徐淮地區7個甘薯莖線蟲群體的形態學及分子生物學鑒定
2021-01-29 10:44:14陳晶偉,馬居奎,張成玲,楊冬靜,唐偉,謝逸萍,孫厚俊
江蘇農業學報 2021年6期
陳晶偉,馬居奎,張成玲,楊冬靜,唐偉,謝逸萍,孫厚俊
摘要:從江蘇省徐淮地區7個采集地的甘薯塊根中分離到7個甘薯莖線蟲群體,通過形態學鑒定,其特征與已報道的腐爛莖線蟲(Ditylenchus destructor)基本一致。利用rDNA-內轉錄間隔區(ITS)通用引物、腐爛莖線蟲特異性引物檢測,發現群體JSXZ為B(L)型,其余6個分離群體均為A(S)型腐爛莖線蟲。結合ITS-5.8S-ITS2區序列分析并構建系統發育樹,結果進一步表明,所獲序列與腐爛莖線蟲相似性最高,且各分離群體與徐淮地區已報道的相同基因型的腐爛莖線蟲群體序列相似性均在99%以上。群體JSXZ沒有與腐爛莖線蟲A型、B型群體聚在一起,而是與C型、D型群體聚為1個分支,且和C型親緣關系更近,所以群體JSXZ被鑒定為C型腐爛莖線蟲;群體JSWJ、JSYJ、JSGS、JSHW、JSLQ和JSNG與A型腐爛莖線蟲群體聚為1個分支,均為A型腐爛莖線蟲,且與其他基因型區分明顯。危害徐淮地區甘薯的腐爛莖線蟲親緣關系近,基因型種類多,存在A型、B型和C型,其中A型腐爛莖線蟲為優勢群體。
關鍵詞:甘薯;腐爛莖線蟲;鑒定;系統發育樹
中圖分類號:S432.4+5文獻標識碼:A文章編號:1000-4440(2021)06-1409-08
Morphological and molecular biology identification of seven stem nematode populations of sweet potato in Xuhuai area
CHEN Jing-wei,MA Ju-kui,ZHANG Cheng-ling,YANG Dong-jing,TANG Wei,XIE Yi-ping,SUN Hou-jun
(Xuzhou Institute of Agricultural Sciences in Xuhuai Area of Jiangsu/Key Laboratory of Biology and Genetic Improvement of Sweet Potato of Ministry of Agriculture and Rural Affairs, Xuzhou 221131, China)
Abstract:Seven populations of sweet potato stem nematodes were isolated from storage roots of sweet potatoes from seven regions in Xuhuai area of Jiangsu province. Characteristics of the stem nematodes were basically identical to Ditylenchus destructor reported previously through morphological identification. By detections using universal primers of rDNA- internal transcribed spacer (ITS) region and specific primers of D. destructor, it was found that population JSXZ belonged to B(L) type of