外界因素和食品內部環境變化對Brevilaterin抗菌特性的影響
2021-02-02 11:34:32王美姿劉楊柳韓盼盼王露露王星星賈英民
食品科學技術學報 2021年1期
王美姿,劉楊柳,韓盼盼,秦 明,王露露,王星星,陳 洲,賈英民
(北京工商大學 食品與健康學院,北京 100048)
食品防腐是保障食品免受微生物污染、延遲保質期的重要舉措,除采用低溫、腌制(高糖或高鹽等)、真空保藏等傳統的防腐方法以外,通常還會添加各類防腐劑以達到協同抗菌的功效[1]。隨著人們對食品安全問題的日益重視,以往常用的化學類防腐劑由于易殘留、過量食用后易導致人體出現健康問題等缺陷已逐步被限制使用[2],一些新型的生物類防腐劑應運而生。抗菌肽(antimicrobial peptides,AMPs)被認為是當下研究最熱、應用前景最好的生物防腐劑之一[3]。AMPs是由宿主產生的能抵御外界微生物侵害、消除體內突變細胞的一類小分子多肽[4],具有抑菌譜廣、抑菌機制獨特且不易殘留等優勢;而且,諸如Nisin等一系列的AMPs物質都已被正式批準并應用在食品防腐當中[5]。
食品是一個復雜多變的混合體系,受組分構成、加工方式以及存儲條件等眾多因素的影響,并由此影響食品添加劑,如防腐劑等的使用[6]:Nisin在酸性食品中能發揮出非常好的抗菌效果,但在中性或堿性條件下卻幾乎無抗菌作用,高溫處理或在含卵磷脂(≥1%)的食品中也同樣會完全失去抗菌活力[7];食品中存在的金屬離子也可能對AMPs的抗菌活性造成影響,如張煒等[8]發現,Fe2+能使抗菌肽BSN-37的活性消失;此外,食品的酸堿性以及表面活性劑等的存在均可能對AMPs的抗菌活性產生影響,如酸處理能顯著降低辣椒籽AMPs[9]的抗菌活性,而吐溫-20和吐溫-80卻能使其抗菌能力提高[10]。可見,AMPs能否適應外界因素和食品內部環境的變化直接制約其在食品防腐領域中的應用。為此,挖掘出適應不同食品的AMPs新資源成為該領域研究者期待解決的關鍵問題之一。
側孢短芽孢桿菌(Brevibacilluslaterosporus)是近年來新發現的生物抗性細菌,能分泌多組分AMPs物質,這些AMPs具有抗細菌、抗真菌、抗癌細胞和抗病毒等生物特性,而且能抵抗溫度、pH值及化學試劑等外界因素對其抗菌活性的影響。本課題組前期已獲得了一株優良的B.laterosporus[11],并發現溫度、pH值、化學試劑以及紫外線處理等外界因素均不會影響該菌分泌的AMPs(Brevilaterin)的抗菌活性,表現出了Brevilaterin良好的應用潛能[12-13]。然而,Brevilaterin是否能抵抗食品中更多外界因素對其抗菌活性的影響還不得而知,這也一定程度上限制了其應用,此外,Brevilaterin在食品發生變化的過程中所表現出的抗菌作用規律也不清楚。本研究將以Brevilaterin為對象,系統研究不同外界因素對其抗菌活性的影響,并考察Brevilaterin在食品內部環境變化過程中的抗菌作用規律,旨在為其在食品防腐中的應用提供充足的理論支撐。
1 材料與方法
1.1 材料與試劑
