羌塘高原藏羚羊棲息地分布及影響因素
2021-01-16 04:28:08魏子謙徐增讓
生態學報 2020年23期
魏子謙,徐增讓
1 中國科學院地理科學與資源研究所, 北京 100101
2 中國科學院大學, 北京 100049
藏羚羊(Pantholopshodgsonii)是《國家重點保護野生動物名錄》確定的國家一級保護野生動物,《瀕危野生動植物種國際貿易公約》[1]附錄I物種,2016年被國際大自然保護聯盟(IUCN)瀕危野生動物保護紅色目錄列為近危(NT)[2],在中國、印度和尼泊爾都有分布,目前主要生活在以羌塘高原為中心的青藏高原高寒荒漠區[3-4]。早在更新世藏羚羊就已生存于青藏高原[5],在長達二百萬年的進化中,從生理特征、生活習性都適應了這里獨特且嚴峻的自然條件,形成按季節遷徙和繁殖規律,被公認為青藏高原動物區系的典型代表[2, 6]。藏羚羊多分布在海拔4500m以上的高寒荒漠生態系統。棲息地覆被類型多為荒漠草原、高山植被,部分為高寒草甸和沼澤,植被群落結構簡單,植株低矮,蓋度低[7-8]。禾本科(Poaceae)、莎草科(Cyperaceae)及豆科(Leguminosae)植物為藏羚羊主要食譜構成[9-10],其中禾本科植物占采食總量的60%,包括紫花針茅(Stipapurpurea)、扇穗茅(Lizzledacaracmosa)、早熟禾(Poaannua)、昆侖嵩草(Kobresializledalea)等[11]。
藏羚羊是羌塘高原有蹄類中唯一的遷徙物種。成年藏羚羊一年中除交配季節,絕大部分時間雌雄分群。每年6月份,雌性藏羚羊逐漸集結并向夏季產羔區遷徙,6月20日至7月10日小羊羔陸續出生,完成生產后雌性藏羚羊于7月上旬回遷,8月返回越冬棲息地[12-13]。羌塘高原的藏羚羊總體呈現由南向北遷徙趨勢。不同區域種群遷徙路線不同,如羌塘西部的藏羚羊多集結至昆侖山南麓,甚至翻越昆侖山至新疆阿爾金山產羔;東部藏羚羊一路北上,朝可可西里方向遷徙;南部藏羚羊則僅向附近僻靜的河流寬谷遷徙小段距離等待產羔[14-15]。每年藏羚羊需在越冬區度過3/4的時間,越冬區多為距離水源較近、植被蓋度較高、可食植物豐富、溫暖濕潤的草原或河湖灘地[16]。產羔區的食物資源和氣候條件都相對較差,但人類和其他野生動物干擾較少,僻靜安全[17-18]。
