栽培方式對寒地水稻產量及光合特性的影響
2021-01-18 04:21:00郭曉紅蘭宇辰王鶴瓔徐令旗孫光旭姜紅芳呂艷東
中國土壤與肥料 2020年6期
郭曉紅,蘭宇辰,胡 月,王鶴瓔,徐令旗,孫光旭,姜紅芳,呂艷東
(黑龍江八一農墾大學農學院/黑龍江省現代農業栽培技術與 作物種質改良重點實驗室,黑龍江 大慶 163319)
黑龍江省是我國最北部的寒地稻作區,也是我國重要的商品糧基地。黑龍江省水稻的高產穩產對我國糧食安全具有重要影響[1]。水稻產量主要來源在于其光合產物,而水稻群體質量和產量形成取決于光合物質的積累、分配、輸出和轉化是否合 理[2]。吳培等[3]研究認為提高水稻群體質量有利于水稻高產。吳桂成等[4]研究認為,提高水稻在生育前期和中期干物質積累量,對水稻增產有重要作用。迄今為止,關于水稻物質生產特性的研究,前人已有大量報道,并認為其受品種特性、栽插密度、氮素營養、插秧方式和種植方式[5-9]等因素的影響。然而,以上研究大都針對某一單因素對水稻的影響,通過栽培技術的集成與優化使水稻達到高產或超高產的群體物質生產研究相對較少。前期研究結果表明,高產高效和超高產栽培在保證水稻高產的同時,可以顯著提高各器官及全株的氮、磷、鉀含量和積累量,有利于寒地水稻養分的高效吸收與利用[10]。為此,本試驗比較了不同栽培方式下葉面積指數、光合特征參數、群體生長特征、干物質積累分配與轉運等方面的影響,為構建寒地水稻高產栽培技術體系提供理論與實踐 依據。
1 材料與方法
1.1 試驗地點及供試材料
試驗于2017~2018年在黑龍江省綏化市綏棱縣上集鎮水稻綜合試驗站(E127°18′,N47°09′)大田條件下進行。該地區屬于典型的寒溫帶大陸季風氣候區,年均降水量為545 mm,年蒸發量在700~800 mm,年平均氣溫為 2.59℃,氣象數據由位于試驗田內的農業氣象站測定。供試土壤為黑土,試驗前0~20 cm土層基礎理化性質為:有機質44.51 g·kg-1,堿解氮160 mg·kg-1,有效磷10.62 mg·kg-1,速 效 鉀185.84 mg·kg-1,pH 6.3。種植制度為一年一作。
供試水稻品種為龍粳46號。
1.2 試驗設計