抗菌肽Brevilaterin,由本實驗室篩選的側孢短芽孢桿菌B.laterosporusS62-9[14]分泌所得,其終效價為861.0 AU/mL。
指示菌:金黃色葡萄球菌(Staphylococcusaureus,CICC No.10001)、單增李斯特氏菌(Listeriamonocytogenes,CICC No.21635)、藤黃微球菌(Micrococcusluteus,CICC No.10269)、銅綠假單胞菌(Pseudomonasaeruginosa,CICC No.10351)、乳酸乳球菌(Lactococcuslactis,CICC No.20711)、嗜堿芽孢桿菌(Halomonasalkalicola,CICC No.11012s)、梭狀芽孢桿菌(Bacillusfusiformis,CICC No.20463),均購自中國工業微生物菌種保藏管理中心。
發酵培養基:葡萄糖15.0 g,魚蛋白胨12 g,CaCl21.38 g,ZnCl20.008 16 g、Tween-20 1 g,蒸餾水1.0 L,pH值7.0。
胰蛋白胨大豆瓊脂(TSA)培養基:胰蛋白胨15.0 g,大豆胨5.0 g,NaCl 5.0 g,瓊脂13.0 g,蒸餾水1.0 L,pH值7.3±0.2。
乳酸菌培養基I(LBMI):酵母膏7.5 g,葡萄糖10.0 g,番茄汁100 mL,蛋白胨7.5 g,KH2PO42.0 g,Tween-80 0.5 g,蒸餾水900 mL,pH值7.0。
MRS肉湯培養基:蛋白胨10 g, 牛肉粉5 g,葡萄糖20 g,酵母粉4 g,乙酸鈉5 g,KH2PO45 g,MgSO40.2 g,檸檬酸三銨2 g,MnSO40.05 g,Tween-80 1 g,蒸餾水 1.0 L,pH 值6.2±0.2。
營養瓊脂(NA)培養基:蛋白胨10 g,牛肉浸粉3 g,NaCl 5 g,瓊脂15 g,蒸餾水1.0 L,pH值7.3±0.1。
1.2 儀器與設備
PREP中壓層析系統,瑞士BUCHI公司;Scan 4000型全自動菌落計數儀,法國Interscience;INFINITE Spark 10M型酶標儀,瑞士Tecan公司;Omega series微生物生長曲線測定儀,德國BMG Labtech公司。
1.3 實驗方法
1.3.1Brevilaterin抗菌活性測定
采用瓊脂擴散法,以S.aureus為指示菌[9]。以經2代活化的菌為種子液發酵制備菌懸液,稀釋菌濃度至OD600為0.7~0.8,按體積分數為3%的量添加到預冷至50 ℃左右的NA培養基中,迅速混勻后倒平板,備用。
取無菌牛津杯擺放在倒好的平板上,加入50 μL Brevilaterin溶液,將平板移至4 ℃冰箱并靜置預擴散1 h,再移至37 ℃恒溫培養箱內靜置培養16 h,待平板上長出致密的菌層,并出現清晰的抑菌圈后取出,用全自動菌落計數儀測量抑菌圈直徑。
1.3.2最小抑菌濃度的測定
最小抑菌濃度(minimum inhibitory concentration,MIC)的測定,采用微量液體稀釋法,參照Kondejewski等[15]的方法并作適當修改。