人類活動是影響藏羚羊棲息地選擇的主要因素[19],因其天生警覺性強,在選取棲息地時會明顯避讓人類活動區域。近年,隨著野生動物保護工作不斷增強,藏羚羊種群數量恢復性增長,對棲息地及水草資源需求不斷擴張[9]。同時,羌塘地區社會經濟發展、放牧生產、道路交通和城鎮建設等人為干擾日益加劇[20],直接或間接侵占了藏羚羊棲息地,其中交通道路會破壞原有棲息地的連通性,并延緩藏羚羊遷徙行為[12, 21],牧業生產將導致草場資源競爭,另外闖入承包草場的藏羚羊會招致牧民驅逐[20]。已有研究采用樣點、樣線等野外調查法對物種棲息地的分布范圍開展調查[4, 12- 14, 17, 19, 22- 24],取得一些珍貴的資料。但藏羚羊分布范圍廣,棲息地可進入性差[22],野外調查難度大,其棲息地數據相對零散,給藏羚羊種群保護帶來困難。
論文以羌塘高原為研究區,把握藏羚羊生活習性,刻畫棲息地選擇偏好特征。采用樣線法收集藏羚羊出現點位數據,利用Maxent(maximum entropy model)模型按藏羚羊遷徙習性分別模擬其繁殖與非繁殖兩季棲息地范圍。考慮城鄉聚落、交通道路、牧業生產等主要人類活動,基于Habitat Quality模型模擬人為活動對棲息地干擾。識別藏羚羊在不同季節的棲息地分布和受人類活動干擾區域,助力相關部門明確關鍵保護地域、科學調整保護區功能分區,維持遷徙通道連通性,以提高野生動物保護的針對性和有效性。
1 研究方法與數據來源
1.1 研究區
羌塘高原藏語意為“北方的曠野”,指岡底斯—念青唐古拉山脈以北,昆侖山脈以南的廣闊地區,行政上屬西藏自治區的那曲與阿里兩地區管轄,是青藏高原的重要組成部分。區域內平均海拔約4500m以上,地勢高亢、開闊坦蕩、起伏平緩,植被類型簡單,蓋度低[8, 25]。野生動物是羌塘高原上最活躍也是最引人注目的部分,十余種國家一級保護動物生活在這片土地上,其中藏羚羊作為草食性有蹄類的典型代表,在羌塘高原具有得天獨厚的優勢,截至2015年,羌塘高原的藏羚羊估計約達20萬只,約占世界種群數量70%[20, 26-27]。
1.2 研究方法
1.2.1樣線法采集野生動物出現點樣本
與當地野生動物保護部門座談、走訪當地居民、查閱現有資料等確定藏羚羊在不同季節的大致棲息地范圍。于2019年5月和8月,開展野外樣線調查,采集羌塘高原藏羚羊出現點位樣本。根據地理、水草資源布設樣線,沿樣線以10—20km/h車速,通過肉眼觀察輔以視得樂8倍夜視雙筒望遠鏡,記錄觀測點坐標、距藏羚羊距離和相對角度,計算得到藏羚羊出現點信息。