試驗共設3個處理,分別為當地農民栽培方式(FP)、高產栽培方式(HY)和超高產栽培方式(SHY)。小區面積200 m2(20 m×10 m),每個處理3次重復,隨機排列。各處理間主要栽培措施和肥料運籌如表1所示。N肥以基肥∶分蘗肥∶調節肥∶穗肥=4∶3∶1∶2的比例施入,P肥作為基肥一次性施入,K肥以基肥∶穗肥=6∶4的比例施入,納米硅復合肥和生物有機肥為基肥一次性施入。基肥、分蘗肥、拔節肥、穗肥分別于移栽前12 d(2017年5月3日和2018年5月1日)、返青期(2017年5月20日和2018年5月19日)、8.5葉(2017年6月28日和2018年6月26日)和10.5葉(2017年7月8日和2018年7月6日)施用。化學肥料分別為尿素(N 46%)、磷酸二銨(N 18%,P2O546%)、硫酸鉀(K2O 50%),納米硅復合肥(有效硅≥55%)、生物有機肥(N+P2O5+K2O≥5%;有機質≥40%)。試驗期間各處理水分管理單排單灌,防止相互影響。播種日期分別為2017年4月11日、2018年4月9日,移栽日期分別為2017年5月15日、2018年5月13日,收獲日期分別為2017年9月28日、2018年9月27日。

表1 不同栽培方式下肥料運籌及栽培措施
1.3 測定內容與方法
1.3.1 干物質積累
分別在分蘗盛期、拔節期、齊穗期、成熟期,每小區按平均莖蘗數取代表性植株10穴,分蘗盛期和拔節期分為葉片、莖鞘2部分,齊穗期和成熟期分葉片、莖、鞘、穗4部分,置于烘箱105 ℃下殺青30 min,80 ℃下烘至恒重,測定各器官干物 質量。
1.3.2 葉面積
分別在分蘗盛期、拔節期、齊穗期,根據平均莖蘗數取代表性植株4穴,采用長寬系數法[10](葉長×葉寬×0.7)測定各時期葉面積,在齊穗期測定劍葉、倒2葉、倒3葉、余葉的葉面積,并計算群體生長特征,各指標計算公式[11]如下:
(1)高效葉面積率(%)=有效莖蘗上3葉總葉面積/有效莖蘗總葉面積×100
(2)表觀輸出率(%)=[表觀輸出量/齊穗期葉(莖、鞘)干重]×100
(3)表觀轉化率(%)=[表觀輸出量/成熟期籽粒干重]×100
(4)光合勢(104m2·d·hm-2)=(L1+L2)× (t1-t2)/2,式中,L1和L2為前后2次測定的葉面積。
(5)群體生長率(g·m-2·d-1)=(W2-W1)/ (t2-t1),式中,W1和W2為前后2次測定的干物 質重。
1.3.3 光合特征參數
在齊穗期葉片全展時,選擇晴朗無風、光照穩定的天氣,于9:00至11:00用CIRAS-3型光合儀(美國)測定劍葉中部凈光合速率(Pn)、氣孔導度(Gs)、胞間二氧化碳濃度(Ci)和蒸騰速率(Tr),重復4次。
1.3.4 產量及產量構成因素
收獲時各小區根據平均成穗數取10穴,考查每穗粒數、結實率、千粒重等性狀。選擇長勢均勻的4點,割取5 m2水稻,脫谷、曬干、風選后測定實際產量。
1.4 數據處理與統計方法