D. destructor, while the other six isolated populations all belonged to A(S) type of D. destructor. The results further suggested that, the sequences of the stem nematodes shared the highest similarity with D. destructor though sequences analysis of ITS-5.8S-ITS2 region and phylogenetic tree construction, and sequence similarity of the separated populations compared with the D. destructor populations carrying the same genotype reported in Xuhuai area were all above 99%. Results of phylogenetic tree analysis showed that population JSXZ did not cluster into one branch with type A and type B populations of D. destructor, but clustered into one branch with type C and type D populations of D. destructor, and had closer affinity with type C. Therefore, JSXZ population from the sweet potato was confirmed as type C of D. destructor. Populations of JSWJ, JSYJ, JSGS, JSHW, JSLQ and JSNG were clustered into one branch with population of type A of D. destructor, which were all confirmed as type A of D. destructor and were clearly distinguished from other genotypes. It was indicated that populations of D. destructor which harmed sweet potatoes in Xuhuai area were closely related and had various genotypes, such as type A, type B and type C, among which type A was the dominant population.
Key words:sweet potato;Ditylenchus destructor;identification;phylogenetic tree
莖線蟲病害嚴重威脅中國甘薯產業的發展,其病原物為腐爛莖線蟲(Ditylenchus destructor),最早在馬鈴薯(Solanum tuberosum)上被發現,故也稱Potato nematode[1]。國內腐爛莖線蟲首先發現于甘薯(Ipomoea batatas)中,主要侵染貯藏根并引起腐爛,造成糠心或糠皮等癥狀,危害嚴重時可導致減產80%以上,乃至絕收[2-3]。隨后,又在馬鈴薯及一些中藥材上發現其危害[4-5]。現已被亞太植物保護組織及許多國家和地區列為植物檢疫性有害生物[6]。
