酵母有絲分裂染色體異常分離機制
2021-02-10 07:37:52魏文青謝澤雄
生物技術通報 2021年12期
魏文青 謝澤雄
(天津大學化工學院 系統生物工程教育部重點實驗室 教育部合成生物學前沿科學中心,天津 300072)
有絲分裂過程發生染色體分離錯誤,導致子代細胞增加或減少部分甚至整條染色體的現象稱為非整倍體化[1]。非整倍體是常見的細胞生理現象,存在于植物、動物、真菌等生命體中,影響疾病發生、生物進化、細胞應激等過程[2]。比如常見的唐氏綜合征(21三體綜合征)和愛德華氏綜合癥(18三體綜合征)等非整倍體疾病的發生[3]。非整倍體誘導的基因組不穩定性是驅動惡性腫瘤生長的重要因素。酵母菌被廣泛的應用于生物學、醫學研究和工業生產等過程,在自然界中一般以單倍體或者二倍體形式存在[4-5]。非整倍體酵母細胞通常會表現出多種形式的基因組不穩定性,如細胞內染色體丟失頻率增加,有絲分裂重組事件以及DNA損傷修復缺陷等(圖1)。Deutschbauer等[6]對基因雜合缺失的二倍體釀酒酵母菌株進行適應性分析,以確定細胞中所有導致單倍劑量不足的生長缺陷基因。通過將完整釀酒酵母菌株中測試得到的184個基因進行敲除發現,這些基因處于雜合狀態時細胞的增殖能力會降低。所以非整倍體基因劑量的失衡會顯著降低細胞的適應性,甚至導致細胞死亡[7]。非整倍體釀酒酵母細胞還會出現明顯的G1期延遲,細胞的長度發生變化等現象[8]。因此解析細胞分裂過程中染色體異常分離的機制,對于調控細胞完成正常增殖過程,以及理解和治療非整倍體疾病都是至關重要的。
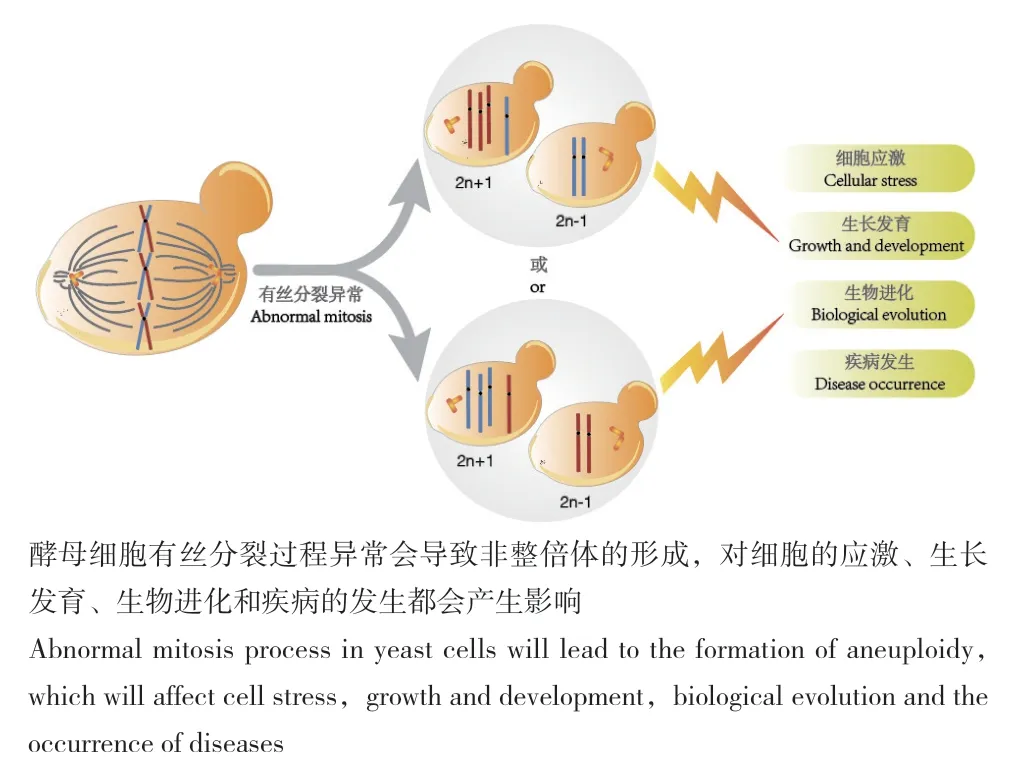
圖1 酵母非整倍體的形成及影響Fig.1 Formation and influence of yeast aneuploidy
酵母主要分為芽殖酵母和裂殖酵母,其中裂殖酵母主要通過有性繁殖和無性繁殖兩種方式進行細胞分裂。通常在營養環境充足的條件下,裂殖酵母主要通過有絲分裂的方式進行一分為二的繁殖[9],芽殖酵母主要通過出芽和接合兩種方式進行繁殖,不同接合型的細胞既可以單獨進行有絲分裂出芽繁殖,也可以與不同接合型細胞進行接合并在一定的條件下開始減數分裂過程[10]。故酵母在不同的繁殖方式中都有可能發生錯誤,形成非整倍體細胞。其中芽殖酵母中比較典型的是釀酒酵母,綜合目前研究發現,釀酒酵母細胞在有絲分裂過程中主要通過“染色體提前分離”和“不分離”兩種方式形成非整倍體[11]。
1 有絲分裂過程染色體提前分離
染色體提前分離是指細胞有絲分裂中期染色體無法正確排列在赤道板上,姐妹染色單體出現提前分離的現象[12-14]。染色體提前分離的原因主要包括姐妹染色單體黏結蛋白缺陷和紡錘體組裝監控機制失效。研究表明這兩種原因都會導致非整倍體子代細胞產生。