通過樣線法共記錄到38個藏羚羊出現點位數據,另外從野生動物保護學會(WCS)青藏高原有蹄類動物現狀及趨勢報告[28]和相關文獻[3, 6, 15]中獲取部分2000年以后藏羚羊出現點位坐標,對其數字化,通過遙感影像和生境信息對比校正,獲取了129個藏羚羊出現點位作為補充。將所有樣點通過ArcGIS緩沖區分析法篩選,以0.5km作為緩沖半徑,若1km范圍出現多點則僅保留1個,以降低樣本空間信息冗余度。
在模擬棲息地時,按遷徙繁殖時間可將藏羚羊棲息地分為暖季產羔區和冷季越冬區。為此,將每年5月下旬至7月末,從藏羚羊開始遷徙至完成產羔后帶領新生幼崽回遷至越冬區這一過程中途經和停留的位置(包括該時期部分沒有遷徙行為的雌、雄性藏羚羊的活動位置)作為藏羚羊的產羔點;將每年除了遷徙繁殖月份以外其他時間棲息、活動點位作為越冬點。最終獲得羌塘高原藏羚羊夏季產羔點61個,越冬點106個,如圖1所示。
1.2.2Maxent模擬棲息地分布
最大熵模型Maxent[29]基于物種出現點位及其對應的環境變量模擬物種棲息地的分布,廣泛應用于野生動物保護研究,是目前模擬與預測能力最強的物種分布模型[30- 33]。它假設物種分布是根據物種分布的概率p確定的,p的大小反映了物種對環境因子偏好程度。Maxent在所有滿足約束條件的模型中,選擇熵值最大(即分布最均勻)的結果預測物種分布[29]。
X∈{x1,x2,…,xn},y為隨機離散變量,X的概率分布為p(xi),i=1,2...,n,y的條件熵為:
(1)
最大熵特征根f(xi,y)為約束條件下的最優化問題的解[34]:
(2)
(3)
模型中的變量y為物種分布可能性大小,X為一系列環境變量,包括氣候、地貌、植被等。研究以藏羚羊實測出現點位(圖1)為訓練和測試樣本,據藏羚羊的生理習性和棲息地選擇偏好,選擇了多個與藏羚羊分布直接相關的環境背景因子,為降低共線性,防止模型過度擬合,對所有環境變量進行相關性分析,排除相關性較強且與藏羚羊棲息地選擇偏好聯系較弱的環境變量,最終得到12個指標(表1),包括7個氣候因子:Bio1年均溫度、Bio2平均日較差、Bio10最暖季平均氣溫、Bio11最冷季平均氣溫、Bio12年降水量、Bio18最暖季降水量、Bio19最冷季降水量;2個地理因子:海拔、坡度,和3個食物水源因子:植被類型、植被蓋度和距水源距離。利用Maxent 3.4.1[35]對藏羚羊的夏季產羔區和越冬區分別進行模擬。