線蟲形態特征復雜且有相似的特點,僅依靠形態學鑒定,難以在種水平及基因型上進行區分,因此分子生物學檢測已成為研究植物寄生線蟲的常用手段[7]。據相關文獻報道,腐爛莖線蟲存在種內分化的現象,如王金成等[8]、黃健等[9]、章淑玲等[10]分析了國內不同腐爛莖線蟲群體的rDNA-內轉錄間隔區(ITS)序列后發現,中國的腐爛莖線蟲明顯存在2種基因型,即A(S)型和B(L)型。宛菲等[11]根據腐爛莖線蟲rDNA-ITS區分別設計了針對這2種基因型的特異性引物,并準確檢測出國內6個省(市、區)的21個甘薯腐爛莖線蟲群體的基因型。國外學者也深入研究了腐爛莖線蟲群體分化的現象,如Subbotin等[12]對世界范圍內不同寄主植物的腐爛莖線蟲群體的rDNA-ITS區序列進行分析比對后發現,不同線蟲群體之間ITS1-5.8S-ITS2序列長度的變異源自ITS1區存在的重復序列,且該重復序列能在RNA二級結構中形成穩定的莖環,在保留A(S)型的基礎上,將B(L)型進一步細分為B~G 6種不同的基因型。Jeszke等[13]結合NCBI已發表的腐爛莖線蟲rDNA-ITS區序列構建系統發育樹,鑒定出波蘭3個不同地區腐爛莖線蟲分離種群均為C基因型。現已發現危害甘薯的腐爛莖線蟲基因型有A型、B型、C型、E型和F型[10,12,14]。
甘薯對于帶動區域農業農村經濟發展有著舉足輕重的作用,且已成為國內許多省份及地區調整農業產業結構、促進農業農村發展的重要農作物之一[15-16]。目前,甘薯莖線蟲病害在中國北方薯區春作地發生嚴重[2]。江蘇省甘薯產業總體發展態勢良好,但甘薯莖線蟲病害嚴重制約甘薯單位面積產量的提升,阻礙甘薯產業發展。本研究針對江蘇省徐淮地區不同甘薯莖線蟲分離群體進行形態學和分子生物學鑒定,并基于rDNA-ITS區構建系統發育樹,以明確線蟲危害種類及系統發育地位,為甘薯莖線蟲病害的快速診斷和防控提供理論依據。
1材料與方法
1.1試驗材料
2019-2020年,對江蘇省徐淮地區7個線蟲危害嚴重的薯田進行樣品采集,并拍攝記錄田間甘薯受害癥狀。采用隨機采樣法,將受害薯塊裝入聚乙烯薄膜自封袋內,并標記采集時間、地點、前茬作物及土壤類型等信息,帶回實驗室進一步分離與鑒定。DdSH、DdTS、DdXY、DEsg為江蘇徐淮地區已報道的甘薯上的腐爛莖線蟲群體[8,11](表1)。
1.2線蟲的分離與純化
將病薯切成碎屑,利用淺盤法分離線蟲[1]。獲得線蟲后,用0.1%青霉素+0.1%鏈霉素浸泡,4 ℃下放置12 h消毒,用無菌水清洗3遍后保存備用。在體式顯微鏡下挑取已消毒的雌蟲和雄蟲各50頭放入茄腐鐮刀菌(Fusarium solani)培養皿中進行純化培養,保存備用。
1.3線蟲的形態觀察和測量
線蟲的殺死、TAF固定、臨時玻片制作均參照文獻[17]。通過配備Leica相機(DFC550)的顯微鏡(Leica DM4000B)觀察制作的線蟲臨時玻片。使用相機配套軟件進行拍攝并記錄各蟲態形態特征圖片;形態測量值利用配套軟件,按照De Man公式[17]進行測量計算,即a=體長/最大體寬;b=體長/食道長度(從頭到食道與腸結合處);c=體長/尾長;V=頭至陰門距離/體長×100。每個特征值,觀察20個標本。
1.4線蟲分子生物學鑒定
