鯉CCAAT 增強(qiáng)子結(jié)合蛋白α 基因的克隆與時(shí)空表達(dá)特征
2021-01-15 02:07:04孫志鵬呂偉華黨紅陽(yáng)曹頂臣匡友誼魯翠云鄭先虎
水產(chǎn)學(xué)雜志 2020年6期
孫志鵬 ,呂偉華,黨紅陽(yáng),2,曹頂臣,匡友誼,魯翠云,鄭先虎
(1.中國(guó)水產(chǎn)科學(xué)研究院黑龍江水產(chǎn)研究所,淡水魚(yú)類育種國(guó)家地方聯(lián)合工程實(shí)驗(yàn)室,黑龍江 哈爾濱 150070;2.上海海洋大學(xué),農(nóng)業(yè)農(nóng)村部淡水水產(chǎn)種質(zhì)資源重點(diǎn)實(shí)驗(yàn)室,上海 201306)
CCAAT 增強(qiáng)子結(jié)合蛋白 (CCAAT enhancer binding protein,C/EBP) 是堿性區(qū)亮氨酸拉鏈(basic region/leucine zipper,Bzip)蛋白家族的一個(gè)轉(zhuǎn)錄因子[1]。C/EBP 轉(zhuǎn)錄因子包含一個(gè)高度保守的DNA 結(jié)構(gòu)域和一個(gè)亮氨酸拉鏈二聚體結(jié)構(gòu)域,形成同源或異源二聚體結(jié)合到相似的序列上[2]。其中,C/EBPα 在脂肪細(xì)胞、骨髓細(xì)胞、肝臟、肺及胎盤等多種細(xì)胞或組織中均有表達(dá)[3-5],具有調(diào)控細(xì)胞分化、參與能量代謝、抑制細(xì)胞增殖等多種生物學(xué)功能[6],脂肪細(xì)胞分化過(guò)程中控制動(dòng)物腹腔脂肪沉積的主效基因或與主效基因緊密連鎖。Lekstrom 和Xanthopoulos[7]發(fā)現(xiàn):C/EBPα 基因的過(guò)表達(dá)能加速前脂肪細(xì)胞向成熟脂肪細(xì)胞分化,在大多數(shù)特異性脂肪細(xì)胞基因轉(zhuǎn)錄之前表達(dá)。Yeh 等[8]和Rosen 等[9]揭示,C/EBPα 在脂肪細(xì)胞分化的終末階段大量表達(dá),與過(guò)氧化物酶體增殖劑激活受體(peroxisome proliferator activated receptor,PPARγ)共同作用,開(kāi)啟一系列脂肪特異性基因的表達(dá),合成、攝取和儲(chǔ)存長(zhǎng)鏈脂肪酸,使細(xì)胞停止增殖,呈現(xiàn)完全分化狀態(tài)。
目前,關(guān)于C/EBPα 與脂肪沉積關(guān)系的研究主要集中在哺乳動(dòng)物和畜禽方面。Wang 等[10]研究發(fā)現(xiàn):小鼠C/EBPα 基因表達(dá)受到干擾后,腹部脂肪沉積明顯下降。Lee 等[11]研究顯示,敲除C/EBPα 基因后,小鼠喪失了磷酸烯醇丙酮酸羧激酶(PEPCK)基因功能,影響糖代謝和脂肪生成。耿蓓等[12]研究表明,高脂飲食會(huì)抑制小鼠血液中C/EBPα 的表達(dá)。張愛(ài)朋等[13]比較了C/EBPα 突變?nèi)怆u與未突變個(gè)體發(fā)現(xiàn),突變個(gè)體腹脂含量及腹脂率顯著高于未突變個(gè)體。