模型運行時隨機抽取20%樣本作為測試數據集,其余80%樣本作為訓練數據集。使用刀切檢驗(Jackknife)測定各環境變量對藏羚羊棲息地分布的貢獻,創建環境變量響應曲線。依據AUC值(area under ROC curve,受試工作特征曲線與橫坐標圍成的面積)進行模型精度評價[36]。Maxent模型推薦的評價標準為:測試AUC=0.6—0.7模擬效果較弱,0.7—0.8一般,0.8—0.9良好,大于0.9為優秀。
1.2.3InVEST生境質量模塊模擬人類活動對生境的威脅
綜合考慮草地放牧、城鄉聚落、交通道路等主要人類活動(表1),采用InVEST3.5.0 Habitat Quality模塊,就人類活動對棲息地質量的影響進行定量分析。一般人類活動指數超過0.09的區域屬于城鎮空間,低于0.02的區域為受人類干擾較弱區域。人類活動強度指數(Dxj)計算見公式4[37]:
(4)
其中:wr為第r種威脅的權重,ry為源于柵格y處的第r種威脅水平;irxy為r威脅從威脅源y到柵格x的衰減率;βx為易受擾動水平;Sjr為棲息地j對r威脅的敏感性。
為便于空間分析,主要威脅因子包括城鎮聚落用地(urb)、鄉村聚落用地(rr)、耕地(crp)、公路(prds)、鄉村路(srds)。根據各種威脅的相對強度,權重依次取1、0.6、0.7、1和0.6,最大影響距離依次取10、8、6、8、6 km。威脅出現,威脅水平取1,未出現取0。易受擾動水平取1。棲息地對各種威脅的敏感性取[0,1]。
1.3 數據來源
藏羚羊點位數據和環境背景數據分別如圖1和表1所示。
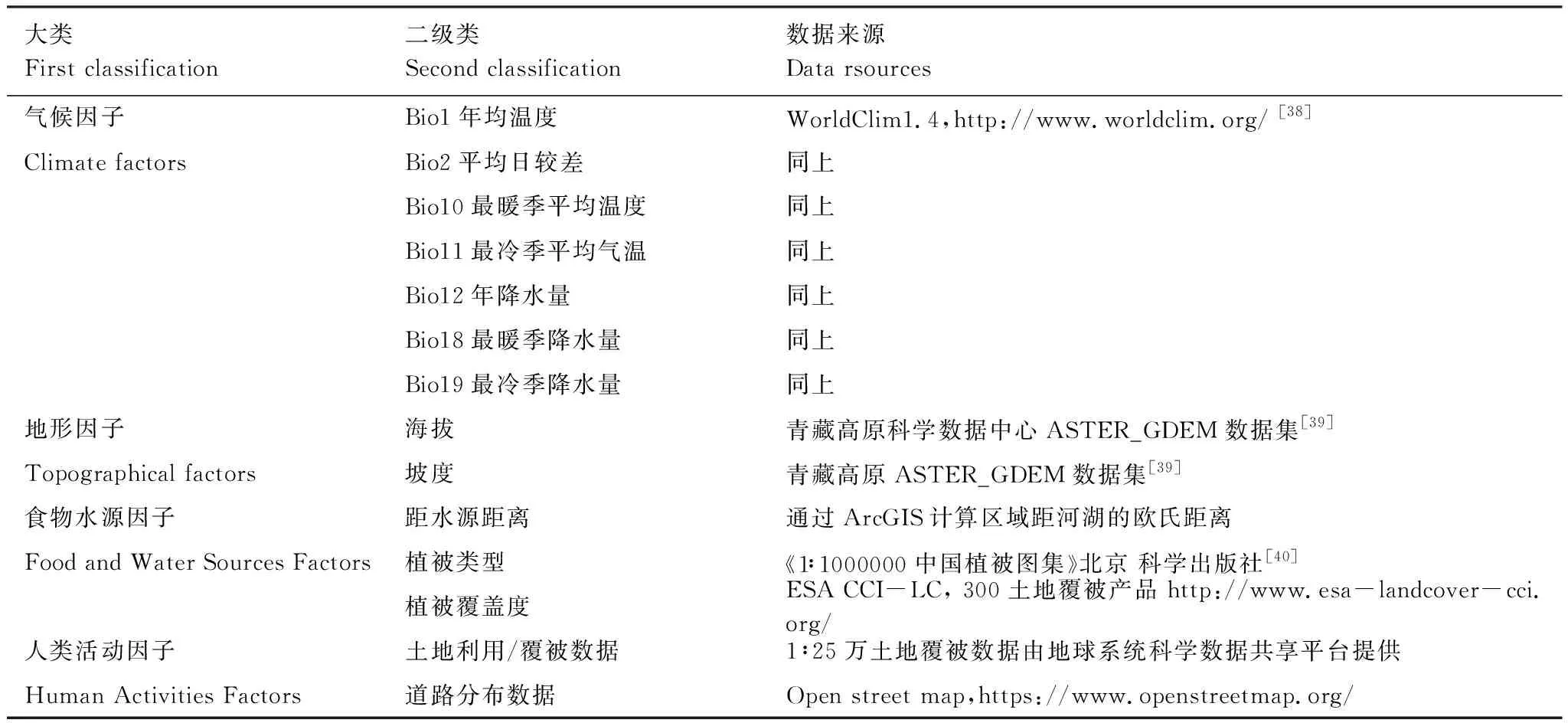
表1 藏羚羊棲息地分布影響因子及數據來源Table 1 Influence variables and their sources
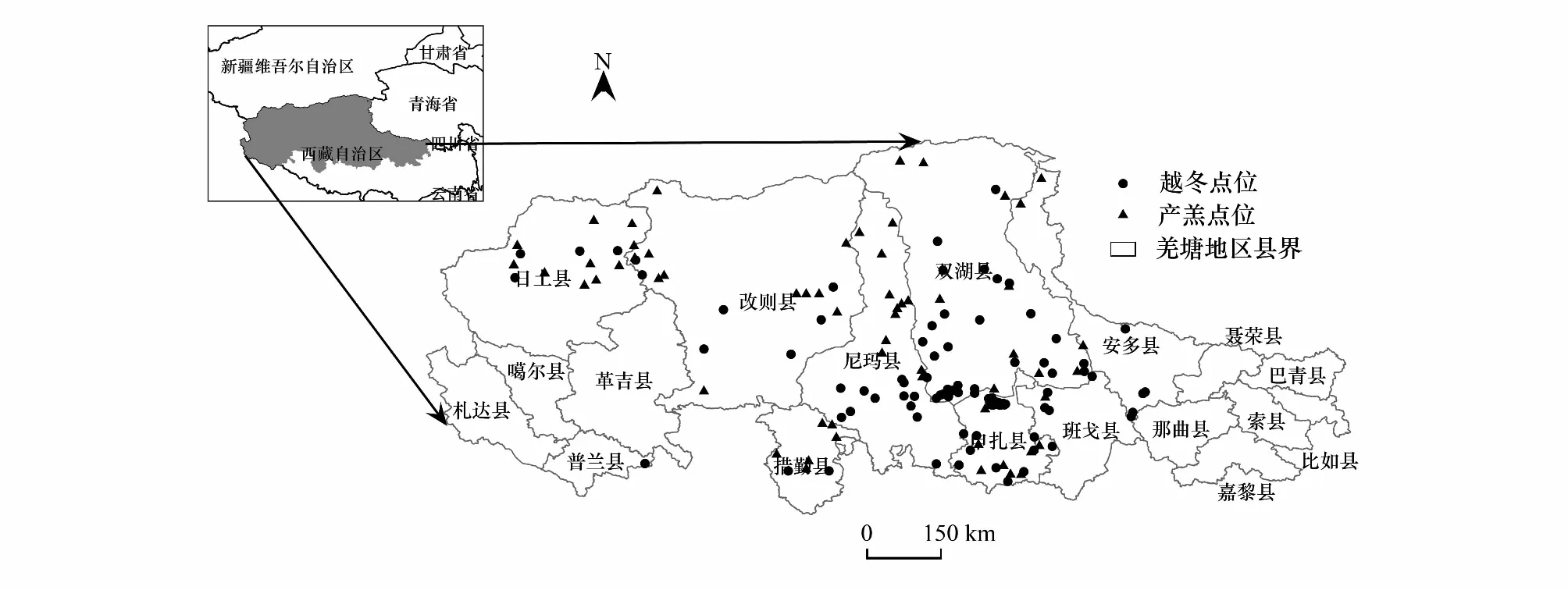
圖1 藏羚羊點位數據Fig.1 Tibetan antelope point data
2 研究結果
2.1 棲息地分布現狀
根據藏羚羊不同月份已知點位數據和相關環境因子,利用Maxent分別模擬其在非繁殖季節(越冬區)和繁殖季節(產羔區)的分布情況。模擬顯示越冬區測試AUC=0.834,產羔區測試AUC=0.805,評級均為良好[29, 36],說明該模型在模擬藏羚羊兩季棲息地方面具有較高可信度。