1)調節菌體濃度。以無菌培養基為空白,測定菌懸液的OD600,調節菌濃度至與0.5麥氏比濁管(McFarland)相同的OD600(約為1×108CFU/mL),再用0.85%無菌生理鹽水稀釋菌濃度至1.0×106CFU/mL,備用。
2)MIC測定。以96板的每行為一個測試組,各設3個平行,1~10列分別為不同濃度的處理組,11列為陽性對照組,12列為空白對照組。向1~11列微孔中分別注入100 μL無菌培養基,第12列加入200 μL無菌培養基。向第1列中加入100 μL質量濃度為0.5 mg/mL的AMPs溶液,再逐級向后進行2倍稀釋至第10列,混勻后舍棄第10列板孔中的100 μL溶液。再分別向1~11列微孔中接入100 μL預調好濃度的菌懸液,使其終濃度為5×105CFU/mL,置于菌株最適生長溫度下培養16~20 h。使用酶標儀檢測600 nm波長處的吸光值(A值),根據A值計算AMPs對菌株的生長抑制率。結合抑制率大于等于90%所對應的濃度,以及肉眼觀察無可見菌生長的最低濃度最終確定MIC。
1.3.3最小殺菌濃度的測定
最小殺菌濃度(minimum bactericidal concentration,MBC)的測定,以MIC測定結果為基礎,分別吸取MIC實驗中無菌生長濃度下的菌懸液100 μL并涂布于固體培養基上,置于該菌最適生長條件下培養24 h,以平皿上菌落數小于5個的最低濃度為MBC值,實驗重復3次。
1.3.4不同外界因素及處理對Brevilaterin抗菌活性影響實驗
1.3.4.1 金屬離子、乳化劑、增稠劑、酶類和蔗糖對Brevilaterin抗菌活性影響的處理方法
考察了不同外界因素對Brevilaterin抗菌活性的影響,包括金屬離子、乳化劑、增稠劑、酶類和高濃度蔗糖。處理方法:分別將金屬離子(Na+、K+、Ca2+、Mg2+、Co2+、Zn2+、Mn2+、Ni2+、Al3+和Fe3+,終濃度為12.5~300.0 mmol/L、乳化劑(甘油、十二烷基苯磺酸鈉、單硬脂酸甘油酯和大豆磷脂,終質量分數為1%)、增稠劑(明膠、羧甲基纖維素鈉、黃原膠和卡拉膠,終質量分數為0.5%)、酶類(脂肪酶、蛋白酶K、胰蛋白酶、胃蛋白酶、復合蛋白酶、溶菌酶、酸性蛋白酶和堿性蛋白酶,終質量濃度為30.0 mg/mL)和蔗糖(終質量分數10%~50%)與Brevilaterin(終質量濃度為0.6 mg/mL)混勻,再按1.3.1的方法測定其抗菌活性。以只添加不同因素的樣品組作為陰性對照組,以只添加Brevilaterin的樣品組作為陽性對照組。
1.3.4.2 反復凍融對Brevilaterin抗菌活性影響的處理方法
將終質量濃度為0.6 mg/mL的Brevilaterin反復凍融(-20 ℃凍結,常溫解凍;0~12次)處理,再按1.3.1的方法測定Brevilaterin的抗菌活性,以不經凍融處理的Brevilaterin為空白對照。
1.3.5食品內部環境變化對Brevilaterin抗菌作用影響實驗
分別考察了食品內部溫度、pH值、滲透壓變化以及氧含量變化對Brevilaterin抗菌作用規律的影響,以MIC和MBC為最終考察指標。
1.3.5.1 溫度變化實驗