1.1 姐妹染色單體黏結蛋白缺陷
黏結蛋白是一種最早在酵母細胞中發現的多亞基復合物,主要由Smc1、Smc3、Scc1和Scc3四個核心亞基組成,在姐妹染色單體的聚合、分離和DNA修復過程中發揮重要的作用[14]。有絲分裂S期形成的姐妹染色單體被黏結蛋白聚合在一起,M期黏結蛋白被酶水解后姐妹染色單體在紡錘體的牽引下向細胞兩極移動,若此過程黏結蛋白發生缺陷則會導致姐妹染色單體因黏聚力缺失而提前分離,隨后紡錘體與姐妹染色單體之間將會發生隨機結合,錯誤的結合將會導致非整倍體細胞的產生[15]。
關于釀酒酵母細胞黏結蛋白如何將姐妹染色單體聚合的模型主要有3類,分別包括單環模型、雙環模型以及聚合桿狀模型。其中單環模型是最常見的模型,即Smc1、Smc3和Scc1/ Scc3形成一個大三角環狀黏結蛋白復合物,能夠環抱捕獲兩個長度約為10 nm的姐妹染色單體[16]。黏結確立蛋白1(Eco1)是一種乙酰轉移酶,是調節黏結蛋白形成的重要因子[17]。Eco1最早在酵母中鑒定成功,在有絲分裂S期促進黏結蛋白亞基乙酰化修飾,形成環狀結構束縛姐妹染色單體[18]。Zhang等[19]通過研究酵母細胞Smc3亞基上,受Eco1乙酰化修飾的兩個賴氨酸位點(K112和K113)突變的影響,實驗發現Eco1對黏結蛋白的修飾作用受到突變的破壞,導致姐妹染色單體黏結蛋白缺陷,子代細胞出現非整倍體現象。
雙環模型,即每個黏結蛋白環都捕獲一個姐妹染色單體。當兩個環產生橋接即Smc1和Smc3形成二聚體環產生內聚力,將姐妹染色單體聚合[20]。同樣,Eco1/Ctf7乙酰轉移酶活性的缺失也會顯著影響雙環模型中黏結蛋白聚合姐妹染色單體的效果[16]。Zhang等[21]利用酵母雙雜交測定法分析黏結蛋白復合物亞基之間的相互作用,實驗發現黏結蛋白復合物中不同亞基之間會出現共同免疫沉淀現象,但是當每個亞基單獨存在時則不顯示相似結果,這證明了雙環模型相比單環模型維持染色單體內聚的靈活性更好。
聚合桿狀模型是指多個Smc1和Smc3組成的異源二聚體之間形成聚合桿狀環繞姐妹染色單體[22]。黏著蛋白與著絲粒特定作用的區域稱為染色體附著區。Surcel等[23]利用微染色體親和純化的方法,從芽殖酵母中分離出含有染色體附著區的微染色體,并利用透射電鏡拍攝和生化分析發現黏結蛋白是桿狀的,且與姐妹染色單體相互作用。實驗還發現DNA的斷裂會直接影響聚合桿狀模型與微染色體的結合,說明黏結蛋白與姐妹染色單體之間的聯系是拓撲結構而不是簡單的物理連接。因此黏結蛋白能夠行使正常活性功能的前提是保證黏結蛋白亞基的乙酰化修飾過程正常。未來關于黏結蛋白的研究還可以通過拓展新的方法,探尋并可視化黏結蛋白聚合姐妹染色單體的實際構象。
1.2 紡錘體組裝監控機制失效