用Excel 2016進行數據處理,用DPS 7.05進行數據統計分析,采用LSD法進行多重比較(P<0.05)。
2 結果與分析
2.1 不同栽培方式對寒地水稻產量及其構成因素的影響
由表2可知,2年試驗產量及產量構成因素變化趨勢基本一致。不同栽培方式下水稻產量存在顯著差異,以SHY處理產量最高,2017年和2018年實際產量較FP處理分別增加24.78%和23.54%,差異達顯著水平(P<0.05)。其次是HY處理。從產量構成因素來看,SHY處理較FP處理水稻穗數、穗粒數均顯著增加,2018年結實率顯著降低;HY處理較FP處理水稻穗數、穗粒數均顯著增加,但2017年結實率和千粒重差異不顯著。SHY和HY處理增產的主要原因是由于穗數和穗粒數的增加。

表2 不同栽培方式下水稻產量及其構成因素的比較
2.2 不同栽培方式對寒地水稻葉面積指數的影響
由表3可知,不同栽培模式下水稻葉面積指數存在明顯差異。從分蘗盛期至齊穗期,各處理葉面積指數隨著生育進程的推進而增加,趨勢均表現為SHY>HY>FP,且在各時期SHY和HY處理與FP處理間的葉面積指數差異均達顯著水平。齊穗期上三葉葉面積指數均表現為SHY>HY>FP的趨勢,各處理間倒2葉葉面積指數均高于劍葉和倒3葉。HY和SHY對水稻高效葉面積率有顯著影響,2年間分別較FP平均顯著提高6.31%和9.76%。可見,HY和SHY在各時期能夠保證較高的葉面積指數,并且可以有效提高水稻齊穗期高效葉面積率,從而增大水稻群體葉源生長 優勢。

表3 不同栽培方式下水稻葉面積指數及齊穗期高效葉面積率的變化
2.3 不同栽培方式對水稻劍葉光合特征參數的 影響
不同栽培方式下水稻劍葉光合特征參數變化如表4所示,2年間光合特征參數變化趨勢較為一致。不同栽培方式下水稻凈光合速率、氣孔導度和蒸騰速率存在顯著差異,變化趨勢表現為HY>SHY>FP,胞間CO2濃度則呈現相反趨勢。HY處理2年間凈光合速率、氣孔導度和蒸騰速率分別較FP平均顯著提高28.49%、59.72%和23.84%。SHY處理2年間凈光合速率、氣孔導度和蒸騰速率分別較FP平均顯著提高11.91%、31.75%和12.85%。

表4 不同栽培方式下齊穗期水稻劍葉光合特征參數變化
2.4 不同栽培方式對寒地水稻光合勢和群體生長率的影響