以S.aureus、L.monocytogenes、M.luteus、P.aeruginosa為指示菌,按照1.3.2和1.3.3的方法分別測定Brevilaterin在溫度變化條件下(4、10、25、37 ℃)對指示菌的MIC和MBC。
1.3.5.2 pH值變化實驗
以L.lactis、S.aureus和H.alkalicola為指示菌,按照1.3.2和1.3.3的方法分別測定Brevilaterin在pH值變化條件下(pH值 2.0~11.0)對指示菌的MIC和MBC。
1.3.5.3 滲透壓變化實驗
以S.aureus為指示菌,按照1.3.2和1.3.3的方法分別測定Brevilaterin在滲透壓變化條件下(蔗糖質量分數為3%~50%;NaCl質量分數為1%~15%)對S.aureus的MIC和MBC。
1.3.5.4 氧含量變化實驗
以B.fusiformis、S.aureus為指示菌,按照1.3.2和1.3.3的方法分別測定Brevilaterin在氧氣含量變化條件下(5%~21%)對B.fusiformis、S.aureus的MIC和MBC。
2 結果與分析
2.1 不同外界因素及處理條件對Brevilaterin抗菌活性的影響
2.1.1不同外界因素對Brevilaterin抗菌活性的影響
本課題組前期探索發現,溫度、pH值、常見的化學試劑以及紫外線等并不直接影響Brevilaterin的抗菌活性[12-13]。在此基礎上,本研究進一步系統考察了更多外界因素對Brevilaterin抗菌活性的影響,包括金屬離子、乳化劑、增稠劑、酶類和高濃度蔗糖。結果表明,Brevilaterin能夠抵抗除大豆磷脂以外的所有測試因素對其抗菌活性的影響。可見,Brevilaterin具有很好的環境適應性,這也為其在更多類別食品中的抗菌應用提供了良好的理論支撐。
2.1.1.1 金屬離子的影響
某些食品(如皮蛋、油條、鹵水豆腐等)因加工的需要通常會添加較高濃度的金屬離子,如Pb2+、Al3+、Mg2+、Ca2+等。這些金屬離子(添加量在10~300 mmol/L)很可能會對蛋白或肽類物質的穩定性產生影響并造成其活性的變化[1,10,16],如金屬離子會抑制魚精蛋白的抑菌活力,并隨著電荷數和濃度的增加而增強[1]。李軍等[16]發現,90.0 mmol/L Ag+使蘿卜籽蛋白提取物的抗菌活性降低70%,而300.0 mmol/L的Ca2+可以使花椒籽蛋白AMPs的活性提高[10]。還有部分肽類抗生素要發揮抗菌作用還需依賴金屬離子的存在,這也為其應用帶來一定的麻煩,如桿菌肽和博來霉素就是典型的金屬依賴類抗菌肽[17]。金屬離子對Brevilaterin抗菌活性研究的實驗結果見表1。
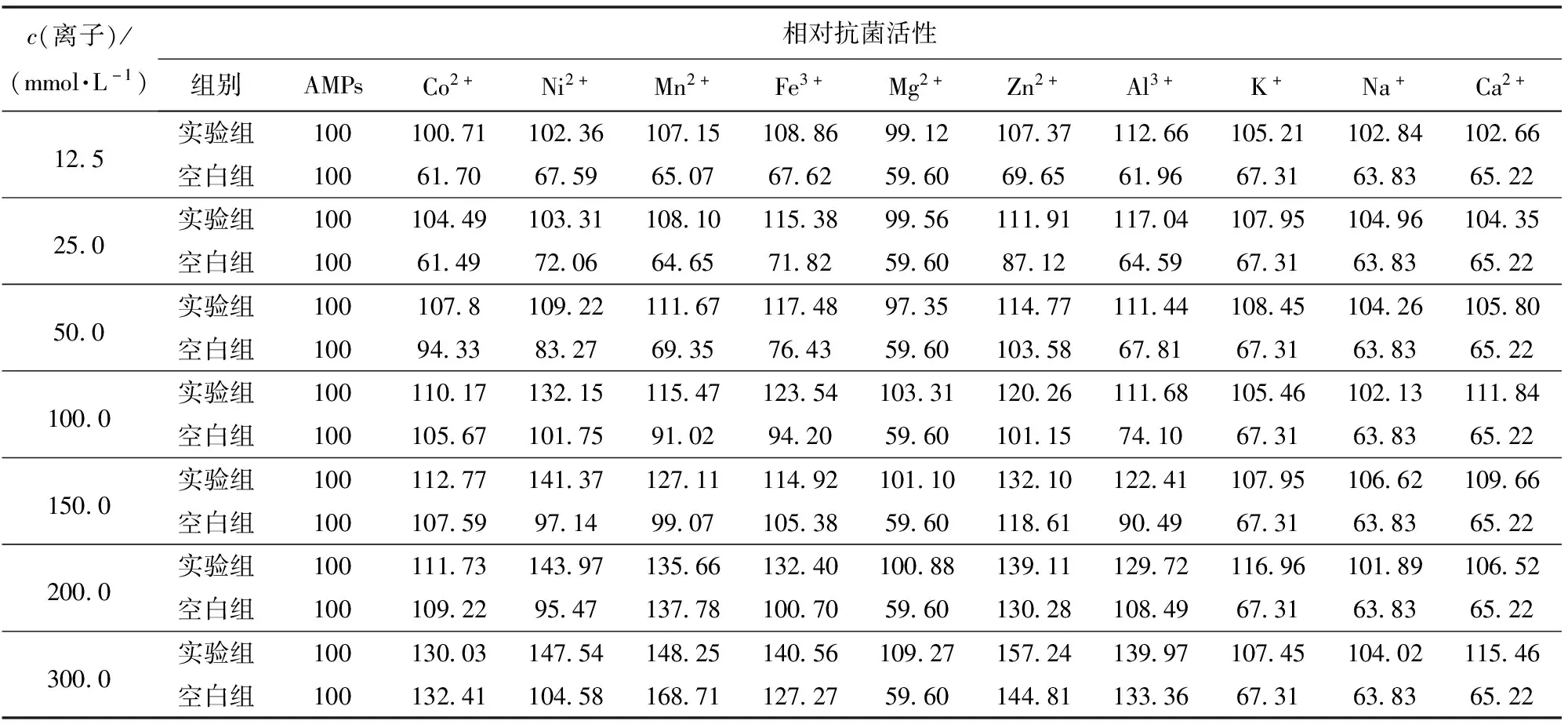
表1 金屬離子對Brevilaterin抗菌活性的影響
2.1.1.2 乳化劑的影響
乳化劑是在面包、蛋糕和餅干等食品中起乳化、分散、潤滑和穩定作用的常用添加劑(添加量在0.5%~5.0%)[18-19]。乳化劑在疏水和靜電作用下,很可能會與蛋白或肽類結合,進而影響它們的活性[20],如G?nzle等[7]曾研究發現,1%的卵磷脂會使Nisin完全失去抗菌活性。乳化劑對Brevilaterin抗菌活性研究的實驗結果見圖1。由圖1可見,Brevilaterin的抗菌活性除了受到大豆磷脂一定程度的抑制以外,均不受其他乳化劑的影響。