1.4.1DNA提取方法參考單條花生莖線蟲(D.arachis) DNA 提取方法[18],用ddH2O把挑到的線蟲清洗干凈后,轉移至裝有8 μl ddH2O、1 μl 10×PCR buffter的PCR管內,將PCR管在掌上離心機中快速離心后,放至液氮中浸泡約30 s,然后加1 μl蛋白酶K(1.2 mg/ml)到管內。最后,將PCR管放置于PCR儀中于65 ℃溫育1.5 h,再于95 ℃放置10 min,取出放至室溫即可用于PCR擴增或置于-20 ℃備用。
1.4.2PCR引物擴增試驗所用引物均由生工生物工程(上海)股份有限公司合成。反應體系為25.0 μl,其中模板DNA 2.0 μl,上游引物(10 μmol/L)1.0 μl,下游引物(10 μmol/L)1.0 μl,PCR Mix(Premix TaqTM,TaKaRa)12.5 μl,ddH2O 8.5 μl。
(1)rDNA-ITS序列擴增。采用引物ITSP1/ITSP2(5′-CGTAACAAGGTAGCTGTAG-3′,5′-TTTCACTCGCCGTTACTAAGG-3′)擴增[8]。反應程序:94 ℃,4 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 2 min,35個循環;72 ℃ 10 min,4 ℃保存。
(2)特異性引物擴增。A(S)型特異性引物為DdS1/DdS2(5′-TCGTAGATCGATGAAGAACGC-3′,5′-ATTATCTCGAGTGGGAGCGC-3′);B(L)型特異性引物為DdL1/DdL2(5′-TTGTGTTTGCTGGTGCGCTTGT-3′,5′-GAGTGAGAGCGATGTCAACATTG-3′)[11]。反應程序:94 ℃ 5 min;94 ℃ 30 s,50 ℃ 30 s,72 ℃ 1 min,35個循環;72 ℃ 10 min;4 ℃保存。
用1%的瓊脂糖凝膠對PCR產物進行電泳確定目的條帶,擴增產物寄送至生工生物工程(上海)股份有限公司進行雙向測序。
1.4.3序列提交、分析及系統發育樹構建依據序列提交步驟將本試驗所得序列上傳至GenBank,獲得序列號(表1)。采用MEGA 6.0和DNAMAN軟件進行多序列比對分析。采用jModelTest v.2.1.2AIC標準進行最佳匹配替代模型估算[19],采用DAMBE v.7.0.28軟件[20]進行基因替代飽和度檢測。在CIPRES Science Gateway(www.phylo.org)[21]平臺上進行在線序列分析。分別使用MrBayes 3.2.6 on XSEDE[22]和RAxML-HPC2 on XSEDE[23-24]進行貝葉斯法與最大似然法分析。貝葉斯系統發育樹分析采用GTR+I+G模式,運算5 000 000代,每運算100代對樹形進行Markov chains采樣,剔除初始25%采樣;最大似然法分析采用GTRCAT模型,完成1 000次自展重復。采用Figtree v1.4.3和Adobe Illustrator CC在貝葉斯50%多數原則一致樹上標注自展值和后驗概率。
2結果與分析
2.1甘薯莖線蟲病主要癥狀特點
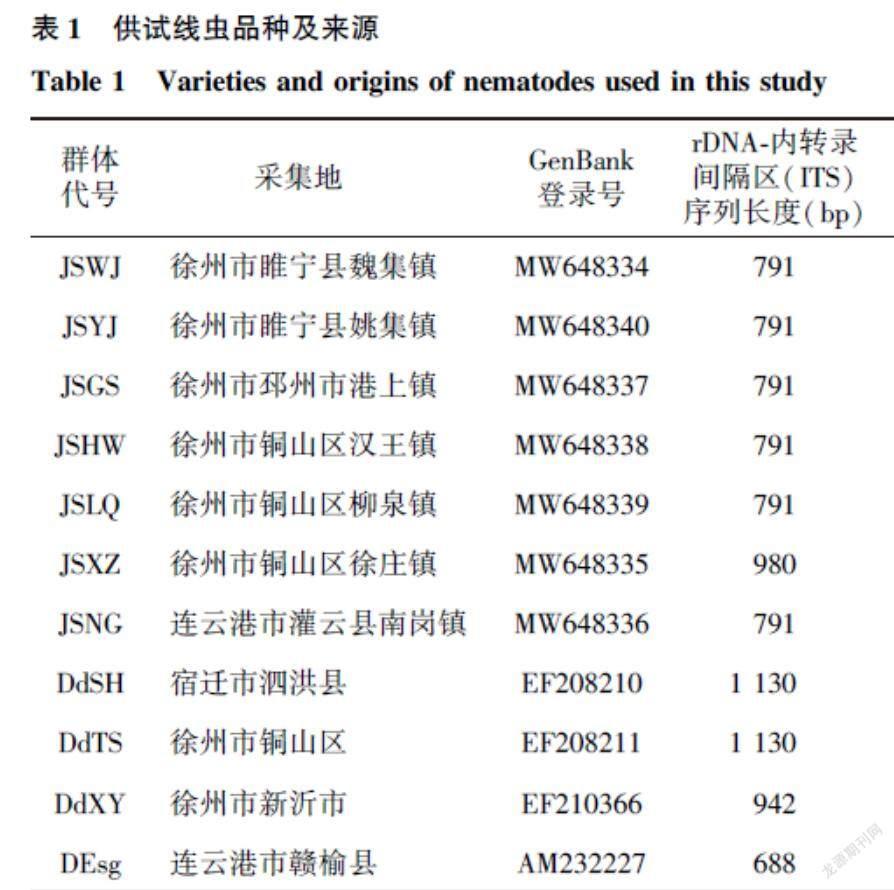
受病原線蟲危害的大田中,甘薯地上部分在生長前期癥狀不明顯,癥狀在中后期顯現,薯蔓近地面的主蔓基部縱切后可見髓部呈黑褐色干腐樣,呈糠心狀,而發病嚴重的植株糠心可達薯蔓頂部,葉片逐步發黃,直至主蔓枯死(圖1A、圖1B、圖1C)。薯塊受害后大多表現為糠皮或糠心,喪失經濟價值。糠皮型薯塊表皮褪色泛青,有的還會出現凹陷或裂口,導致薯皮龜裂。縱切后可見皮下組織干腐變褐,呈圓環狀(圖1D、圖1E)。糠心型薯塊的皮層與健康薯塊無差異,但是薯塊質量顯著下降,切開后可見薯塊內呈褐白相間的干腐癥狀(圖1F)。甘薯受害后期,發病嚴重時,糠皮和糠心會混合發生,并常伴隨真菌、細菌和螨類等二次侵染(圖1G)。