紡錘體組裝檢查點(spindle assembly checkpoint,SAC)最早發現于釀酒酵母細胞中,是一套精確的反饋調節系統[24]。紡錘體組裝監控機制通過監控有絲分裂后期開啟,確保啟動前姐妹染色單體與紡錘體微管連接正確。釀酒酵母細胞中的獨立遺傳學篩選實驗首次發現Mad1、Mad2、Mad3、Bub1和Bub3(紡錘體檢查點動粒蛋白基因)基因參與酵母細胞紡錘體組裝監控通路[25]。后續研究發現紡錘體極復制基因(Msp1)和極光激酶B基因(AuroraB)也是SAC的組成成分[26]。有絲分裂前中期,Mad2基因編碼的蛋白能夠感應細胞中未與紡錘體微管正確連接的動粒發出的信號[27]。促進后期復合體(anaphase promoting complex/cyclosome,APC/C),是指含有多個亞基的E3泛素連接酶,參與調控細胞周期從中期向后期過渡,是SAC激活的關鍵底物[28]。Mad1通過發生磷酸化改變Mad2的構象,促使O-Mad2(Open-Mad2)轉化為具有穩定活性的C-Mad2(Closed-Mad2)構型,并結合APC/C的關鍵激活因子Cdc20形成有絲分裂檢查點復合物(MCC)[29-30]。MCC通過抑制APC/CCdc20的活性延遲酵母細胞有絲分裂后期的開始(圖 2)[31]。
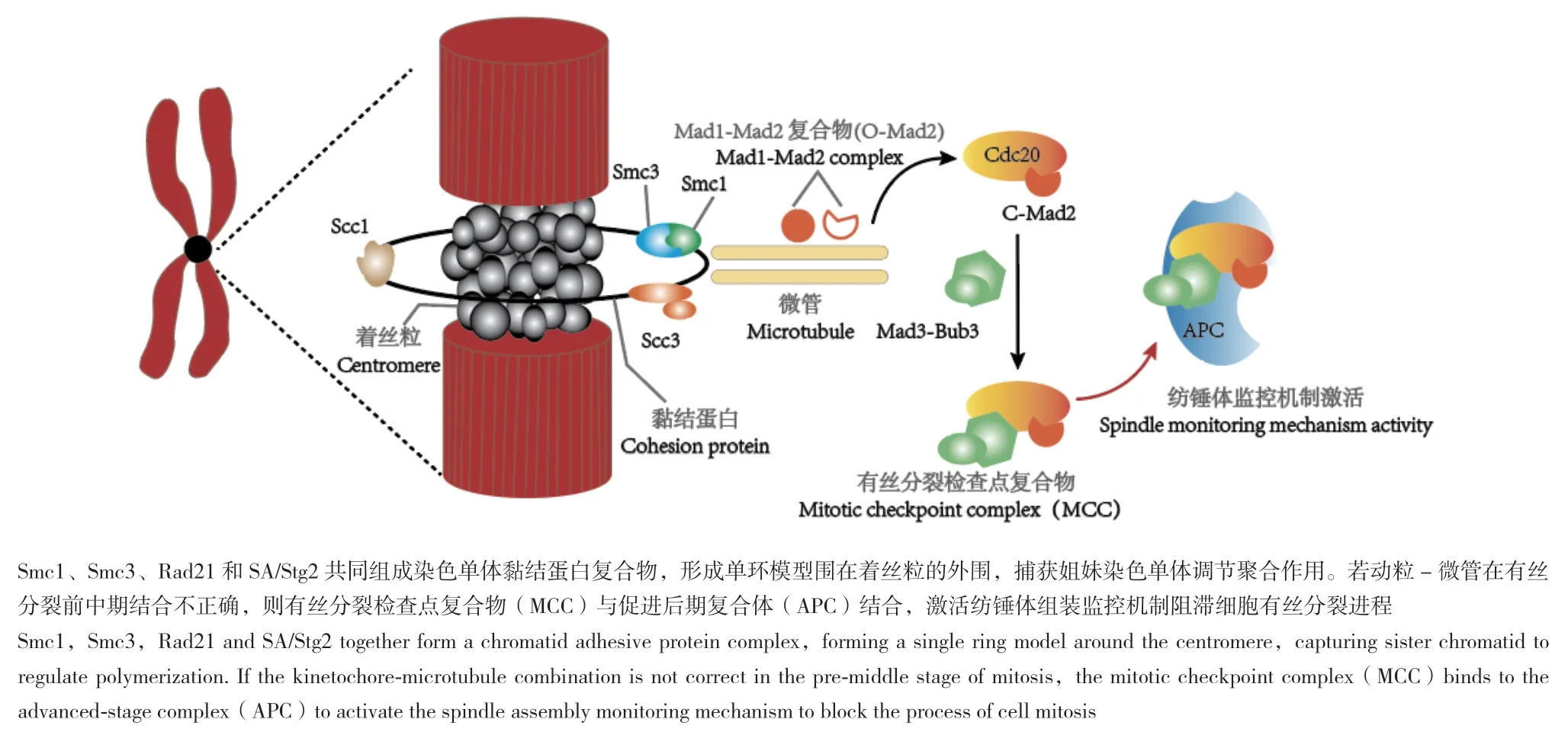
圖2 姐妹染色單體黏結蛋白與紡錘體監控機制Fig.2 Sister chromatid adhesion protein and spindle monitoring mechanism
泛素化是指泛素分子特定連接蛋白Lys位點進行的修飾作用。泛素化修飾過程主要有泛素活化酶E1、泛素結合酶E2和泛素連接酶E3三種酶參與,調節蛋白質的泛素化水平[32]。Reddy[33]等發現紡錘體檢查點的失效是一個耗能的過程,并依賴于APC/C的多泛素化過程。實驗通過在細胞中過表達APC/C,觀察到APC/C發生多泛素化作用會導致Mad2從Cdc20上解離,使紡錘體組裝監控機制失效。
磷酸化修飾是指將ATP的磷酸基團加載到特定蛋白質上的過程,需要磷酸轉移酶的調控作用[34]。Wartrin等[35]發現發生DNA損傷的細胞存活率會顯著降低且染色體變異事件增加,黏結蛋白在進行細胞DNA損傷修復的同時亞基Smc1和Smc3發生磷酸化作用,也會導致SAC功能缺陷。此外,Lopes等[36]發現SAC基因尤其關鍵組分(如Mad2)失活或表達不足也會導致SAC失效,進而產生非整倍體細胞甚至誘導腫瘤的發生。
綜上所述,有絲分裂過程中染色體的正常分離需要功能正常的SAC蛋白維持和調控。若SAC相關亞基的泛素化、磷酸化以及基因表達不足都會導致紡錘體組裝監控機制失效,并對非整倍體現象普遍存在的癌癥的發生具有促進作用[37]。
2 有絲分裂過程染色體不分離