然而,C/EBPα 基因在魚(yú)類中卻鮮有報(bào)道。本研究通過(guò)克隆鯉Cyprinus carpio C/EBPα 基因全長(zhǎng)cDNA 序列,并將其編碼氨基酸序列與其他物種進(jìn)行了同源性多重比較及系統(tǒng)進(jìn)化分析,研究C/EBPα 基因在鯉不同組織中及其在不同胚胎發(fā)育時(shí)期的表達(dá)特征,以期為深入研究C/EBPα 基因在鯉脂肪細(xì)胞分化及脂肪發(fā)生過(guò)程中的作用機(jī)制提供參考。
1 材料與方法
1.1 材料
實(shí)驗(yàn)魚(yú)5 尾為1 齡魚(yú),體質(zhì)量200~250 g,取自中國(guó)水產(chǎn)科學(xué)研究院黑龍江水產(chǎn)研究所呼蘭試驗(yàn)站同一養(yǎng)殖池塘,禁食24 h 后,采集腹腔脂肪(脂肪組織)、腦組織、肝臟、心臟、脾臟、腎臟、腹部肌肉、腸組織和血液等樣本,迅速放入液氮中保存?zhèn)溆茫灰来尾杉幨芫哑凇? 細(xì)胞期、囊胚晚期、原腸胚晚期、7 體節(jié)期、心跳期、體循環(huán)期、出膜期、開(kāi)口期和出苗5 d 等10 個(gè)不同發(fā)育時(shí)期胚胎樣本,保存于液氮中備用。
1.2 基因全長(zhǎng)cDNA 獲得
參照RNeasy Min Elute Clean up Kit(QIAGEN)試劑盒說(shuō)明提取總RNA,使用Prime ScriptTMRT reagent Kit 試劑盒合成cDNA 第一鏈。從NCBI 檢索并下載斑馬魚(yú)等硬骨魚(yú)類C/EBPα 基因的cDNA 序列,選擇保守區(qū)域與鯉轉(zhuǎn)錄組文庫(kù)進(jìn)行比對(duì),獲得鯉C/EBPα 基因部分cDNA 序列,并設(shè)計(jì)同源簡(jiǎn)并引物(表1)。以反轉(zhuǎn)錄產(chǎn)物cDNA 為模板,PCR 產(chǎn)物純化后連接到PMD18-T(TaKaRa)載體,轉(zhuǎn)入DH5α 感受態(tài)細(xì)胞,選擇陽(yáng)性克隆送至蘇州金唯智生物科技有限公司測(cè)序,獲得C/EBPα 基因核心片段序列。參照SeqAmpTMDNA Polymerase(Clontech)試劑盒說(shuō)明進(jìn)行5’/3’端RACE 擴(kuò)增,以獲得非編碼區(qū)(UTR)序列。測(cè)序結(jié)果經(jīng)BLAST(https://www.n cbi.nlm.nih.gov/)比對(duì)、DNAMAN 分析和Staden(1.7)拼接,最終獲得C/EBPα 基因的全長(zhǎng)cDNA 序列[14]。
1.3 基因序列特征及系統(tǒng)進(jìn)化分析
利用ExPASy(https://www.expasy.org/proteomics)等在線軟件分析信號(hào)肽、跨膜結(jié)構(gòu)、蛋白二級(jí)結(jié)構(gòu)、蛋白疏水性及脂肪系數(shù)等蛋白特征。使用MEGA6.0軟件,采用鄰接法(Neighbor-Joining method,NJ)構(gòu)建系統(tǒng)進(jìn)化樹(shù),Bootsrap 值取1 000 次,由JTT 距離矩陣模型(JTT matrix-based method)計(jì)算進(jìn)化距離[ 15 ]。
