOH調節pH至6.5,用雙蒸水溶解至100 mL,4 ℃保存.
勻漿buffer:2 mol/L山梨醇 30.0 mL,1 mol/L Tris-HCl(pH 7.5)1.0 mL,0.1 mol/L EDTA 1.0 mL,10 mmol/L PMSF 1.0 mL,用雙蒸水溶解至100 mL.
SEM buffer:100 mmol/L MOPS(pH 7.2)10.0 mL,2.5 mol/L 蔗糖10.0 mL,0.1 mol/L EDTA 1.0 mL,用雙蒸水溶解至100 mL.
Extraction buffer:0.1 mol/L Na2CO3,1 mmol/L PMSF,1 μg/mL pepstatin,用NaOH調pH至11.5.
Solubilization solution:2% SDS,0.8% β-巰基乙醇,20% 甘油,50 mmol/L Tris-HCl(pH 6.8),0.02%溴酚藍.
1.2 實驗方法
1.2.1 線粒體的粗提取
取YES固體培養基上的酵母菌,接入5 mL YES中,30 ℃,200 r/min振蕩培養12 h. 將培養的母液轉接至200 mL YES液體培養基中,調節起始OD600=0.2,30 ℃與37 ℃,200 r/min振蕩培養12 h,收菌. 用超純水和S buffer各清洗一遍菌體,離心收菌. 用S buffer重懸菌體,稱重菌體,加入菌體質量4倍的S buffer和1/10倍溶壁酶Lysing Enzymes from Trichoderma harzianum,37 ℃ 200 r/min 孵育2~4 h,用顯微鏡觀察裂解情況,裂解至80%以上的酵母變圓后離心收菌,加入菌體4倍體積的預冷的勻漿buffer吹打混勻,將菌液轉移至Dounce組織勻漿器中,冰上研磨15-20次. 4 ℃ 4 000 r/min離心5 min,取上清液. 4 ℃ 5 000 r/min離心5 min,取上清液,重復一次. 4 ℃ 12 000 r/min離心5 min,棄上清液. 加5 mL SEM buffer,置于冰上吹打混勻. 4 ℃ 12 000 r/min離心15 min,棄上清液. 沉淀為粗線粒體. 加1 mL SEM buffer,重懸沉淀,-80 ℃保存.
1.2.2 分離線粒體基質蛋白與膜蛋白
取1.2.1中提取的粗線粒體,4 ℃ 12 000 r/min離心15 min,棄上清液. 加入預冷的Extraction buffer重懸線粒體,冰浴30 min,每10 min混勻一次. 冰浴結束后超速離心,4 ℃ 100 000 g離心1 h. 取上清液,4 ℃ 100 000 g再次超速離心1 h. 取上清液,加入等體積20%三氯乙酸溶液,冰上沉降15 min. 4 ℃ 12 000 r/min 離心15 min,棄上清液,用1 mL預冷的丙酮重懸沉淀. 4 ℃ 12 000 r/min離心15 min,棄上清液,于室溫揮發丙酮. 加入Solubilization solution重懸沉淀,此時樣品為可溶性蛋白和與膜結合力弱的蛋白(S). 取第一次超速離心的沉淀,加入預冷的Extraction buffer重懸,4 ℃ 100 000 g超速離心1 h,棄上清液. 加入Solubilization solution重懸沉淀,此時樣品為疏水性膜蛋白(P). 蛋白樣品煮沸后可用于Western Blot檢測,或-80 ℃保存.
1.2.3 Western Blot 檢測蛋白水平
取粗線粒體樣品,加入5×Protein Loading buffer后100 ℃煮沸10 min,上樣于12%聚丙酰胺凝膠進行SDS-PAGE電泳. 300 mA,90 min轉膜. Blocking buffer(0.137 mol/L NaCl,0.02 mol/L Tris,5%脫脂奶粉)封閉NC膜1.5 h. TBST(0.02 mol/L Tris,0.137 mol/L NaCl,0.1% Tween 20,調pH至7.4)振蕩清洗,加入一抗,4 ℃ 12 h或常溫2 h孵育,TBST清洗后,加入二抗,避光孵育1 h,ODYSSEY掃描顯示結果.
2 結果與討論
2.1 Hsp60蛋白結構分析
使用MitoProt II軟件預測Hsp60的線粒體定位序列(mitochondria targeting sequence,MTS),顯示其定位于線粒體信號肽為N端前33個氨基酸,具體序列為MVSFLSSSVSRLPLRIAGRRIPGRFAVPQVRTY,其中包含6個堿性氨基酸和6個羥基氨基酸殘基,但沒有酸性氨基酸殘基. 在Pfam數據庫中,Hsp60蛋白存在一個Chaperonin Cpn60/TCP-1 family結構域,其位置為第54到第558個氨基酸,如圖1所示.