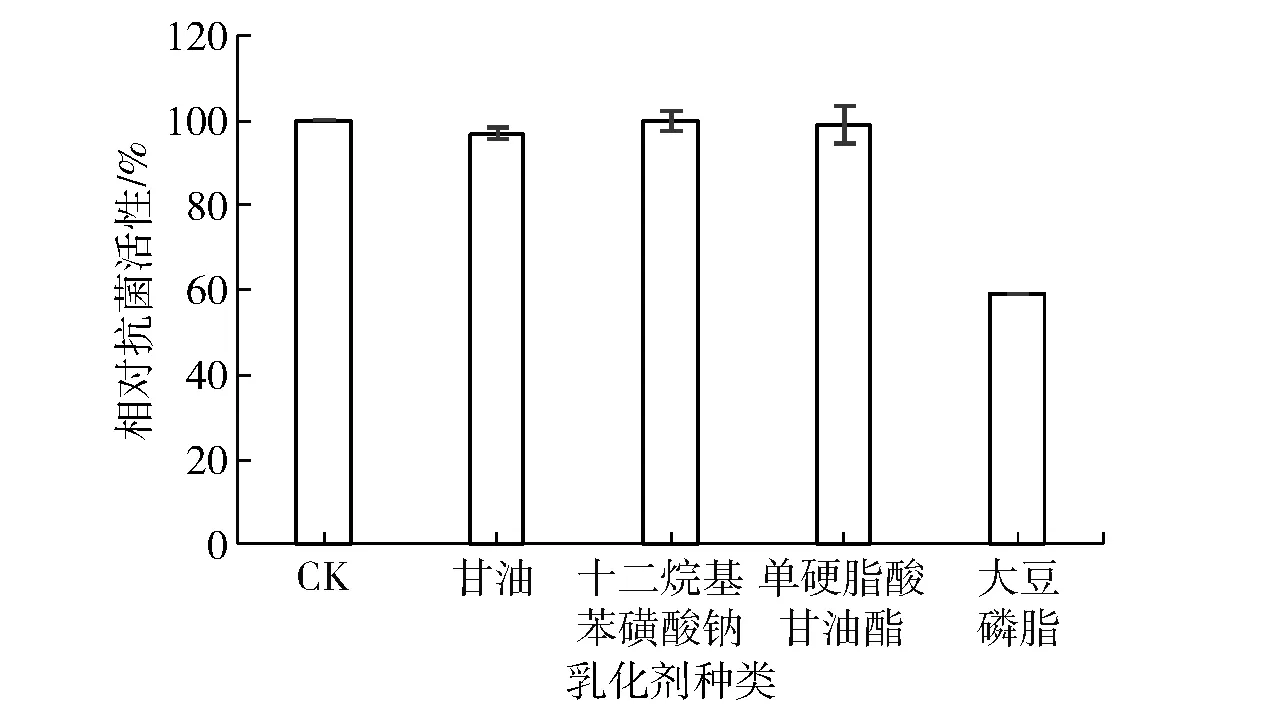
圖1 乳化劑對Brevilaterin抗菌活性的影響
2.1.1.3 增稠劑的影響
增稠劑主要用于改善和增加食品的黏稠度,保持流態食品、膠凍食品的色、香、味和穩定性,改善食品物理性狀,并能使食品有潤滑適口的感覺。增稠劑常添加在果凍、調味糖漿、軟糖等食品中[21],常用添加量為0.5%~5.0%[22]。如李鵬等[23]發現,用質量分數為1.05%卡拉膠和0.45%海藻酸鈉組成的復合膠制備南瓜果凍,能更好地保持果凍的柔軟性和彈性。增稠劑對Brevilaterin抗菌活性研究的實驗結果見圖2。由圖2可見,Brevilaterin的抗菌活性基本不受增稠劑的影響,因此,Brevilaterin適合果凍、軟糖等食品的防腐應用。

圖2 增稠劑對Brevilaterin抗菌活性的影響
2.1.1.4 酶制劑的影響
焙烤食品、釀造食品、肉類食品等,在加工過程中通常會添加某種酶制劑來有效提高其加工程度、改進加工工藝以及改善其風味和品質[24-25];然而,酶的添加(質量濃度一般達10 mg/mL以上時),尤其是蛋白酶,通常會對蛋白或肽類物質的結構造成破壞,致使其生物活性喪失[12,23]。研究發現,不同來源的AMPs對蛋白酶具有不同的敏感性,如蘿卜籽蛋白提取物基本不受9 mg/mL蛋白酶影響[16],而格氏乳球菌分泌的garviecin LG 34極易被蛋白酶(木瓜蛋白酶、中性蛋白酶、胃蛋白酶和胰蛋白酶,5 mg/mL)水解[26]。基于抗菌肽Brevilaterin是由13個氨基酸、N末端有一個C6脂肪酸組合而成的結構特點,本研究重點考察蛋白酶與脂肪酶對其抗菌活性的影響,實驗結果見圖3。圖3表明,Brevilaterin對多種蛋白酶均表現出很強的抗性;此外,Brevilaterin的抗菌活性同樣不受脂肪酶影響。
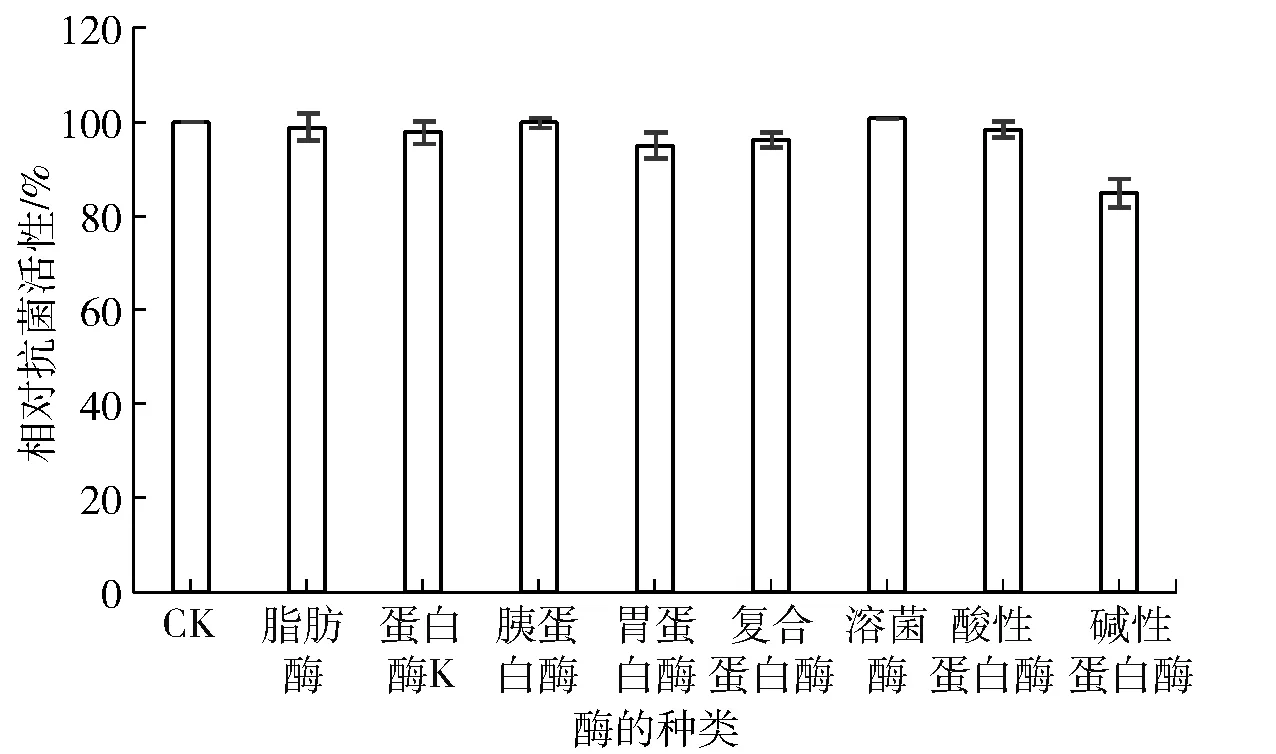
圖3 蛋白酶對Brevilaterin抗菌活性的影響
2.1.1.5 高濃度蔗糖的影響
果脯、蜜餞以及罐頭類食品通常采用過量加糖的方式來擬造高滲的保藏環境,以此來防止微生物的侵染,如此極端的食品屬性就對生物防腐劑提出更高的要求。本研究發現(實驗結果見圖4),Brevilaterin在含有50%蔗糖的環境中依然能夠保持原有的抗菌活力,這表明其非常適用于高滲的保藏環境下的防腐保鮮。
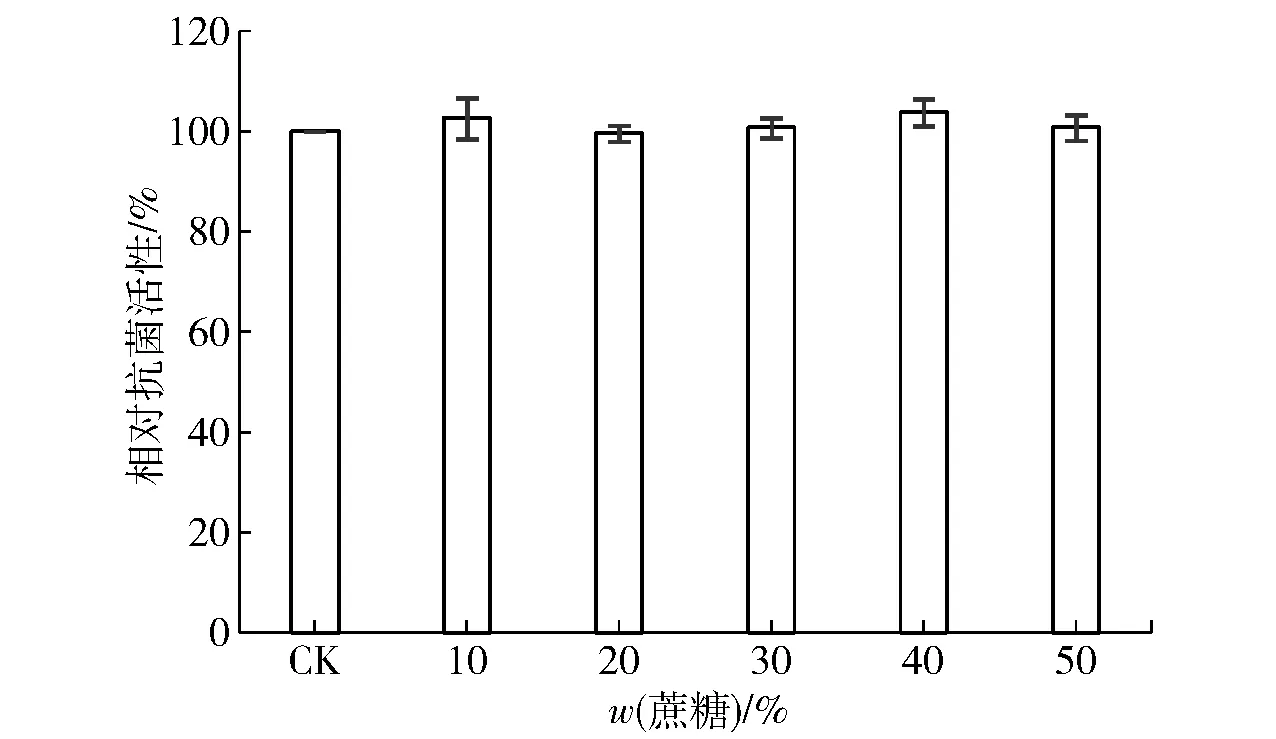
圖4 蔗糖對Brevilaterin抗菌活性的影響
2.1.2反復凍融處理對Brevilaterin抗菌活性的影響
本研究還考察了反復凍融處理對Brevilaterin抗菌活性的影響,實驗結果見圖5。圖5表明,Brevilaterin能夠完全耐受反復凍融處理對其抗菌活性的影響,表現出優異的穩定性。