結合羌塘地區野生動物保護部門的巡查數據,當地牧戶訪談結果和已有文獻記錄,將Maxent運算結果區分為“潛在棲息地”和“非潛在棲息地”,并對潛在棲息地的重要等級進行閾值劃分。設定藏羚羊潛在越冬區的出現概率的閾值為大于0.1,如圖2所示。設定夏季產羔區出現概率的閾值為大于0.15,如圖3所示。
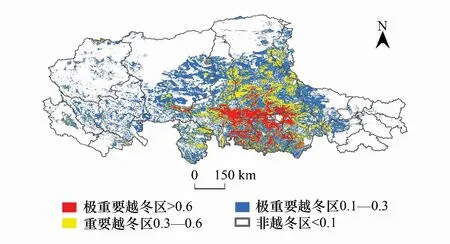
圖2 藏羚羊越冬區分布Fig.2 Distribution of Tibetan antelope over-winter area
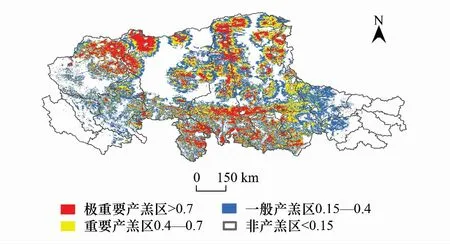
圖3 藏羚羊產羔區分布Fig.3 Distribution of Tibetan antelope lambing area
藏羚羊越冬區主要分布在羌塘高原東南部,集中于那曲地區的西南部及阿里部分地區,越冬區面積約為263984km2,占研究區的38.3%。其中重要和極重要越冬區面積分別為66801.6km2和32802.8km2,集中分布在雙湖、尼瑪和申扎三縣交界處,圍繞色林錯、吳如錯和達則錯,另外在阿里地區改則縣、措勤和日土也有部分分布。產羔區相對于越冬區則明顯呈現向北部擴散的趨勢,成片狀分布在日土縣東北部、改則縣西北、那曲地區東南部和北部區域,面積約為304848km2,占研究區的44.2%,重要和極重要產羔區面積分別為111526km2和79084km2,分布相對分散,在北部和南部水系附近多分布有重要產羔區。
2.2 棲息地分布的自然影響因素
通過環境因子的響應曲線可以看出藏羚羊在選擇越冬區時,傾向于選擇海拔大于4800m,年均溫度高于-5℃,坡度小于10°,年降水量大于200mm,植被類型為紫花針茅、珠峰苔草高寒草原,紫花針茅、墊狀駝絨藜高寒草原和苔狀蚤綴與墊狀點地梅墊狀植被,水母雪蓮、鳳毛菊高山稀疏植被,距水源5km以內、食物充足,溫暖,地勢平緩,植被蓋度相對高的區域。夏季產羔區則傾向于選擇植被類型為墊狀駝絨藜高寒荒漠、紫花針茅高寒草原、墊狀駝絨藜高寒草原、青藏苔草高寒草原、小嵩草高寒草甸、嵩草沼澤化高寒草甸,另外坡度小于10°,海拔大于4800m,平均氣溫日較差11—16℃,距水源10km以內,最暖季降水量小于300mm的區域。