1.4 基因表達(dá)特征分析
參照測(cè)序后獲得的C/EBPα 基因全長(zhǎng)cDNA 序列,設(shè)計(jì)熒光定量引物(表1)。以反轉(zhuǎn)錄產(chǎn)物cDNA為模板,以鯉18S 為內(nèi)參基因,參照One Step SYBRPrime ScriptTMRT-PCR Kit II(TaKaRa)試劑盒使用方法,利用ABI 7500 Real-time PCR 定量?jī)x,測(cè)定了鯉不同組織間和不同胚胎發(fā)育時(shí)期C/EBPα 基因的相對(duì)表達(dá)量,每個(gè)樣品設(shè)3 個(gè)重復(fù)。采用2-△△Ct方法計(jì)算基因相對(duì)表達(dá)量。利用SPSS 17.0 單因素方差分析(one-way ANOVA)分析了基因相對(duì)表達(dá)量的差異顯著性,差異顯著性水平P<0.05,差異極顯著性水平P<0.01[16]。

表1 引物序列Tab.1 Primers used in this study
2 結(jié)果與分析
2.1 鯉C/EBPα 全長(zhǎng)cDNA 序列分析
采用RACE 擴(kuò)增方法獲得C/EBPα 基因的cDNA 全長(zhǎng)總長(zhǎng)為1 463 bp 的鯉C/EBPα(GenBank 登錄號(hào):MN073500)。其中,234 bp 的5’端非編碼區(qū)(5’-UTR),861 bp 的開(kāi)放閱讀框(編碼286 個(gè)氨基酸)和368 bp 的3’端非編碼區(qū)(內(nèi)含30 bp 的polyA尾巴),polyA 上游176 bp 處存在一個(gè)“ACTAAA”加尾信號(hào)。
經(jīng)ExPASy 的ProtParam 工具預(yù)測(cè)蛋白分子式為C1409H2210N426O438S13,總原子數(shù)為4 496 個(gè)。蛋白質(zhì)相對(duì)分子量為32.5 kD,理論P(yáng)I 值7.02;不穩(wěn)定系數(shù)為52.48,蛋白穩(wěn)定性較低;脂肪系數(shù)為63.50,親水性平均系數(shù)為-0.962。氨基酸預(yù)測(cè)分析表明,脯氨酸含量最高(8.0%),不含色氨酸、苯丙氨酸、絲氨酸;天冬酰胺和谷氨酰胺共帶負(fù)電荷殘基總數(shù)36個(gè),精氨酸和賴氨酸帶正電荷殘基總計(jì)35 個(gè)。
2.2 氨基酸序列多重比較與進(jìn)化樹(shù)構(gòu)建
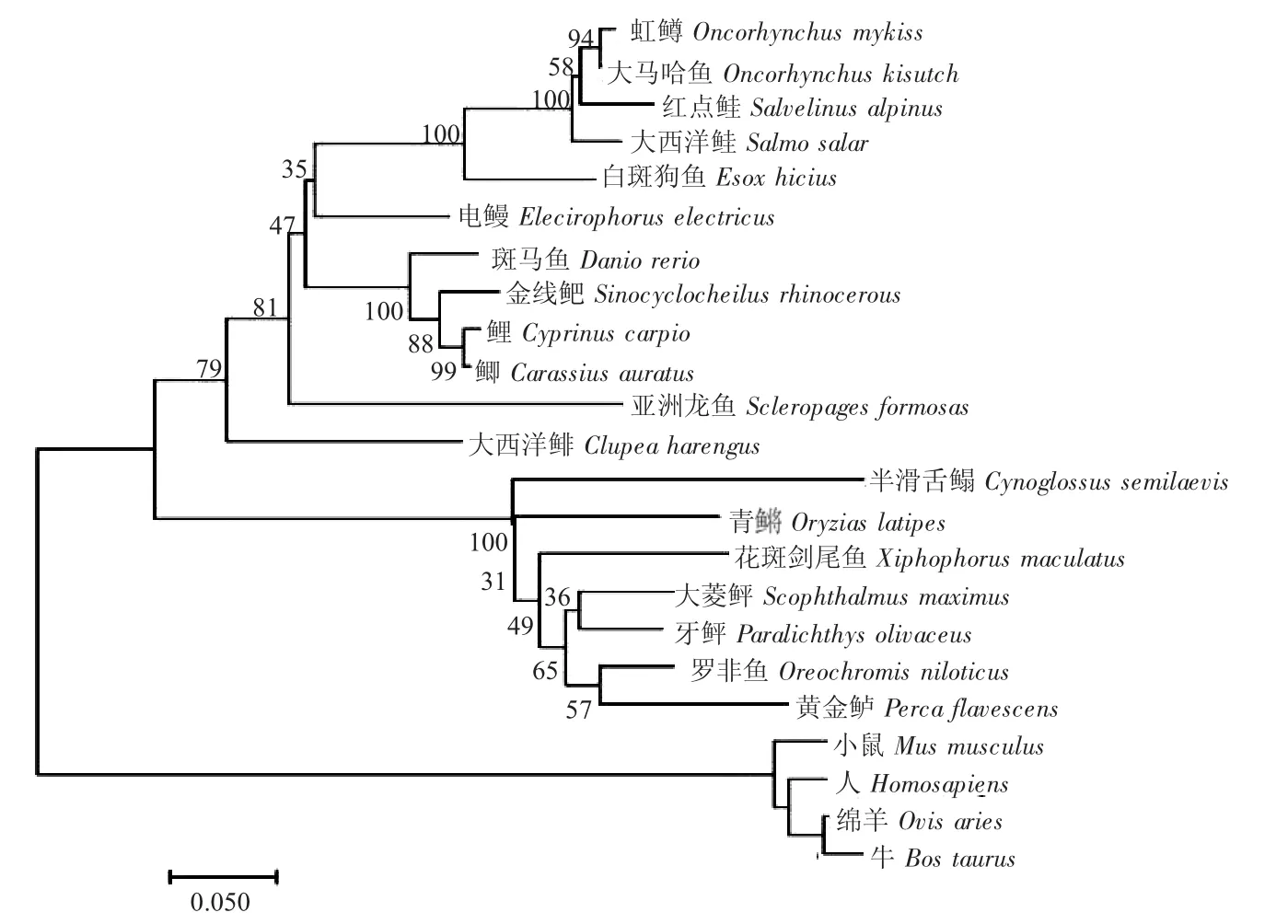
圖2 基于不同物種C/EBPα 氨基酸序列構(gòu)建的系統(tǒng)進(jìn)化樹(shù)Fig.2 Phylogenetic tree based on C/EBPα amino acid sequences of different species