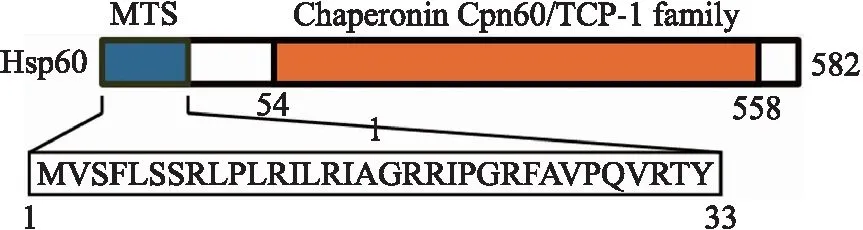
MTS:線粒體定位序列圖1 Hsp60蛋白特征Fig.1 Protein features of Hsp60
使用SWISS-MODEL(http://swissmodel.expasy.org)對Hsp60蛋白質結構進行同源建模,蛋白三維結構模型的模板為HomosapiensHSPD1(SMTL id 4pj1.1),序列相似度為56.90%. Hsp60二級結構主要由α螺旋(紫色)與β折疊(綠色)構成,模型顯示了一種“美式足球”型中間物,有14個亞基,由兩個7亞基的分子伴侶環組成,如圖2所示.

紫色顯示α螺旋結構;綠色顯示β折疊結構圖2 Hsp60蛋白三維結構Fig.2 3D structure of Hsp60
2.2 生物信息學分析
本研究通過NCBI數據庫分析發現了芽殖酵母(Saccharomycescerevisiae)與人類(Homosapiens)的Hsp60的同源基因,并對其蛋白序列進行了生物信息學比對,如圖3所示. 可以發現,在蛋白中段屬于 Chaperonin Cpn60/TCP-1 family結構域中有大量相同序列與保守序列(黃色、藍色),而N端的線粒體定位序列部分有較大差異. 其中,Hsp60與人和芽殖酵母分別有56%或67%的同源性和71%或79%的同源性. 本研究在117—122位氨基酸序列中發現了一個高度保守的ATP結合位點(GDGTTT/S),且在C端發現了一個富含Gly+Met的模體.
2.3 Hsp60蛋白定位
為了證實Hsp60的線粒體定位,首先通過粗提線粒體方法,用Hsp60自身抗體檢測其在不同細胞組分中的表達量. 檢測發現,在線粒體(M)和全細胞蛋白(T)中可檢測得到Hsp60,除線粒體以外的部分(PMS)Hsp60信號極弱,說明Hsp60定位于線粒體而不是細胞質或其他組分,如圖4(a)所示.

黃色表示相同序列;綠色表示微弱相似度序列;藍色表示保守序列圖3 Hsp60同源蛋白序列比對Fig.3 Protein sequence alignment of Hsp60 homologous
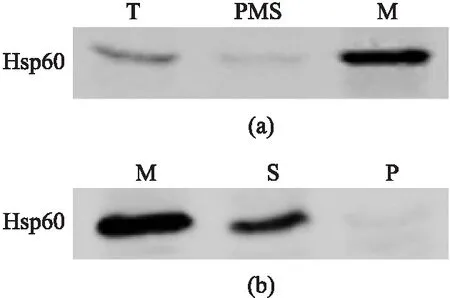
T:全細胞;PMS:上清;M:粗提線粒體;S:上清;P:沉淀圖4 Hsp60定位于線粒體Fig.4 Hsp60 locates in the mitochondria
為進一步探究Hsp60在線粒體中的定位,使用Na2CO3處理粗提線粒體. 使用Na2CO3處理線粒體后,線粒體膜會破碎,釋放出基質蛋白,通過離心方法,即可分離線粒體基質蛋白與膜蛋白. 若蛋白信號在上清(S)中能夠檢測得到,則說明蛋白質定位于線粒體基質;若蛋白信號在沉淀(P)中能夠檢測得到,則說明蛋白質定位于線粒體膜上. 檢測結果如圖4(b)所示,只有在粗提線粒體(M)和上清(S)中可檢測到Hsp60蛋白信號,說明Hsp60定位于線粒體基質而不是線粒體膜上.
2.4 不同生長條件下Hsp60蛋白表達量研究
本文檢測了野生型細胞在30 ℃和37 ℃條件下Hsp60蛋白水平,以及生長對數期和穩定期的蛋白水平,如圖5所示. 細胞在30 ℃和37 ℃培養12 h后,提取線粒體,檢測其Hsp60、Cox4蛋白的表達量,其中Cox4作為對照內參. 可以發現,Hsp60蛋白水平在37 ℃條件下顯著增加(見圖5(a)). 本文還檢測了細胞在正常生長條件下的Hsp60蛋白水平. 細胞在30 ℃培養12 h、60 h后,提取線粒體,檢測其Hsp60、Cox4蛋白的表達量,其中Cox4作為對照內參. 可以發現,在生長對數期和穩定期Hsp60蛋白水平無明顯變化(見圖5(b)).