在影響藏羚羊棲息地選擇的12個環境因子中,植被類型、距水源距離、坡度、Bio2(平均氣溫日較差)和Bio18(最暖季降水量)是決定產羔地分布的最主要因素,累計貢獻值達94.4%。影響藏羚羊越冬區選擇的環境因素貢獻值相比產羔區更加分散,植被類型和Bio11(最冷季平均氣溫)貢獻值突出,分別為36.3%和25.2%,Bio18(最暖季降水量)、Bio12(年降水量)、坡度、Bio10(最暖季平均氣溫)、Bio1(年均氣溫)也較重要。由兩類棲息地環境因子的貢獻值差異(表2)可以看出藏羚羊在選擇越冬區時更偏好食物充足,植被覆蓋度較高并且氣候溫暖的區域,屬于資源偏好。相比之下,藏羚羊在遷徙選擇產羔地時則更加注重水源、坡度、海拔、氣溫日較差等,對植被資源的選擇傾向較越冬區弱,更加重視遷徙通道連貫性和產羔區安全性。
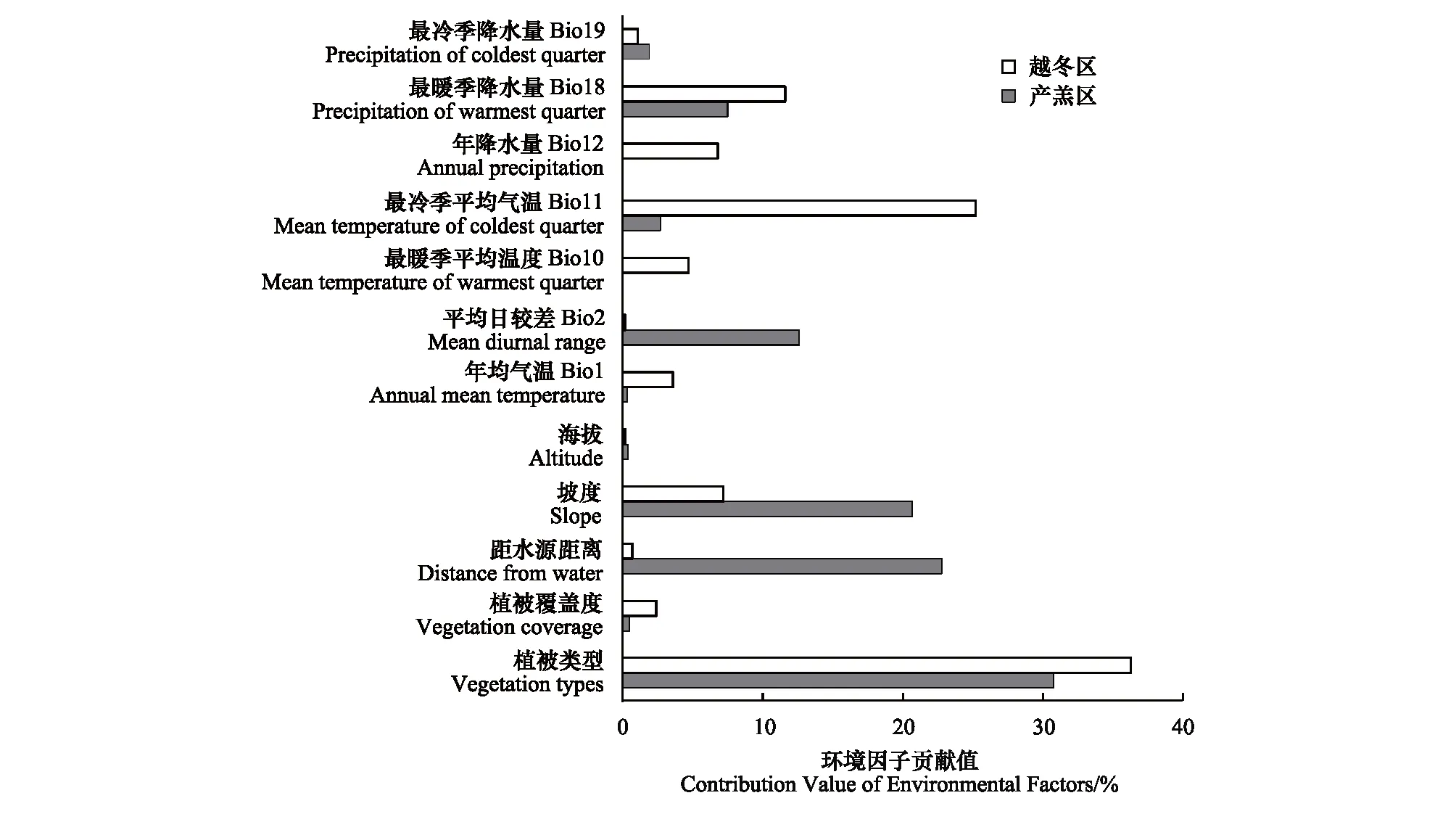
圖4 兩類棲息地環境因子貢獻值Fig.4 Contribution value of environmental factors in two kind of habitats
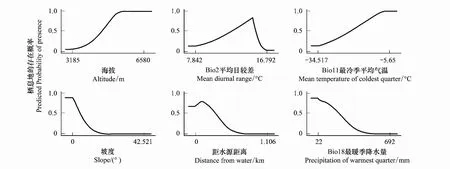
圖5 產羔區主要環境變量響應曲線Fig.5 Response curves of habitat variables in Maxent models
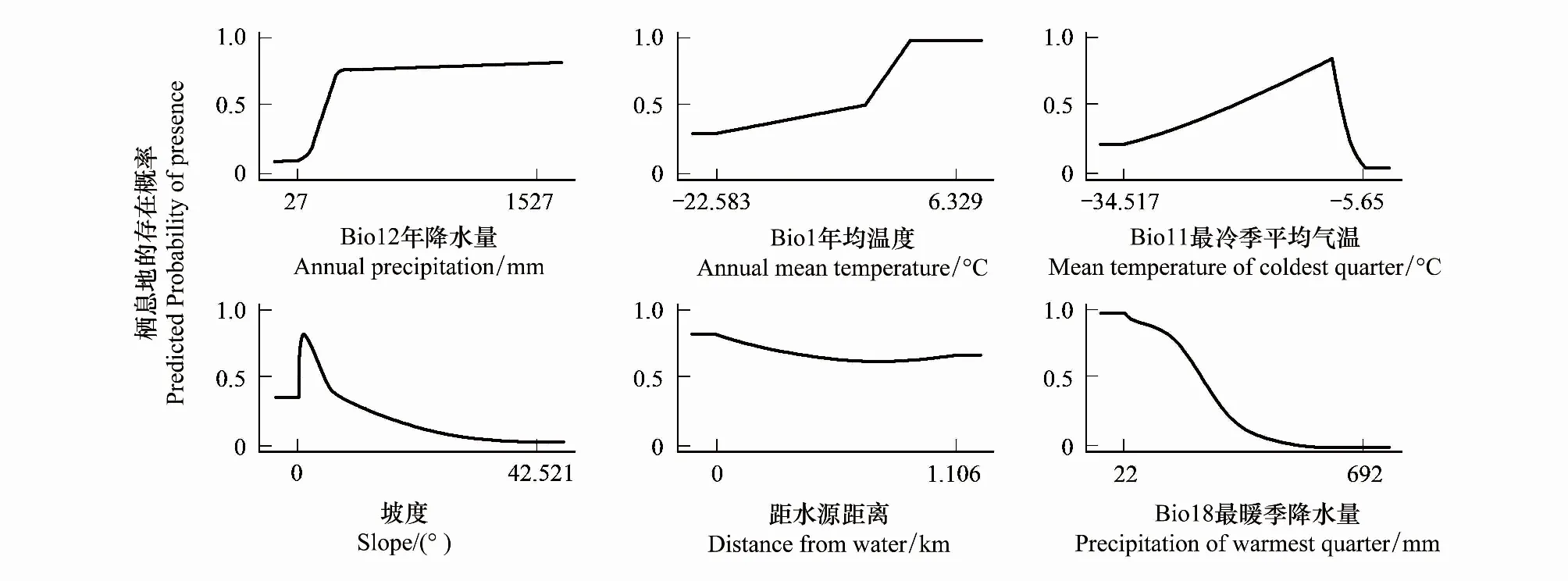
圖6 越冬區主要環境變量響應曲線Fig.6 Response curves of habitat variables in Maxent models
2.3 人類活動對棲息地的影響
隨著羌塘地區社會經濟的發展,棲息地受到多種人類活動干擾,致使其適宜性下降。采用Habitat Quality模擬了人類活動對棲息地的影響強度指數。選取人類活動強度指數0.025作為閾值,小于該閾值人類活動對藏羚羊干擾較弱。以0.025、0.03、0.04為分界點將人類活動劃分為“較弱、一般、較強、強”四個等級,如圖7所示。雖然羌塘高原整體人類活動水平較低,但是在那曲地區南部、阿里地區東南和西南部仍有部分人類活動較活躍區域。
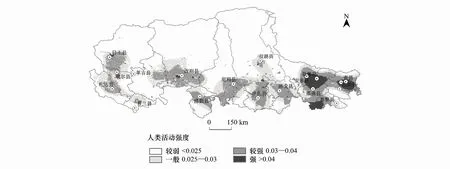
圖7 人類活動強度分區Fig.7 Division of human activity intensity