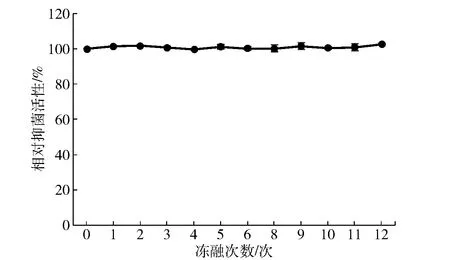
圖5 凍融次數對Brevilaterin抗菌活性的影響
低溫及冷凍環境可以保持水果、蔬菜、奶制品、鮮肉等食品品質(嫩度、持水性、脂肪氧化)和商品品質(色澤、質構),但反復解凍過程又很容易引入食品腐敗菌的侵染;同時,蛋白和肽類物質的生物活性很可能會受凍融處理的影響而改變,給其應用帶來一定的隱患[27]。圖5顯示,Brevilaterin在經過12次反復凍融之后依然能夠保持原有的抗菌活力,表明其能夠適應于較大的溫度反復變化。其他來源的AMPs如BSN-37、F1的粗提物,經10~12次、5次反復凍融后[28],抗菌活性也能保持不變。
2.2 食品內部環境變化對Brevilaterin抗菌作用的影響
食品的屬性及其加工工藝不盡相同,為了實現有效抑制食品腐敗菌的生長、延長貨架期,存儲時通常都會選用差異性很大的食品保藏環境。降低保藏溫度、調整酸堿度、創造高滲環境和應用真空包裝或脫氧劑是最常用于控制食品腐敗菌生長的重要方式,但為了延長食品保存的貨架期,目前大部分食品保藏時除了選用上述特殊食品保藏環境外,還需加入一定量的防腐劑來強化抑制食品腐敗菌的生長。
隨著食品貯藏時內部環境的變化,食品腐敗菌的生長狀態和生長量可能會受到不同程度的影響。特別是當生長條件偏離菌株最適生長條件時,菌株的生長會出現緩慢,甚至休眠的狀態,菌體生長量也會隨之出現一定程度的下降[29]。為此,本研究以OD600為指標,考察了受試菌在生長環境發生變化時的生長量變化。結果表明,受試菌在最適生長環境下,菌體生長良好,但逐漸偏離受試菌最適生長條件時,菌體生長緩慢,菌體量出現一定程度的減少(具體結果未列出)。
此外,雖然前期研究已證實Brevilaterin在不同因素的影響以及不同條件的處理下均能保持良好的抗菌效果[12-13],但是,Brevilaterin在食品內部環境發生變化時是否會對食品腐敗菌表現出不同的抗菌作用規律呢?本研究通過模擬不同食品內部環境的變化(溫度、pH值、滲透壓以及氧含量),考察抗菌肽Brevilaterin對食品腐敗菌的抗菌作用規律,旨在明確Brevilaterin能夠抑制或者殺死受試菌所需要添加的參考量。
2.2.1溫度變化對Brevilaterin抗菌作用的影響
選取S.aureus、L.monocytogenes、M.luteus和P.aeruginosa為指示菌,分別測定4 ℃(冷藏)、10 ℃(低溫)、25 ℃(室溫)和37 ℃(菌體最適生長溫度)下Brevilaterin對各指示菌的MIC和MBC,實驗結果見表2。表2表明,Brevilaterin對所有測試溫度下的菌株均有很好的抑制效果,但隨著溫度的改變,Brevilaterin對各指示菌的MIC和MBC也產生了不同程度的變化。在4 ℃下,Brevilaterin對M.luteus的MIC和MBC均為37 ℃下的1/8;在10 ℃下,Brevilaterin對P.aeruginosa的MIC和MBC均為37 ℃下的1/4,對L.monocytogenes的MIC和MBC分別為37 ℃下的1/4和1/2,對S.aureus的MIC和MBC均為37 ℃下的1/2。