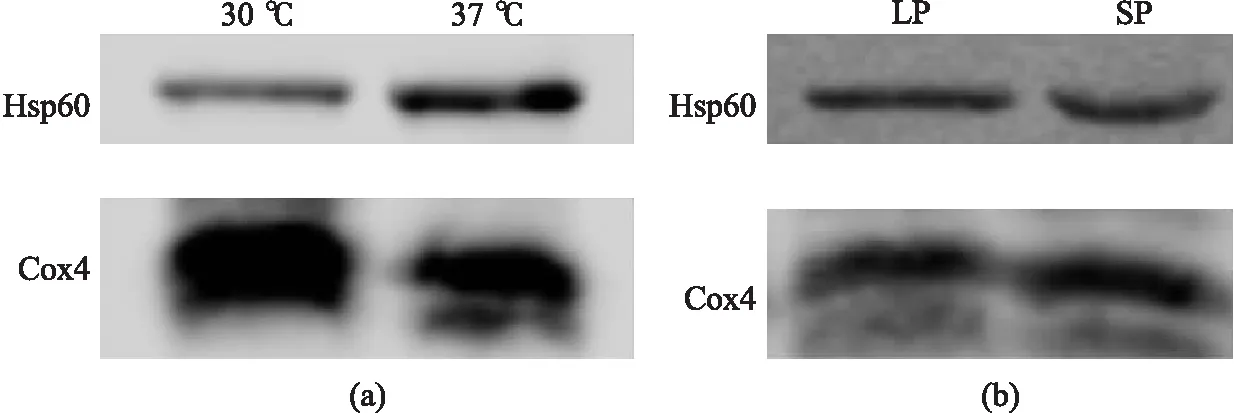
LP:對數期;SP:穩定期圖5 不同生長條件下Hsp60蛋白表達量Fig.5 Hsp60 Protein expression at different temperatures
3 結論
本文經生物信息學研究分析,發現裂殖酵母Hsp60是一類高度保守的蛋白,其中部屬于Chaperonin Cpn60/TCP-1 family結構域的氨基酸序列在人、芽殖酵母中都具有相似性. Chaperonin Cpn60/TCP-1 family結構域在原核生物、真核生物中都十分保守,其蛋白家族包含兩種伴侶蛋白,一種為10 kDa伴侶蛋白(細菌中的Cpn10-或GroES),以6-8個相同亞基的環狀寡聚物形式存在;另一種為60 kDa伴侶蛋白(細菌中的Cpn60-或GroES),由2個堆疊的環組成,每個環包含7個相同的亞基,Hsp60屬于此類伴侶蛋白.
Hsp60在N端含有線粒體定位序列,由細胞核基因編碼,在細胞質中合成后定位于線粒體的蛋白質. 芽殖酵母中的HSP60會定位于線粒體并在線粒體內正確加工為有功能的蛋白質[14]. 人類可以檢測到HSPD1在細胞中定位于線粒體基質[15]. 除此之外,其定位還與細胞狀態密切相關,如誘導細胞凋亡過程中HSPD1會在細胞質內積累[16]. 本文通過提取細胞線粒體及使用Na2CO3處理線粒體,通過Western Blot方法檢測蛋白水平,發現Hsp60定位于線粒體基質.
Hsp60作為一個高度保守的熱激蛋白,在不同的物種中已被廣泛研究. 在芽殖酵母中,當細胞從25 ℃轉移到39 ℃條件下培養時,Hsp60的mRNA水平會升高2~3倍[17]. 而在裂殖酵母中僅有文獻報道,25 ℃培養的細胞轉移到35 ℃培養條件下時,Hsp60的mRNA水平會瞬間升高,然后逐漸降低[18],而蛋白水平的變化還未知. 本文檢測了不同條件下Hsp60的蛋白水平,發現Hsp60在37 ℃熱激條件下,其蛋白表達量顯著增加,之后會輕微降低,這與mRNA水平一致,說明Hsp60在高溫刺激下蛋白表達水平上升,是一個定位于線粒體基質的熱激蛋白.
本研究發現在使用Western Blot方法研究不同生長時期線粒體蛋白水平變化時,Hsp60蛋白表達量保持不變,因此可使用Hsp60與Cox4作為內參蛋白;由于Hsp60在熱激條件下表達量會顯著上升,在研究熱激條件下線粒體蛋白水平變化時應使用Cox4作為內參蛋白,而不能使用Hsp60. 本研究表明,Hsp60可以為研究線粒體蛋白不同生長時期表達量水平提供良好的內參工具.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