有絲分裂過程染色體不分離是指細胞分裂后期姐妹染色單體無法正常分離,一起進入相同子細胞核中,形成染色體數目不均等的子細胞[38]。總結介紹釀酒酵母有絲分裂染色體不分離的原因主要有兩個,動粒與紡錘體微管連接錯誤和多中心體途徑。其中動粒與紡錘體連接錯誤在酵母細胞中發生的頻率較高,相關的研究結果比較充分。多中心體途徑在一些疾病的發生過程中較為常見。
2.1 動粒-紡錘體微管連接錯誤
動粒是著絲粒上的多亞基蛋白復合物,對于有絲分裂染色體的正確分離起著決定性作用[39]。Dam1復合體位于動粒的外層,是動粒的重要組分之一,在動粒與微管之間起重要的連接作用[40]。動粒在有絲分裂過程中的作用主要包括3個方面,(1)動粒與染色體的運動密切相關,即微管與動粒正確連接且滿足張力時,染色體運動并促進有絲分裂后期進程的開始;(2)紡錘體微管通過與動粒連接捕獲姐妹染色體單體;(3)由動粒通過感知微管連接的張力大小決定紡錘體檢查點的激活或沉默[41]。目前關于釀酒酵母動粒的研究較為深入,許多組成動粒模型的相關蛋白或者亞基都是高度保守的[42]。
有絲分裂早期動粒與紡錘體微管的結合不穩定,可能會發生連接錯誤[43]。酵母細胞動粒與微管錯誤結合的方式主要有3種:Monotelic連接、Syntelic連接和Merotelic連接(圖3-a)。Monotelic連接(單極連接)是指有絲分裂中期一對姐妹染色體單體中只有一條染色體的動粒與一極紡錘體微管連接,而另一條染色單體的動粒沒有被紡錘體微管捕獲連接[44]。Cheeseman等[45]通過在酵母細胞內構建熒光表達蛋白鑒定證明出4個組成Dam1復合體的新亞基,且無法實現磷酸化的Dam1復合體不能及時建立正確的動粒-微管連接,從而導致Monotelic連接在有絲分裂后期出現。
Syntelic連接(同極連接)是指一對姐妹染色單體的動粒都與一側紡錘體微管連接。有絲分裂中期紡錘體組裝監控機制在接收到Monotelic連接、Syntelic連接的信號后,會延遲細胞有絲分裂后期的開始。若紡錘體組裝檢查點的功能失活,有絲分裂后期開始前動粒-微管的錯誤連接則得不到糾正,最終會造成細胞有絲分裂后期一條甚至多條染色體不分離的現象[46]。極光激酶是調控細胞有絲分裂過程正確進行的一類重要絲氨酸激酶,極光酶B在校正酵母細胞動粒-微管連接錯誤事件中發揮重要的作用[47]。缺陷的極光酶B無法感知動粒-微管中產生的張力變化,從而導致細胞中出現Syntelic連接[48-49]。
Merotelic連接(梅洛蒂克連接)是指一對姐妹染色單體中有一個染色體的動粒與兩極紡錘體微管都發生連接,且有絲分裂后期Merotelic連接的染色單體會依據連接兩極微管的多少進行運動定位。Gregan等[50]認為Merotelic連接是生物有絲分裂過程形成非整倍體的主要機制,同時也是促進癌細胞染色體不穩定的主要原因。所以動粒與紡錘體微管之間正確連接的關鍵是動粒能否正常發揮功能,以及感知動粒-微管連接張力變化的極光激酶是否可以正常接收信號。