2.2甘薯莖線蟲形態學鑒定
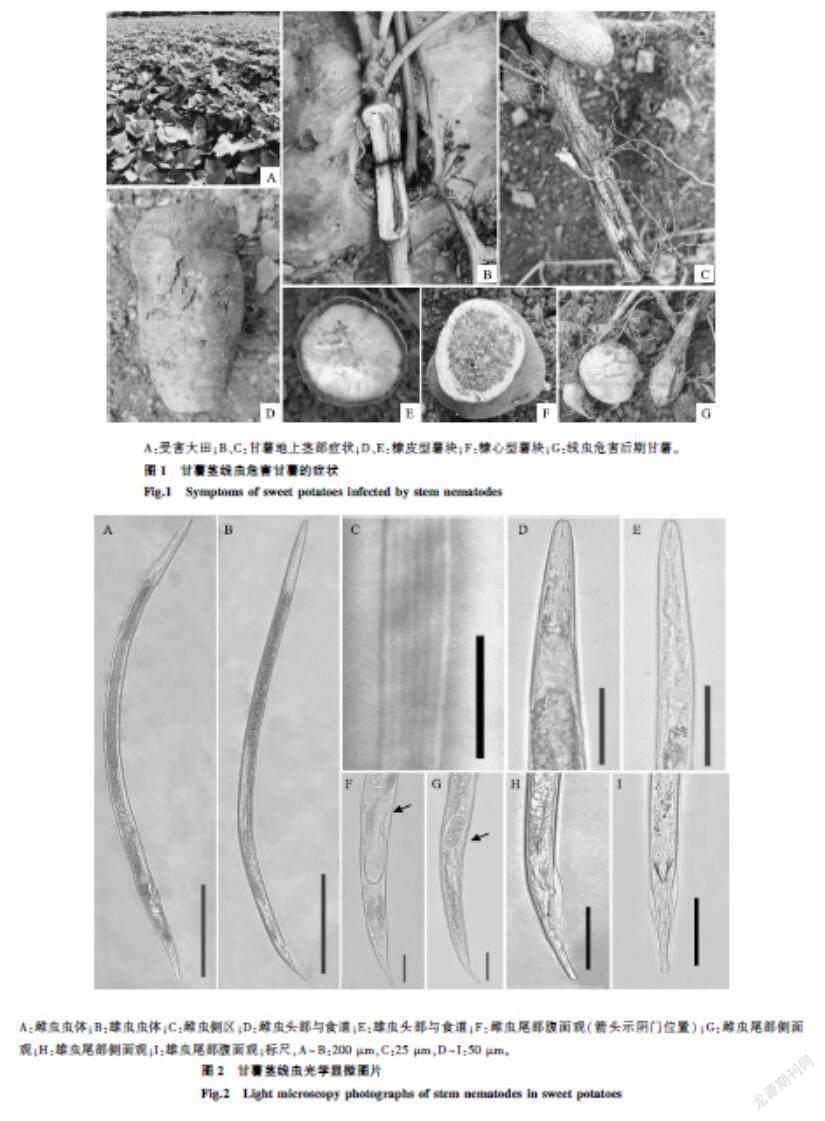
雌蟲溫熱殺死后可見整個蟲體略向腹面彎曲(圖2A)。蟲體側區可見6條清晰側線(圖2C)。唇區平滑,頭區稍有縊縮,口針細直,口針基部球明顯,中食道球似紡錘形,瓣膜可見,排泄孔位于食道腸道連接處略靠上方,后食道腺覆蓋于腸道背面(圖2D)。陰門明顯,側面觀可見向外突起,后陰子宮囊明顯,長約占肛陰距的2/3,尾略向腹面彎曲,末端圓滑(圖2F、圖2G)。
雄蟲溫熱殺死后,蟲體前端較為平直,尾端向腹面彎曲度大(圖2B)。交合刺發達,尾部側面可見交合刺向腹面彎曲,交合傘由交合刺前端延伸至尾長約3/4處,尾端窄圓(圖2H、圖2I)。其余特征與雌蟲類似。
A:受害大田;B、C:甘薯地上莖部癥狀;D、E:糠皮型薯塊;F:糠心型薯塊;G:線蟲危害后期甘薯。
A:雌蟲蟲體;B:雄蟲蟲體;C:雌蟲側區;D:雌蟲頭部與食道;E:雄蟲頭部與食道;F:雌蟲尾部腹面觀(箭頭示陰門位置);G:雌蟲尾部側面觀;H:雄蟲尾部側面觀;I:雄蟲尾部腹面觀;標尺,A~B:200 μm,C:25 μm,D~I:50 μm。
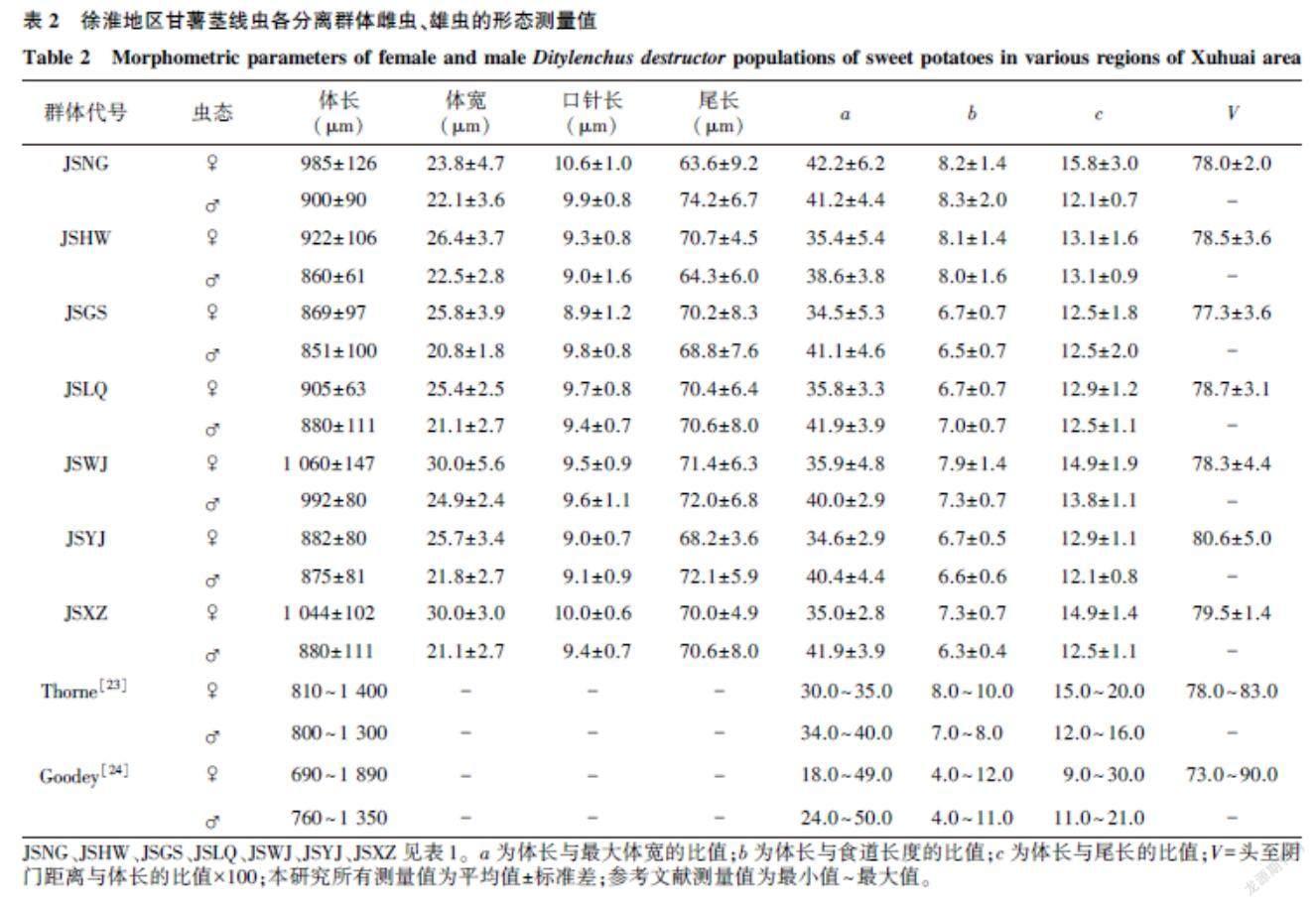
本研究中分離純化的甘薯線蟲群體的形態測量值見表2。由表2可知,部分線蟲分離群體與Thorne[25]記載的腐爛莖線蟲形態測量值存在一定差異,但與Goodey[26]的記載相符。根據主要蟲態的形態學特征和主要形態測量值,初步推斷各甘薯線蟲群體均為腐爛莖線蟲。
2.3分子生物學鑒定
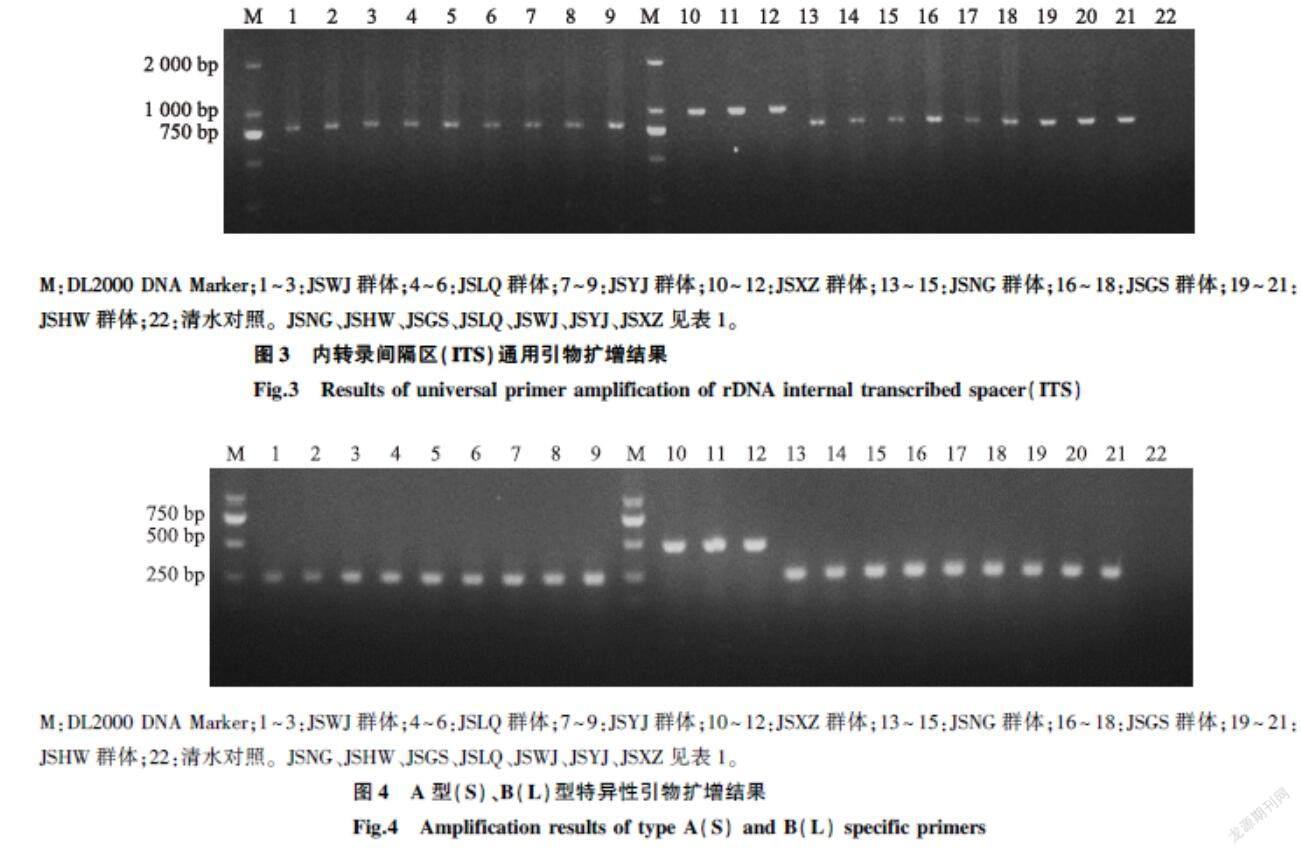
2.3.1甘薯莖線蟲不同分離群體rDNA-ITS通用引物檢測結果分別以江蘇徐淮地區7個不同的甘薯莖線蟲總DNA為模板,采用通用引物ITSP1/ITSP2擴增rDNA-ITS區序列,電泳結果表明,7個分離群體樣本中,除JSXZ群體擴增出約1 000 bp條帶外,其余皆擴增出約780 bp條帶(圖3),結合形態鑒定結果,推斷JSXZ群體可能為B(L)型腐爛莖線蟲,其余均為A(S)型腐爛莖線蟲。
2.3.2A(S)型和B(L)型特異性引物擴增結果為進一步確認各分離群體基因型,針對江蘇徐淮地區不同甘薯莖線蟲分離群體,分別隨機挑取單頭成蟲提取DNA,使用特異性引物DdS1/DdS2與DdL1/DdL2進行PCR擴增,除JSXZ群體擴增出485 bp條帶外,其余皆為252 bp,對照組沒有條帶(圖4),確定JSXZ群體為B(L)型腐爛莖線蟲,其余群體均為A(S)型腐爛莖線蟲。
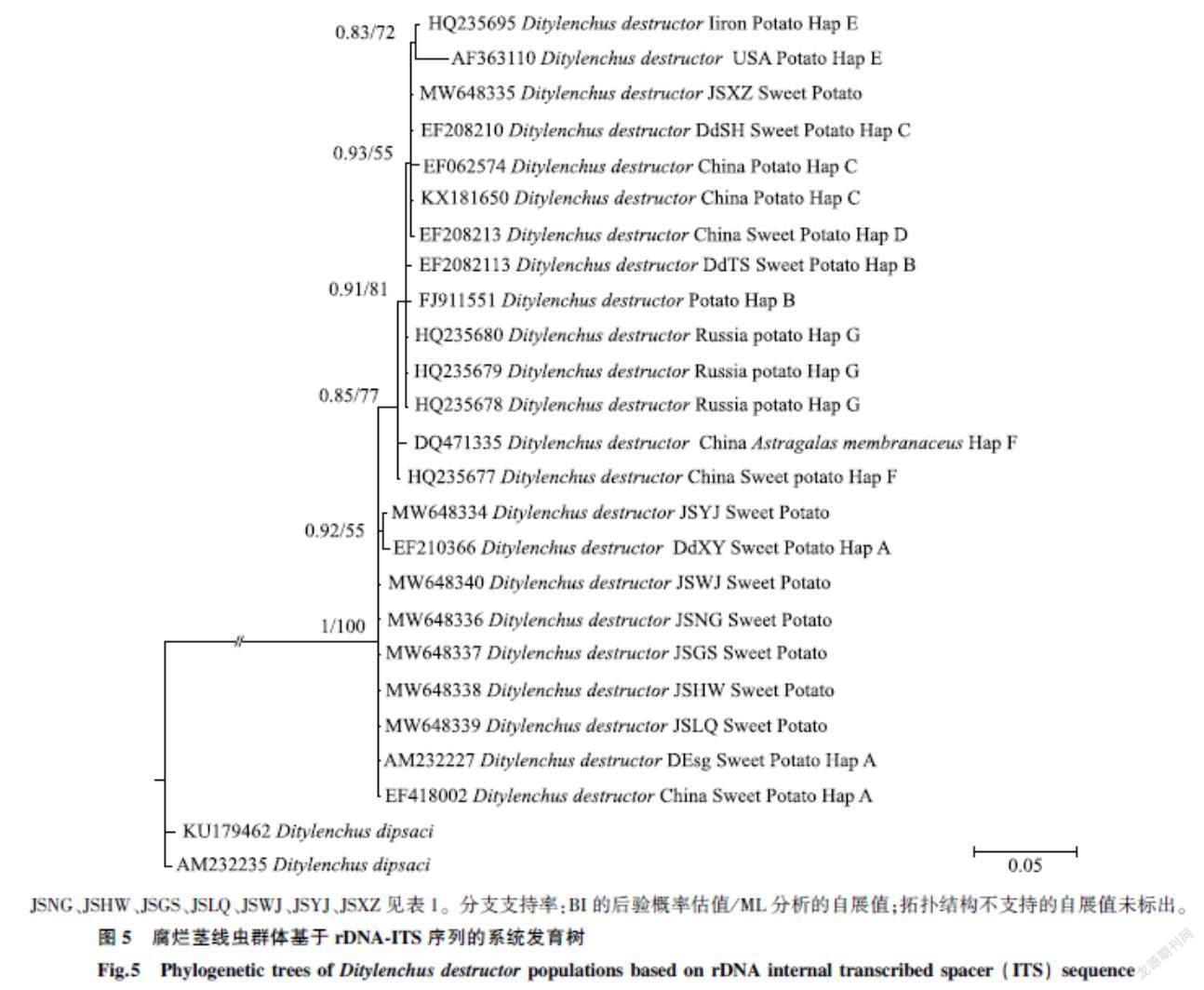
2.3.3基于rDNA-ITS的系統發育樹構建及序列分析對采自江蘇徐淮地區甘薯莖線蟲的不同分離群體進行靶標基因的測定,通過NCBI中BLAST進行比對,發現本研究所有群體與GenBank中江蘇省內腐爛莖線蟲群體序列相似性在99%~100%,獲取序列后上傳至GenBank,獲得序列登錄號,詳細信息見表1。依據比對結果篩選數據庫中17條不同基因型的腐爛莖線蟲序列,2條起絨草莖線蟲(D. dipsaci)序列,加上本試驗獲得的7條序列,共26條序列,基于ITS1-5.8S-ITS2基因,采用貝葉斯和最大似然法構建系統發育樹。結果(圖5)表明所有D. destructor群體聚為一支,與2條D. dipsaci序列區分明顯。JSYJ(MW648334)、JSWJ(MW648340)、JSGS(MW648337)、JSHW(MW648338)、JSLQ(MW648339)和JSNG群體(MW648336)與已知A型腐爛莖線蟲群體聚為一支,表明該6個群體均為A型腐爛莖線蟲,與分子鑒定結果一致。其中,JSYJ群體與DdXY群體聚為一支,表明兩者遺傳距離較近。