將人類活動強度指數大于0.025的區域與藏羚羊產羔區和越冬區疊加(圖8):發現受到人類活動干擾的越冬區面積為104763km2,占越冬區39.7%;受到人類活動干擾的產羔區面積為106341km2,占產羔區的34.9%;羌塘高原有195651km2區域同時作為藏羚羊繁殖季節和非繁殖季節棲息地,其中80161km2受到了人類活動的干擾,主要分布在那曲地區班戈縣、申扎縣,尼瑪縣東南部和雙湖縣南部,以及阿里地區東南局部,干擾類型多為居民地、放牧生產和交通旅游業等人類活動。
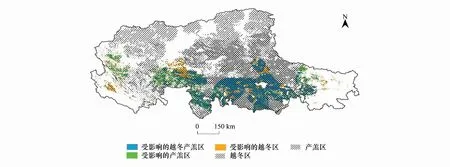
圖8 受人類活動干擾的棲息地范圍Fig.8 Habitat disturbed by human activities
3 討論
以往研究表明,藏羚羊主要分布在羌塘高原的高寒荒漠地帶,棲息地植被蓋度低,多見于河道寬谷、盆地或湖濱平原地帶[7, 15]。羌塘高原上已被學者和野生動物保護部門證實的藏羚羊“產房”昆侖山南麓、瑪依崗日以東以及雙湖縣江愛藏布江畔上的羚羊渡江集結地等[3,6, 12, 14, 17],基本都被本研究識別為藏羚羊重要、極重要產羔區。重要越冬區色林錯、友誼湖周邊灘地等[14-15]也被識別為藏羚羊越冬區。環境變量響應曲線顯示了藏羚羊對植被類型(禾本科、莎草科)的偏好,尤其是對紫花針茅和小蒿草的選擇傾向,表明羌塘地區藏羚羊與新疆阿爾金山[9]和青海可可西里地區[10-11]藏羚羊的食性基本相同。李維東等人通過對阿爾金山藏羚羊產羔區的研究發現該區域植被不僅覆蓋度低,且適口性和營養價值都較差,認為藏羚羊在遷徙過程中對食物偏好較其他時期弱[18],本研究發現羌塘藏羚羊在資源選擇方面也具有相似特征。
藏羚羊遷徙行為與棲息地季節分布。藏羚羊遷徙規律較復雜,長距離遷徙行為常見于雌性藏羚羊[12],生活在不同區域的種群遷徙方向、距離各不相同。劉務林等1987—2005年通過追蹤不同種群,發現羌塘地區雌性藏羚羊表現出三種遷徙模式,部分個體整個夏天都待在越冬區,沒有長距離遷徙行為;部分個體僅遷移一小段距離,到達夏季產羔區;一些雌性群體會從越冬區出發,向北遷移較長的距離[15]。雄性藏羚羊在6、7月份也存在遷徙行為,但往往距離較短。雌、雄性以及不同群體的藏羚羊遷徙行為差異較大,很難準確界定產羔區和越冬區。本文僅以時間作為節點劃分和模擬藏羚羊的越冬區和產羔區,反映了藏羚羊在不同季節的大致活動范圍。尤其是產羔區中包括了一定數量的不遷徙藏羚羊群體。