由圖1可知,在不同栽培方式下,各生育階段光合勢在移栽至分蘗盛期最小,其次是分蘗盛期至拔節期,拔節期至齊穗期呈現最大值,呈現逐漸增加的趨勢。2年間在HY和SHY下各生育階段的光合勢較FP平均分別顯著提高30.77% ~ 62.29%、26.14% ~ 46.68%和13.71% ~ 22.94%;各生育階段群體生長率均表現為SHY>HY>FP的變化趨勢,且在拔節期至齊穗期群體生長率最高,在移栽至分蘗盛期、分蘗盛期至拔節期和拔節期至齊穗期,各處理之間群體生長率差異顯著,從齊穗期至成熟期,HY和SHY群體生長率較FP高,但差異并不顯著。
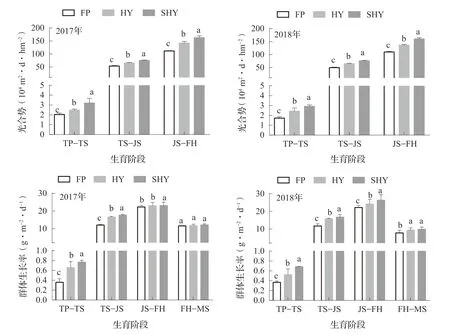
圖1 不同栽培方式下水稻光合勢和群體生長率的變化
2.5 不同栽培方式對寒地水稻群體干物質積累的影響
不同栽培方式下水稻群體干物質重變化如圖2所示,2年間在各時期群體干物質重均在SHY下為最高,其次是HY,HY和SHY在分蘗盛期、拔節期、齊穗期和成熟期群體干物質重平均分別較FP顯著提高50.0% ~ 90.0%、38.7% ~ 45.8%、15.3% ~ 24.7%和14.9% ~ 22.9%。HY、SHY處理尤其是拔節期之前,群體干物質重顯著提高,為水稻高產或超高產搭建良好的前期物質基礎。
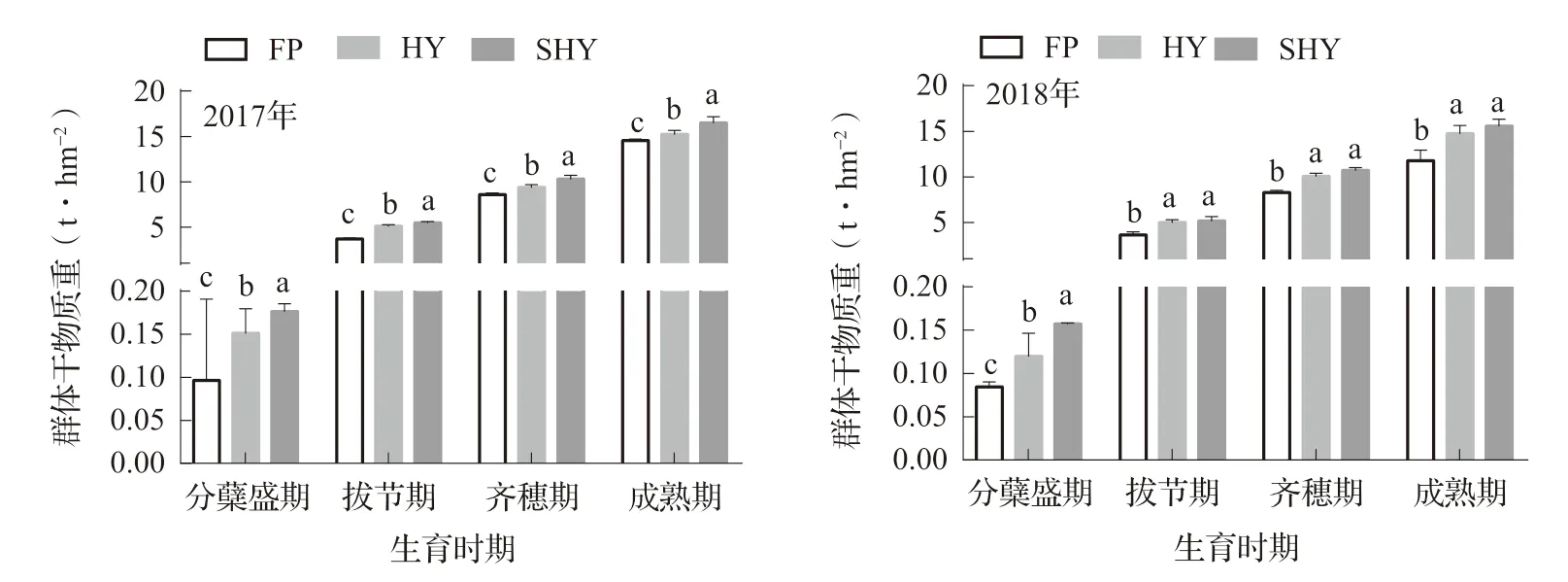
圖2 不同栽培方式水稻群體干物質重變化
2.6 不同栽培方式對寒地水稻干物質轉運的 影響
由表5可知,不同栽培方式對水稻抽穗前貯藏物質的轉運存在明顯差異。水稻葉、莖、鞘干物質輸出率、轉化率在各處理間均表現為鞘>葉>莖,各器官的輸出率、轉化率均表現為 SHY>HY>FP的趨勢,且SHY均顯著高于其它2種栽培方式。但各處理間莖的輸出率和2018年轉化率均為負值,說明各處理均出現干物質二次充實回升現象。由此可見,不同栽培方式下,HY和SHY均能促進水稻干物質向籽粒中轉運,尤其是鞘向籽粒中的轉運。
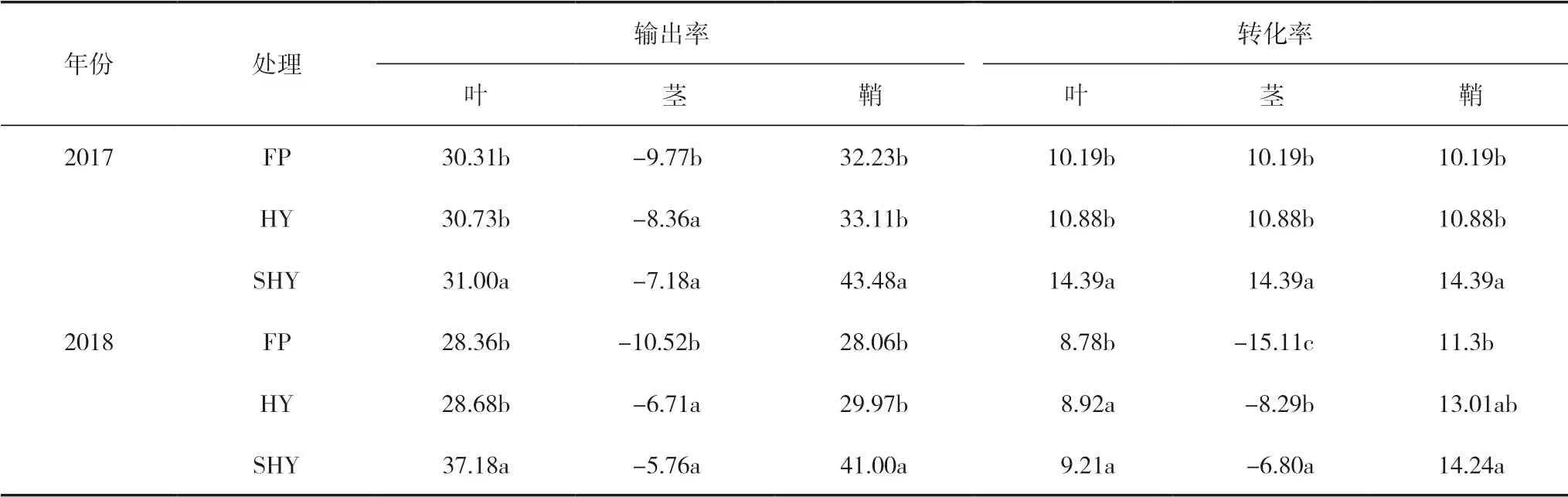
表5 不同栽培方式下水稻葉、莖、鞘干物質的輸出和轉化 (%)
2.7 不同栽培方式下葉面積指數、光合物質生產與產量的關系
由表6可得,不同栽培方式下水稻拔節期和齊穗期葉面積指數、齊穗期凈光合速率、各階段群體生長率及拔節-齊穗期光合勢整體上與成熟期干物質重、有效穗數、每穗粒數及產量呈顯著或極顯著正相關,而與結實率和千粒重整體呈負相關,但差異不顯著。

表6 不同栽培方式下葉面積指數、光合物質生產與產量的相關系數
3 討論