JSXZ群體(MW648335)與A型、B型群體未聚在一起,而是與已知的C型群體(EF208210、EF062574、KX181650)和D型群體(EF208213)聚為一支。通過DNAMAN軟件進行序列比對分析,JSXZ群體與已知的C型群體(EF208210、EF062574、KX181650)相似性均達99%以上,其中與宿遷市C型甘薯群體EF208210的序列在ITS1區第193 bp處有1個堿基替換。與C型馬鈴薯群體EF062574在ITS1區41 bp和363 bp處各有1個堿基替換,與C型馬鈴薯群體KX181650在ITS1區96 bp、126 bp、203 bp、215 bp和227 bp處各存在1個堿基替換。而與D型群體(EF208213)同源性僅為96%,在ITS區共存在29個堿基差異,分別在ITS1區存在2個堿基缺失、2個堿基插入以及25個堿基的替換。因此,JSXZ群體與3個C型群體親緣性更近,確定其為C型腐爛莖線蟲。
M:DL2000 DNA Marker;1~3:JSWJ群體;4~6:JSLQ群體;7~9:JSYJ群體;10~12:JSXZ群體;13~15:JSNG群體;16~18:JSGS群體;19~21:JSHW群體;22:清水對照。JSNG、JSHW、JSGS、JSLQ、JSWJ、JSYJ、JSXZ見表1。
M:DL2000 DNA Marker;1~3:JSWJ群體;4~6:JSLQ群體;7~9:JSYJ群體;10~12:JSXZ群體;13~15:JSNG群體;16~18:JSGS群體;19~21:JSHW群體;22:清水對照。JSNG、JSHW、JSGS、JSLQ、JSWJ、JSYJ、JSXZ見表1。
JSNG、JSHW、JSGS、JSLQ、JSWJ、JSYJ、JSXZ見表1。分支支持率:BI的后驗概率估值/ML分析的自展值;拓撲結構不支持的自展值未標出。
3討論
本研究應用形態學結合分子生物學手段對江蘇徐淮地區主要甘薯產區的病原線蟲進行了鑒定,對采集的7個甘薯線蟲群體進行形態學觀測,其形態特征均與張紹升等[27]對腐爛莖線蟲的描述一致。部分群體測量值與Thorne[25]的原定種(正模)的參考值存在一定差異。對此,Goodey[26]、Wu等[28]認為這可能是由于寄主的變化,使得線蟲的各測量值發生一定程度的變化,并定出比原定種(正模)測量數據范圍更廣的變異范圍,本研究所有測量數據均在此變異范圍內。鑒于寄主或不同地理因素差異會導致線蟲形態數據的變異,應注意每個測量值的觀測線蟲數目不應過少。
目前,諸多研究結果已表明腐爛莖線蟲具有明顯的種內分化現象,如Jones等[29]認為腐爛莖線蟲存在寄主專化小種;王金成等[8]對8個腐爛莖線蟲種群ITS區序列進行分析,發現中國腐爛莖線蟲的7個地理種群可明顯分為A、B 2個分支,A分支種群在ITS1區有188 bp片段的缺失,并推測腐爛莖線蟲在中國很可能是1個至少由2種線蟲組成的復合種。章淑玲等[10]設計引物對甘薯腐爛莖線蟲rDNA-ITS區進行分析比對,發現中國的腐爛莖線蟲群體的ITS1區序列分化為短型(S)和長型(L)2種基因型。宛菲等[11]設計并篩選出A(S)型、B(L)型腐爛莖線蟲的2對特異性引物DdS1/DdS2和DdL1/DdL2,對甘薯腐爛莖線蟲rDNA-ITS區進行分析比對,將宿遷市甘薯莖線蟲群體(DdSH)和徐州市銅山區甘薯莖線蟲群體(DdTS)鑒定為B(L)型腐爛莖線蟲。