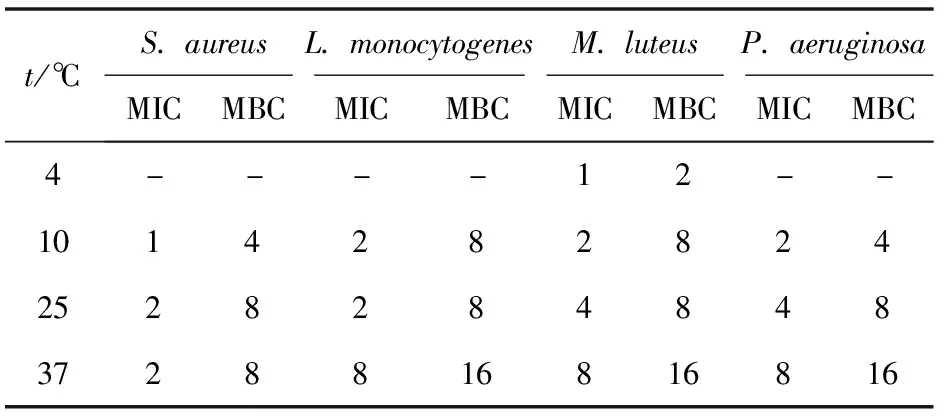
表2 溫度變化對Brevilaterin抗菌作用的影響
溫度是調控微生物生長性狀的主要因素之一,而降低保藏溫度也是食品防腐的重要措施[30]。所以,闡明溫度變化時Brevilaterin對食源性致病菌的抗菌作用規律對于科學地指導AMPs的防腐應用有著重要的意義。本研究發現,隨著溫度的降低,Brevilaterin對各指示菌的MIC和MBC逐漸減小,這也反映出Brevilaterin對食源性致病菌的抗菌作用逐漸增強。可能原因是,低溫條件下微生物逐步進入休眠狀態,生長繁殖趨緩,代謝和免疫等功能逐漸弱化,在此環境下Brevilaterin便能發揮出更好的抗菌效果[31];同時,這也表明了在達到相同的抗菌效果時,Brevilaterin在較低溫度下所需添加的量便能大大減少,降低了其應用成本[32]。
2.2.2pH值變化對Brevilaterin抗菌作用的影響
以L.lactis、S.aureus、H.alkalicola為指示菌,分別測定pH值變化時Brevilaterin對各指示菌的MIC和MBC,實驗結果見表3。表3表明,Brevilaterin對所有測試pH值下的菌株均有很好的抑制效果,但隨著pH值的改變,Brevilaterin對各指示菌的MIC和MBC也產生了不同程度的變化。pH值為2時,Brevilaterin對L.lactis的MIC和MBC降低為pH為6值時的1/2;pH值為8~9時,Brevilaterin對S.aureus的MIC和MBC減小到pH值為7時的1/2;而Brevilaterin在pH值為7.0值時會對H.alka-licola表現出最強的抗菌活性。
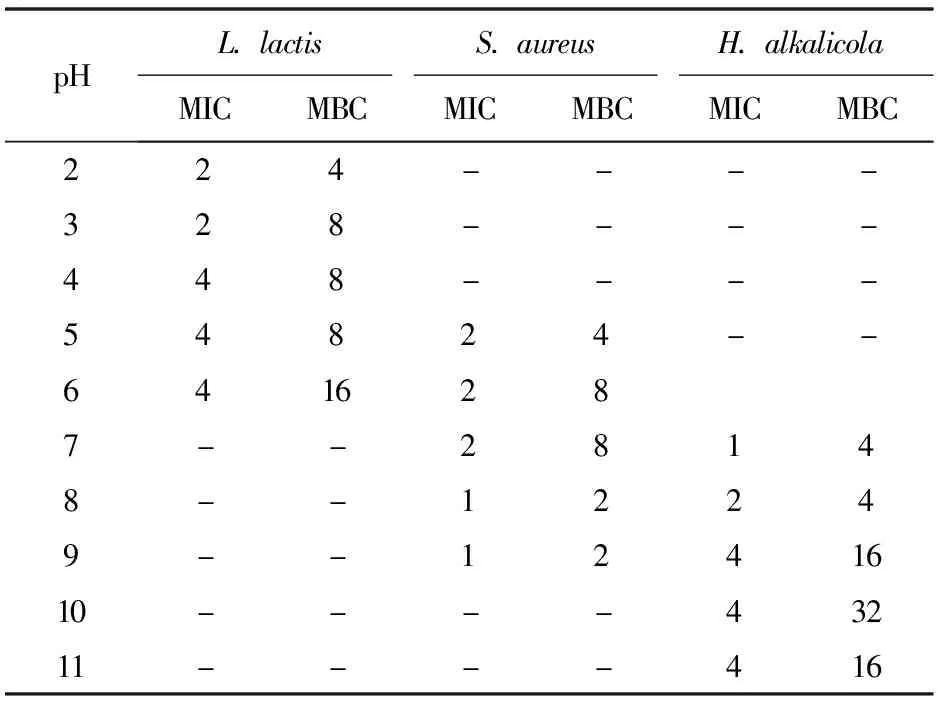
表3 pH值變化對Brevilaterin的抗菌作用的影響
酸堿環境是影響微生物生長的重要因素之一,而調整食品保存的pH值也是有效防止食品腐敗變質的重要方法。本課題組前期研究結果證實,pH值的變化對Brevilaterin抗菌活性并不產生影響[14]。本研究發現,在偏離菌株最適的生長pH值時,Brevilaterin對指示菌的MIC和MBC顯著減小,說明其對食源性致病菌的抗菌作用增強。產生該結果的原因:一方面是因為菌株生長趨緩;另一方面可能是,pH值改變會引起膜電勢的改變[1],從而導致指示菌對Brevilaterin敏感性改變,進而使Brevilaterin的添加量減少,同樣達到防止腐敗的效果。
2.2.3滲透壓變化對Brevilaterin抗菌作用的影響
以S.aureus為指示菌,分別測定不同濃度的蔗糖和NaCl存在環境下,Brevilaterin對S.aureus的MIC和MBC,實驗結果見表4。表4結果表明,隨著蔗糖或者NaCl濃度的改變,Brevilaterin對S.aureus的MIC和MBC也發生了變化。在蔗糖質量分數達12.5%及以上時,Brevilaterin對S.aureus的MBC為初始條件下的1/2;類似的,在8%NaCl存在時,Brevilaterin對S.aureus的MBC為對照的1/2。