人類活動對藏羚羊棲息地的影響。羌塘地區南部居民地、牧業和道路等人為活動明顯干擾了藏羚羊棲息地,尤其以在色林錯為中心的藏羚羊棲息地與人類活動區域存在較多空間重疊。部分區域建設網圍欄切斷了藏羚羊的水路、草路和遷徙通道[20]。較強的人類活動致使藏羚羊棲息地質量下降。對受到干擾的棲息地,要優化保護格局、維持遷徙通道的連通性,結合地區實際出臺相應準入清單,分地段、分時段,協調牧民、游客等與藏羚羊的關系。同時,隨著藏羚羊恢復性增長及對水草資源需求增加,其頻繁造訪牧民承包的草場。目前仍無相關辦法對草食性野生動物占用草場資源進行補償,牧民排斥甚至驅逐藏羚羊事件時有發生,亟待量化藏羚羊造成的牧草采食,以完善野生動物肇事補償體系。維持草地生態平衡、促進野生動物保護和牧業生產協調發展。
4 結論
研究采用野外調查與物種分布模型相結合的辦法,基于藏羚羊棲息地選擇偏好,明確在繁殖和非繁殖季節的棲息地分布,并分別識別兩個時期受人類活動干擾的區域,以助力野生動物保護工作和人與野生動物關系協調。結果表明:藏羚羊在非繁殖季節較為集中的分布在羌塘高原東南部,圍繞在色林錯等水系周圍,其越冬區面積約為26萬km2。傾向選擇海拔4800m以上、氣候溫暖、靠近水源且食物資源豐富的區域。藏羚羊在繁殖季節棲息地明顯呈現向北部擴散的趨勢,多在水系周圍呈小片狀分布于羌塘東北、西北和南部區域,產羔區面積約為30萬km2。選擇產羔區時則更加注重坡度、水源、海拔、氣溫日較差等,對植被資源的選擇傾向較非繁殖季弱,更重視遷徙通道連貫性和產羔區安全性。受人類活動影響的棲息地主要在那曲地區南部和阿里西南部,受到干擾的棲息地分別占越冬區的39.7%,產羔區的34.9%。
猜你喜歡
少先隊活動(2022年5期)2022-06-06 03:45:04
家庭科學·新健康(2022年3期)2022-05-10 00:32:13
哈哈畫報(2022年4期)2022-04-19 11:11:50
大科技·百科新說(2021年6期)2021-09-12 02:37:27
中老年保健(2021年2期)2021-08-22 07:31:10
英語文摘(2021年2期)2021-07-22 07:56:54
少先隊活動(2021年1期)2021-03-29 05:26:36
快樂語文(2020年30期)2021-01-14 01:05:38
好孩子畫報(2020年5期)2020-06-27 14:08:05
意林·全彩Color(2019年6期)2019-07-24 08:13:50