Subbotin等[12]基于腐爛莖線蟲ITS1區中重復序列在RNA二級結構的差異,并結合rDNA-ITS系統發育關系,在前人研究基礎上將腐爛莖線蟲劃分為A~G 7個基因型,并發現腐爛莖線蟲A(S)型特異性引物只能檢測出A基因型,B(L)型特異性引物可檢測出除A基因型之外的所有基因型,并將群體DdSH鑒定為腐爛莖線蟲C型群體,群體DdTS鑒定為腐爛莖線蟲B型群體。在本研究中,利用ITS通用引物和特異性引物對徐淮地區7個不同的分離種群進行分子鑒定,結果表明JSXZ群體為B(L)型腐爛莖線蟲群體,其余6個均為A(S)型腐爛莖線蟲群體,且各分離群體與徐淮地區已報道的相同基因型甘薯腐爛莖線蟲序列相似性均在99%以上。JSXZ群體與C型DdSH群體序列EF208210幾乎一致,僅在ITS1區有1個堿基替換。系統發育樹分析結果進一步表明,6個A型腐爛莖線蟲群體與已知A型群體單獨聚為一支,JSXZ群體與DdSH群體及其他2個C型群體(EF062574、KX181650)聚為一支,最終將群體JSXZ鑒定為C型腐爛莖線蟲。現已發現危害甘薯的腐爛莖線蟲基因型有A型、B型、C型、E型和F型[10,12,14],本研究結果及文獻報道表明危害徐淮地區甘薯的腐爛莖線蟲基因型種類較豐富,存在A~C 3種基因型,A型為優勢種群。這與于海英等[30]調查發現國內危害甘薯的腐爛莖線蟲中A型群體占大多數且分布廣泛的結果一致。而就目前的發現來看,A型腐爛莖線蟲僅在中國出現,且只危害甘薯,這表明中國腐爛莖線蟲可能存在寄主專化性。
腐爛莖線蟲不同地區群體的差異明顯,說明其正處于快速進化過程中,且不同地理來源的線蟲種群在形態特征[9]、致病力[31]、對乙酰膽堿酯酶抑制劑的敏感性[32]和抗藥性[33]等方面存在一定差異。徐振等[34]的研究結果表明甘薯腐爛莖線蟲在南方薯區存在較大的定殖風險。江蘇省內甘薯腐爛莖線蟲群體是否還存在其他不同基因型,各基因型腐爛莖線蟲生態適應性、寄主范圍、生物學特性等是否有差異,還需進一步調查研究。
參考文獻:
[1]劉維志. 植物病原線蟲學 [M]. 1版.北京: 中國農業出版社, 1999.
[2]周忠,馬代夫. 甘薯莖線蟲病的研究現狀和展望[J]. 園藝與種苗, 2003, 23(5):288-290.
[3]朱秀珍,田希武, 王隨保,等. 甘薯莖線蟲病發病規律及綜合防治[J]. 山西農業科學, 2004, 32(3):54-57.
[4]丁再福,林茂松. 甘薯、馬鈴薯和薄荷上的莖線蟲的鑒定[J]. 植物保護學報, 1982, 9(3):169-172.
[5]陳品三,鄭經武. 當歸麻口病中致病莖線蟲的鑒定研究[J]. 植物保護, 1988, 14(6):12-14.
[6]HOOPER D J. Ditylenchus destructor CIH description of plant parasitic nematodes No.21[M].UK:CAB International Wallingford, 1973.
[7]YAGHOUBI A, POURJAM E, YE W, et al. Description and molecular phylogeny of Ditylenchus gilanicus n. sp. (Nematoda: Anguinidae) from northern forests of Iran[J]. European Journal of Plant Pathology,2018,152(3):735-746.