本研究結果表明,與農民栽培方式相比,高產栽培方式和超高產栽培方式均能通過提高穗數和穗粒數來實現產量的提高,尤以超高產栽培方式增產最高。此研究結果與前人研究結果相符。有研 究表明,適當減少基肥氮用量,增加蘗肥和穗肥氮用量等氮后移措施,可提高分蘗數,增加成穗率和每穗實粒數[12-13]。另有研究表明,有機無機肥配施能夠滿足水稻生育前期的速效養分的需求,促進有效穗數的增加,進而使水稻產量增加[14-15]。除此,薛亞光等[16]也認為水稻增產增效栽培和超高產栽培均是通過穗數增加來提高產量。本試驗中高產栽培方式和超高產栽培方式較農民栽培方式相比,除增加了蘗肥和穗肥氮用量外,還增施了生物有機肥和復合肥,且超高產栽培方式又采用了缽苗擺栽及寬窄行栽培技術,這些措施的集成與優化是水稻獲得高產的基礎。
水稻能進行有效的光合物質生產是實現高產的重要保障。光合物質生產是水稻有機物質的來源,同時也是水稻產量形成的重要基礎。葉面積指數、光合速率、光合勢和群體生長率等作為反映水稻群體光合生產能力的重要指標,尤其是葉面積指數可以反映光合源數量的多少,與產量密切相關。張洪程等[17]研究認為較大的光合勢和群體生長率能夠增加水稻群體干物質積累量,從而保證群體質量較高。邢智鵬等[18]研究認為水稻群體生長率變化為生育前期高、后期低,在齊穗期達到峰值,成熟期直線下降。楊惠杰等[19]對水稻在超高產栽培條件下的研究也得出相同的結論。本研究結果表明,在高產栽培和超高產栽培方式下,水稻各時期光合勢、凈光合速率、群體生長率(分蘗盛期-拔節期、拔節期-齊穗期)均顯著高于當地農民栽培方式,說明增施基肥生物有機肥和復合肥,增加蘗肥和穗肥氮用量,以及采用缽苗擺栽及寬窄行栽培技術,能夠有效改善水稻生育后期田間通風透光條件,個體生長潛力得到充分發揮,因此,二者均有較高的葉面積指數,增強了植株葉片的光合能力,促進了光合性能,為后期達到高效物質生產奠定了基礎,這與前人研究結果基本一致。
水稻的光合速率不僅受品種自身特性影響,外界因素和生長環境對水稻光合速率也有影響[20]。本研究結果表明,高產栽培和超高產栽培方式在齊穗期的凈光合速率、氣孔導度和蒸騰速率均較當地農民栽培方式有顯著提高,胞間CO2濃度則顯著降低。有研究表明[21],胞間CO2濃度值越低,濃度差就越大,導致較多的二氧化碳進入到氣孔當中,從而提高光合速率,即較低的胞間CO2濃度可以提高光合速率,這與本研究結果基本一致。
水稻干物質生產的80% ~ 90%都來自于光合作用,且水稻群體質量和產量形成也取決于光合物質的積累、輸出和轉化是否合理。抽穗后至成熟期的干物質積累量是衡量水稻群體質量的重要指標之一[22]。吳桂成等[4]研究認為,生育前期和中期保持較高的群體干物質量,是保證水稻高產的關鍵。薛亞光等[23]研究認為,高產栽培和超高產栽培方式可以顯著提高水稻從齊穗至成熟期的群體干物質量。