利用CLUSTAL 2.1 在線分析軟件對(duì)鯉C/EBPα 氨基酸序列進(jìn)行比對(duì)。結(jié)果顯示,其與斑馬魚(yú)Danio rerio、大西洋鮭Salmo salar、小鼠Mus musculus 氨基酸序列的相似性分別為93.36%、63.64%和52.47%。多重比較結(jié)果顯示,6 個(gè)物種間C/EBPα 氨基酸序列同源性的差異主要集中在C-端的多肽上(圖1)。
基于23 個(gè)物種C/EBPα 氨基酸序列,采用MEGA 6 軟件,用N-J 法構(gòu)建系統(tǒng)進(jìn)化樹(shù)。結(jié)果顯示,全部魚(yú)類聚為一大支,其中鯉、鯽、斑馬魚(yú)等鯉科魚(yú)類聚為一支;虹鱒、大西洋鮭等鮭科類魚(yú)聚為一支;牙鲆Paralichthys olivaceus、半滑舌鰨Cynoglossus semilaevis 等鲆鰈類魚(yú)聚為一支,哺乳動(dòng)物聚為另一大支(圖2)。
2.3 C/EBPα 基因在鯉不同組織表達(dá)特征
實(shí)時(shí)熒光定量PCR 結(jié)果顯示,C/EBPα 基因在實(shí)驗(yàn)組織中均有表達(dá)(圖3),9 個(gè)組織中基因相對(duì)表達(dá)量由高到低依次為:脂肪、腸、腦、脾、肝、腎、心臟、血液和腹部腹肌。脂肪的相對(duì)表達(dá)量相對(duì)高于其他組織,且差異極顯著(P<0.01);腦、脾、肝和腎組織中的表達(dá)量相當(dāng),無(wú)顯著差異(P>0.05)。
2.4 C/EBPα 基因鯉不同發(fā)育時(shí)期表達(dá)特征
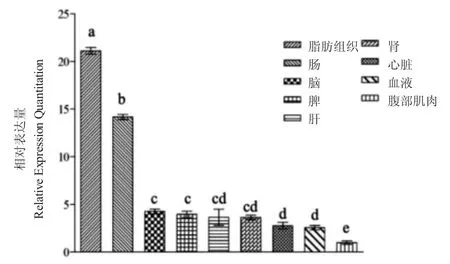
圖3 C/EBPα 基因在鯉不同組織中的相對(duì)表達(dá)量Fig.3 Relative expression levels of C/EBPα gene in different tissues of common carp C.carpio

圖4 鯉不同胚胎發(fā)育時(shí)期C/EBPα 基因的相對(duì)表達(dá)Fig.4 Relative expression levels of C/EBPα gene in different embryonic stage of common carp C.carpio
C/EBPα 基因在鯉胚胎不同發(fā)育時(shí)期表達(dá)量變化情況見(jiàn)圖4。由圖4 可知,受精后至心跳期,隨著胚胎的發(fā)育,C/EBPα 基因的表達(dá)量逐漸上升,心跳期表達(dá)量最高,相對(duì)其他時(shí)期差異極顯著(P<0.01)。心跳期后基因表達(dá)量顯著下降,出膜期后C/EBPα基因表達(dá)量有所升高。
3 討論