[8]王金成,季鐳,黃國明,等. 腐爛莖線蟲不同地理群體ITS區序列比對及系統發育[J]. 河北農業大學學報, 2007, 30(5): 79-83.
[9]黃健,戚龍君,王金成,等. 腐爛莖線蟲種內不同群體形態及遺傳分析[J].植物病理學報,2009,39(2):125-131.
[10]章淑玲,張紹升. 甘薯莖線蟲rDNA-ITS1區的PCR擴增與序列分析[J]. 植物病理學報, 2008, 38(2):132-135.
[11]宛菲,彭德良,楊玉文,等. 馬鈴薯腐爛莖線蟲特異性分子檢測技術研究[J].植物病理學報, 2008, 38(3):263-270.
[12]SUBBOTIN S A,MOHAMMAD D A,ZHENG J Z,et al. Length variation and repetitive sequences of internal transcribed spacer of ribosomal RNA gene, diagnostics and relationships of populations of potato rot nematode, Ditylenchus destructor Thorne, 1945 (Tylenchida: Anguinidae)[J]. Nematology, 2011, 13(7):773-785.
[13]JESZKE A, BUDZISZEWSKA M, DOBOSZ R, et al. A comparative and phylogenetic study of the Ditylenchus dipsaci, Ditylenchus destructorand and Ditylenchus gigas populations occurring in Poland[J]. Journal of Phytopathology, 2014, 162(1):61-67.
[14]劉晨,楊藝煒,王家哲,等. 陜西不同地區馬鈴薯腐爛莖線蟲的分離鑒定及同源性分析[J]. 西北農業學報, 2020, 29(5):793-800.
[15]劉中華,林志堅,李華偉,等.甘薯種質資源遺傳多樣性的ISSR分析[J].南方農業學報,2019,50(11):2392-2400.
[16]趙冬蘭,周志林,唐君,等. 不同基因型甘薯的耐瘠性研究[J]. 江蘇農業科學,2019,47(3):74-78.
[17]張紹升.植物線蟲病害診斷與治理[M]. 福州: 福建科學技術出版社, 1999.
[18]ZHANG S L, LIU G K, JANSSEN T, et al. A new stem nematode associated with peanut pod rot in China: morphological and molecular characterization of Ditylenchus arachis n. sp. (Nematoda: Anguinidae) [J].Plant Pathology,2014,63(7):1197-1206.
[19]DARRIBA D, TABOADA G L, DOALLO R, et al. jModelTest 2: more models, new heuristics and parallel computing[J]. Nature Methods, 2012, 9(8): 772.
[20]XIA X, XIE Z. DAMBE: software package for data analysis in molecular biology and evolution[J]. Journal of Heredity, 2001, 30(7): 1720-1728.
[21]QIAO K, BAI M, HE Y, et al. Description of Labrys fuzhouensis sp.n. and first record of Coslenchus rafiqi (Nematoda: Tylenchidae) from China[J]. Nematology, 2019, 21(7):693-708.
[22]RONQUIST F, HUELSENBECK J P. MrBayes 3: bayesian phylogenetic inference under mixed models[J]. Bioinformatics, 2003, 19(12): 1572-1574.
[23]FELSENSTEIN J. Evolutionary trees from DNA sequences: a maximum likelihood approach[J]. Journal of Molecular Evolution, 1981, 17(6): 368-376.
[24]STAMATAKIS A, HOOVER P, ROUGEMONT J. A rapid bootstrap algorithm for the RAxML web servers[J]. Systematic Biology, 2017, 57(5): 758-771.
[25]THORNE G. Ditylenchus destructor n.sp. the potato rot nematode, and Ditylenchus dipsaci (Kühn, 1857) Filipjev, 1936, the teasel nematode (Nematoda: Tylenchidae).[J].Proceedings of the Helminthological Society of Washington, 1945, 12(2) :27-33.
[26]GOODEY J B. Investigations into the host ranges of Ditylenchus destructor and D. dipsaci[J]. Annals of Applied Biology, 1952,39(2):221-228.
[27] 張紹升,章淑玲,王宏毅,等. 甘薯莖線蟲的形態特征[J]. 植物病理學報, 2006, 36(1):22-27.
[28]WU L Y. Comparative study of Ditylenchus destructor thorne, 1945 (Nematoda:Tylenchidae), from potato, bulbous iris, and dahlia, with a discussion of de Man’s ratios[J]. Canadian Journal of Zoology, 1960, 38(6):1175-1187.
[29]JONES B L. First report of Ditylenchus destructor in pods and seeds of peanut [J]. Plant Disease, 1988, 72(5):453-453.
[30]于海英,彭德良,胡先奇,等. 馬鈴薯腐爛莖線蟲28S rDNA-D2/D3區序列分析[J]. 植物病理學報, 2009, 39(3):254-261.
[31]王宏寶,毛佳,李茹,等. 不同地理來源腐爛莖線蟲種群雜交后代致病力測定[J]. 華北農學報, 2011, 26(6):212-216.
[32]丁中,彭德良,高必達. 不同地理種群甘薯莖線蟲乙酰膽堿酯酶對抑制劑的敏感性[J]. 植物保護學報, 2007, 34(4):410-414.
[33]丁中,彭德良,何旭峰,等. 不同地理種群甘薯莖線蟲對不同類型殺線劑的敏感性[J]. 農藥, 2007, 46 (12):63-65.
[34]徐振,孫厚俊,楊冬靜,等. 甘薯莖線蟲在南方薯區的發生風險及影響因素研究[J]. 廣西農學報, 2017, 32(3):26-29.
(責任編輯:陳海霞)
收稿日期:2021-04-07
基金項目:國家現代農業產業技術體系建設專項(CARS-10-B15);徐州市農業科學院基金項目(RC2019002)
作者簡介:陳晶偉(1993-), 男, 陜西漢中人,碩士,研究實習員,從事甘薯病蟲害防治研究。(Email)ibcjw0825@126.com
通訊作者:孫厚俊,(Email)sunhouj1980@163.com