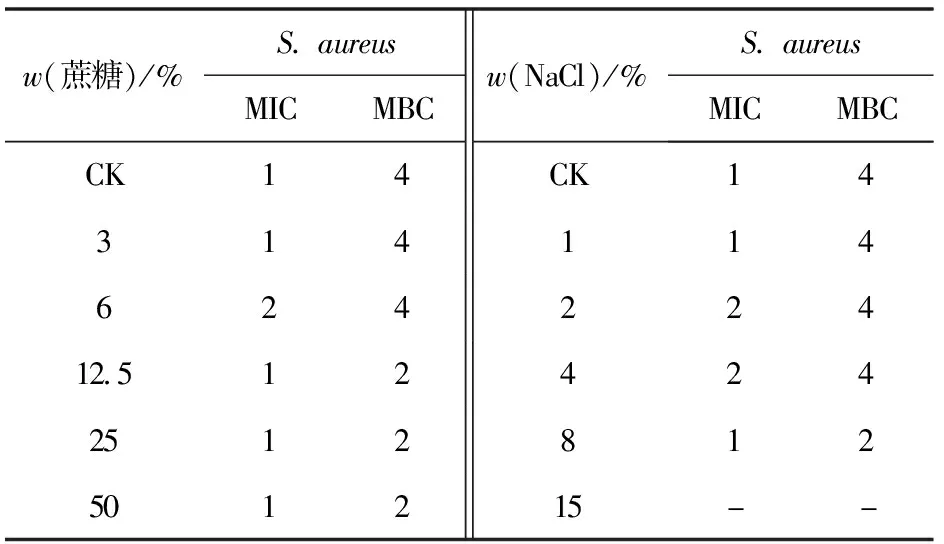
表4 滲透壓變化對Brevilaterin抗菌作用的影響
擬造高滲環境是最常用于控制微生物生長的食品存儲方法之一[33]。本研究發現,當食品的滲透壓較高時,Brevilaterin對S.aureus的MBC會顯著減小,可見其對食源性致病菌的抗菌作用增強,Brevilaterin與滲透壓的協同能對食源性致病菌產生更強的抗菌作用。產生該結果的可能原因是,微生物離開其生長最適的水分活度后[34],菌株逐漸出現緩慢、休眠等狀態,菌體量也隨之降低,進而使得Brevilaterin抑制指示菌所需用量減少。
2.2.4氧含量對Brevilaterin抗菌作用的影響
以B.fusiformis和S.aureus為指示菌,分別測定氧氣體積分數為5%、10%、15%、21%時,Brevilaterin對各指示菌的MIC和MBC,實驗結果見表5。表5表明,氧含量的變化僅能一定程度上調控菌株的生長狀態,但并不影響Brevilaterin對指示菌的抗菌效果。
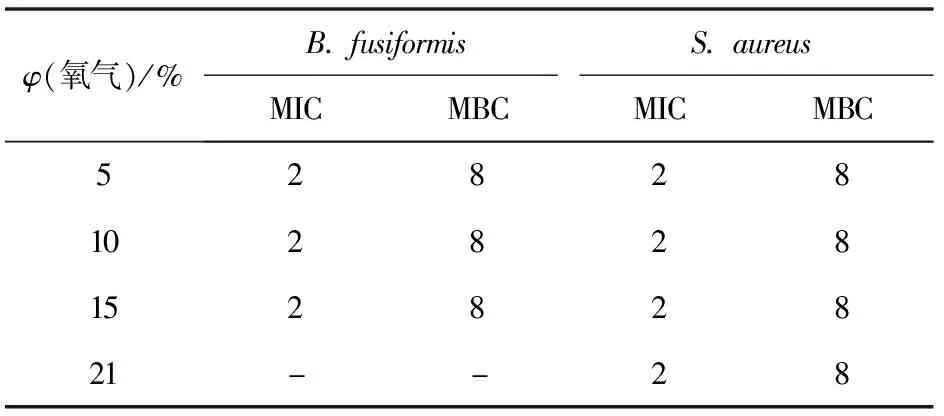
表5 氧含量變化對Brevilaterin抗菌作用的影響
氧氣是影響微生物生長不可缺少的因素之一,使用真空包裝或應用脫氧劑都是食品防腐的重要手段[35],但常見的食源性致病菌多屬于兼性厭氧型菌株,它們在有氧和無氧環境中均能生長,如大腸桿菌、沙門氏菌。本研究發現,氧含量的變化并不影響Brevilaterin對厭氧菌B.fusiformis和對照菌S.aureus的抗菌作用,因此Brevilaterin在厭氧環境也能達到防腐抗菌的效果。
3 結 論
本研究考察了不同外界因素對Brevilaterin抗菌活性的影響,結果表明,Brevilaterin能抵抗除大豆磷脂以外的所有因素以及反復凍融處理對其抗菌活性的影響。本研究還深入探討了食品內部環境變化對Brevilaterin抗菌作用規律的影響,結果表明,隨著溫度、pH值和滲透壓的改變,Brevilaterin的抗菌作用規律均會發生顯著變化。隨著溫度的降低,Brevilaterin對S.aureus、L.monocytogenes、M.luteus和P.aeruginosa的抗菌作用逐漸增強,MIC和MBC減少至菌株生長最適溫度下的1/8~1/2;當pH值偏離L.lactis、S.aureus和H.alkalicola生長最適pH值時,Brevilaterin對這些菌株的MIC和MBC均減小為初始的1/4~1/2;滲透壓升高后,Brevilaterin對S.aureus的MBC也降低為常壓下的1/2。總體上,當食源性致病菌的生長條件偏離其最適條件時,Brevilaterin能表現出更強的抗菌作用,而此時的MIC和MBC均降低至最適條件下的1/8~1/2。Brevilaterin良好的穩定性,以及能隨著食品內部環境變化而表現出不同的抗菌作用的特點,可為Brevilaterin在食品防腐中的科學應用提供理論依據。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