自1989 年被發(fā)現(xiàn)至今,大量預(yù)測(cè)研究表明,C/EBPα 蛋白也位于細(xì)胞核內(nèi),屬于b-Zip 家族,具有典型亮氨酸拉鏈結(jié)構(gòu)[17,18]。不同物種間C/EBPs 轉(zhuǎn)錄因子的DNA 結(jié)合域和激活域高度同源,尤其是DNA 結(jié)構(gòu)域在進(jìn)化過(guò)程中變異率極低。小鼠C/EBPα 基因編碼區(qū)長(zhǎng)度約1 188 bp,編碼395 個(gè)氨基酸[11],秦川牛基因編碼區(qū)長(zhǎng)度1 062 bp,編碼353 個(gè)氨基酸[19],雞C/EBPα 基因編碼區(qū)長(zhǎng)度約1 300 bp[20],朗德鵝基因編碼區(qū)長(zhǎng)度約975 bp,編碼324 個(gè)氨基酸[21]。本研究克隆了鯉C/EBPα 基因全長(zhǎng),開(kāi)放閱讀框861 bp,共編碼286 個(gè)氨基酸,編碼氨基酸數(shù)較哺乳動(dòng)物和畜禽類少。鯉C/EBPα 基因編碼氨基酸序列與斑馬魚(yú)相似度為93.36%,而與人相似度僅為52.47%,可見(jiàn)物種間差異較大。系統(tǒng)進(jìn)化分析結(jié)果顯示,哺乳動(dòng)物聚為一大支,魚(yú)類聚為一大支,鯉科魚(yú)類聚為一小支,說(shuō)明C/EBPα 基因在進(jìn)化過(guò)程中具有較高的保守性。盤道興等[22]研究表明,C/EBPα基因在豬皮下脂肪中表達(dá)量最高,肝和小腸中表達(dá)較高,心和腎中表達(dá)量最低,與本研究結(jié)果相近,說(shuō)明在豬和鯉組織中C/EBPα 基因表達(dá)量與脂肪細(xì)胞的實(shí)際分布成正比。
大多數(shù)魚(yú)類在饑餓狀態(tài)下優(yōu)先動(dòng)用脂肪作為能量來(lái)源[23,24],而且對(duì)脂肪的利用能力很高,其用于機(jī)體增重和分解供能的總利用率達(dá)90%以上[25]。C/EBPα 在脂肪細(xì)胞分化、發(fā)育過(guò)程中起決定作用,眾多脂肪細(xì)胞特異性表達(dá)的基因都含有C/EBPα 的結(jié)合位點(diǎn),可激活大部分脂肪細(xì)胞特異性表達(dá)基因,如磷酸烯醇丙酮酸羧基酶(PEPCK)基因、葡萄糖轉(zhuǎn)運(yùn)體(GLUT)-4 基因及瘦素(leptin)等的調(diào)控區(qū)域,從而啟動(dòng)相關(guān)基因的轉(zhuǎn)錄表達(dá),同時(shí)促進(jìn)細(xì)胞中脂肪的積累[26,27]。脂肪沉積主要通過(guò)脂肪細(xì)胞數(shù)量增多和體積增大來(lái)實(shí)現(xiàn)。韋璇等[28]發(fā)現(xiàn),C/EBPα 基因表達(dá)上調(diào)會(huì)促進(jìn)脂肪細(xì)胞體積增大,導(dǎo)致阿勒泰羊尾部脂肪沉積。然而對(duì)魚(yú)類脂肪性狀的研究,應(yīng)系統(tǒng)探究脂肪沉積、脂肪分解、脂肪酸的形成等,單純孤立研究某一個(gè)或兩個(gè)基因遠(yuǎn)遠(yuǎn)不夠,需要更多地發(fā)掘魚(yú)類的脂質(zhì)產(chǎn)生與代謝相關(guān)基因表達(dá)的差異,從整體上把握這些基因控制魚(yú)類脂肪性狀的相關(guān)性,清楚地了解魚(yú)類脂肪性狀的控制機(jī)制。