張洪程等[17,24]研究也指出,在超高產栽培方式下,提高拔節期至成熟期干物質積累量對促進水稻高產有關鍵作用。本研究結果表明,在整個生育過程中,通過栽培措施的集成與優化,均能有效提高各階段干物質積累量,尤其是齊穗后群體干物質積累量,整個生育期中,齊穗至成熟期是水稻群體干物質積累量迅速提升的階段,隨著成熟期水稻群體干物質量的增加,產量水平顯著提高。由此可知,在保證生育前期干物質生產量適宜范圍的基礎上,通過提高齊穗后高效物質生產量,保證水稻在齊穗后干物質持續生產,以高效率的物質生產形成高質量的水稻高產群體,是高產栽培和超高產栽培方式提高物質生產能力和產量的物質基礎。
籽粒灌漿是產量形成的直接過程,灌漿物質來自抽穗后的光合產物以及葉、莖、鞘中貯藏物質的分配。“庫大、源足”是水稻高產的重要基礎[25],干物質輸出與轉化是衡量水稻源庫關系是否協調的重要指標[26]。龔金龍等[27]研究表明,高產水稻在生育前期葉、莖、鞘中貯藏的光合產物多,則在后期能較多的向穗部運轉。多項研究表明[28-29],高產水稻群體的莖鞘干重在灌漿后期會出現二次增重,即莖鞘干物重在抽穗后仍有回升的現象。本研究結果表明,在不同栽培方式下,莖干物質輸出量均為負值,說明莖干物質在后期光合產物仍有所積累,而鞘的干物質則始終向外輸出,莖、鞘干物質輸出率和物質輸出轉化率在高產栽培和超高產栽培方式中有所提高,尤其是超高產栽培方式尤為顯著。由此可知,在高產栽培方式和超高產栽培方式中,水稻的葉、莖、鞘向穗部輸出更多的光合產物。本研究中水稻各器官輸出率及物質輸出轉換率在高產栽培和超高產栽培方式得到顯著提高,有利于源庫流關系的協調,在擴大庫容的基礎上,又保證了“源”的供給,達到了更高水平的庫源平衡,從而提高產量。
4 結論
綜上所述,不同栽培方式下水稻產量、光合物質生產特性均存在顯著差異。高產栽培方式和超高產栽培方式提高了葉片光合性能,顯著提高了物質生產能力,促進物質由源向庫的轉化,從而提高了寒地水稻產量,為寒地水稻高產栽培方式提供理論支持。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
軍事文摘(2021年22期)2021-11-26 00:43:51
河北農機(2020年10期)2020-12-14 03:13:30
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
今日農業(2019年15期)2019-01-03 12:11:33
聚氯乙烯(2018年9期)2018-02-18 01:11:34