2.2 多中心體途徑
中心體是主要的微管組織中心,不僅調控微管的成核運動,而且推動著細胞有絲分裂進程[51]。每個細胞周期中心體僅復制一次,復制形成的兩個中心體產生微管并移動到細胞兩極。若細胞不能正確調控中心體的復制將會出現中心體數目異常現象[52-54]。當細胞中中心體數量超過兩個時,便會出現紡錘絲與動粒之間短暫的多極連接,姐妹染色單體因受力不均勻分離形成非整倍體子細胞[55](圖 3-b)。

圖3 動粒-微管結合錯誤和多中心體途徑Fig.3 Miscellaneous kinetochore-microtubule binding and polycentric pathway
多中心體產生的原因主要包括:中心體擴增、中心粒聚合缺陷以及中心體片段化[56]。Polo樣蛋白激酶4(PLK4)是參與調控中心體復制過程的重要組分,當PLK4過表達時中心體復制機制會發生缺陷,將會導致一個細胞周期內中心體發生多次復制[57-58]。Meraldi等[59]進行細胞中心體復制實驗發現,E2F轉錄因子和Cdk2-周期蛋白A活性的激活也是中心體復制必須的。所以這些蛋白或因子的過表達都有可能促使細胞內中心體的擴增。
然而當細胞中某些蛋白表達不足時,中心粒之間的聚合張力會降低甚至消失,導致中心粒聚合發生缺陷[60]。此外,中心體結構的缺陷可能導致片段化的中心粒與微管連接形成極紡錘體[57]。以上3種多中心產生的機制都會導致細胞發生多極分裂,即有絲分裂過程形成3個甚至更多個子細胞核。很多研究表明多極分裂最終導致非整倍體細胞的產生,尤其在多種癌癥的發生中較為明顯[61]。
3 總結與展望
隨著生物學技術的發展,對酵母有絲分裂過程中染色體異常分離機制的研究已經有了顯著的成果。從最初的細胞學觀察到如今深層次的遺傳學研究,發現很多蛋白和調節因子參與有絲分裂染色體分離,并且起到密切的調控作用。但是綜合目前的研究會發現還有很多蛋白或亞基的作用機制未闡釋清晰,如調節動粒-微管連接的Dam1復合體相關亞基。此外,是否還有新的調控蛋白或通路未發現對于研究者而言也是一個挑戰。其次,染色體分離異常直接導致非整倍體的產生。現有的研究都表明非整倍體是癌癥細胞的顯著特征,增加了癌細胞基因組的不穩定性,但是非整倍體與腫瘤發生的關系亟待進一步的研究闡明。最后,在酵母細胞中染色體分裂異常也會導致多倍體細胞的形成,但是多倍體細胞與非整倍體細胞之間的調控關系仍是一個懸而未決的問題。系統且全面的了解和建立不同物種非整倍體形成方式及調控網絡是必要的,縮小研究的成本及時間范疇,探尋和解釋細胞有絲分裂染色體分離異常的原理,并應用其解決與人類息息相關的實際問題是本研究的終極任務。