大豆質(zhì)核互作雄性不育系花器官發(fā)育生理生化特性的變化
2021-01-07 07:32:58崔天宇曹霞孫佳堯李志剛劉鵬王鵬年
江蘇農(nóng)業(yè)科學(xué) 2021年24期
關(guān)鍵詞:大豆
崔天宇 曹霞 孫佳堯 李志剛 劉鵬 王鵬年



摘要:以大豆質(zhì)核互作雄性不育系JLCMS9A及其同型保持系JLCMS9B為試驗材料,測定花芽期、花蕾期、成花期中超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)的活性以及丙二醛(MDA)、淀粉、可溶性蛋白、可溶性糖、游離脯氨酸的含量,分析3個時期生長素(IAA)、赤霉素(GA3)、異戊烯基腺嘌呤核苷(iPA)、脫落酸(ABA)的含量及變化。結(jié)果表明,在花芽期不育系中的SOD、CAT活性以及MDA、游離脯氨酸的含量均高于保持系,而在花蕾期和成花期相反,其值均低于保持系;不育系的POD活性在花芽期顯著低于保持系,在花蕾期和成花期相反,其值均高于保持系;不育系9A的淀粉、可溶性糖含量整體呈上升趨勢,在花蕾期和成花期均低于保持系9B。花器官整個發(fā)育過程中,不育系9A的IAA含量呈先升后降的變化趨勢,iPA含量為逐漸升高趨勢,不育系各激素含量均低于保持系。不育系的IAA/ABA、IAA/GA3、IAA/iPA、ABA/GA3的比值與保持系存在差異。由此推斷,花器官發(fā)育生理特性的異常與大豆質(zhì)核互作雄性不育有關(guān),花蕾期可能是大豆質(zhì)核互作雄性不育系花器官生理生化指標發(fā)生異常的關(guān)鍵時期。
關(guān)鍵詞:大豆;質(zhì)核互作雄性不育;花器官;生理生化特性;抗氧化酶活性;內(nèi)源激素含量
中圖分類號:S565.101 文獻標志碼: A
文章編號:1002-1302(2021)24-0075-07
收稿日期:2021-04-07
基金項目:博士科研啟動基金(編號:BS514);內(nèi)蒙古自治區(qū)科技儲備項目(編號:2018MDCB02);內(nèi)蒙古自治區(qū)科技計劃(編號:2018KJJH1702);內(nèi)蒙古民族大學(xué)科研立項項目(編號:NMDSS2159)。
作者簡介:崔天宇(1996—),女,內(nèi)蒙古赤峰人,碩士,主要研究方向為作物遺傳育種。E-mail:2105842265@qq.com。
通信作者:李志剛,博士,教授,主要研究作物遺傳育種。E-mail:13948651158@126.com。
大豆原產(chǎn)于我國,迄今已有5 000多年的栽培史,因此我國有豐富的大豆資源。大豆作為主要糧食作物之一,是人類所需蛋白質(zhì)和食用植物油的主要來源。隨著我國經(jīng)濟的發(fā)展,各方面對大豆的需求也日益增加。但由于大豆的產(chǎn)量和經(jīng)濟效益較低,導(dǎo)致其種植面積低于玉米、水稻、小麥等主要糧食作物,我國大豆總產(chǎn)量明顯供不應(yīng)求。因此培育高產(chǎn)、優(yōu)質(zhì)、抗性強的大豆是我國大豆育種的首要目標,而實現(xiàn)這一目標最有效的措施就是利用雜種優(yōu)勢。大豆質(zhì)核互作雄性不育系的發(fā)現(xiàn)為大豆雜種優(yōu)勢的利用提供了基礎(chǔ)[1]。
近年來,對于棉花[2]、水稻[3]、油菜[4]、煙草[5]、白菜[6-8]、玉米[9]等生理生化特性的研究較多,而關(guān)于大豆不育系及其生理生化特性的研究卻鮮有報道。研究者們發(fā)現(xiàn)植物雄性不育與其器官的物質(zhì)代謝、能量代謝、內(nèi)源激素以及其抗氧化酶活性有關(guān)。雄性不育系體內(nèi)的物質(zhì)含量減少,代謝速度降低。研究指出抗氧化酶[超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)]具有清除植物代謝過程中產(chǎn)生的氧自由基的作用,可以保持質(zhì)膜的穩(wěn)定性[10],從而維持植物體內(nèi)活性氧的平衡。內(nèi)源激素與植物的不育性有關(guān),是調(diào)節(jié)其生長發(fā)育的重要因子[11-12]。
我國大豆雄性不育的主要類型有細胞核雄性不育[13-15]、質(zhì)核互作雄性不育[16-18]和光溫敏雄性不育3種[19-21]。本試驗以大豆質(zhì)核互作雄性不育系及其同型保持系作為試驗材料,通過測定花器官發(fā)育不同時期中的SOD、POD、CAT活性,丙二醛(MDA)、淀粉、可溶性蛋白、可溶性糖、游離脯氨酸的含量以及生長素(IAA)、赤霉素(GA3)、異戊烯基腺嘌呤核苷(iPA)、脫落酸(ABA)等內(nèi)源激素含量及其動態(tài)含量的變化,探究其與不育性的相關(guān)性,以期為選育大豆優(yōu)良不育系及不育機制研究提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
以吉林省農(nóng)業(yè)科學(xué)院大豆研究所提供的大豆不育系JLCMS9A與同型保持系JLCMS9B為試驗材料,下文分別用9A、9B表示。于2019年5月播種于內(nèi)蒙古民族大學(xué)北區(qū)試驗地,試驗設(shè)為3個小區(qū),每個小區(qū)的面積為20 m2,采用等行距相間種植,株行距為15 cm×60 cm,每個小區(qū)種植6行。試驗材料均在同一時期進行播種,且在生長期間采用相同的管理措施進行田間管理。
1.2 樣品采集
于盛花期開始取樣,對主莖第4節(jié)位以上的花芽、花蕾、成花分別進行取樣,重復(fù)3次,將樣品放入液氮進行速凍,然后放入-80 ℃冰箱保存。
1.3 測定項目與方法
可溶性蛋白含量采用考馬斯亮藍G-250染色法測定,可溶性糖及淀粉含量采用硫酸蒽酮比色法測定,游離脯氨酸含量采用酸性茚三酮法測定,CAT活性采用紫外吸收法測定,SOD活性采用氮藍四唑(NBT)光還原法測定,POD活性采用愈創(chuàng)木酚法測定,MDA含量采用硫代巴比妥酸(TBA)法測定。
采用間接酶聯(lián)免疫(ELISA)法分別測定不育系及同型保持系花器官不同發(fā)育時期生長素(IAA)、赤霉素(GA3)、異戊烯基腺嘌呤核苷(iPA)和脫落酸(ABA)的含量,3次重復(fù)。
1.4 數(shù)據(jù)分析
利用軟件DPS 16.05對測定指標數(shù)據(jù)間的差異顯著性進行分析,使用Excel 2016進行柱狀圖繪制。
2 結(jié)果與分析
2.1 花器官發(fā)育不同時期抗氧化酶活性的變化
如圖1所示,在花器官發(fā)育過程中,不育系9A的SOD活性呈下降趨勢,且在成花期達到顯著性差異;保持系9B的SOD活性則是先升高后降低,且花蕾期達到最高值(254.58 U/g),達到顯著性差異。在花蕾期、成花期,保持系9B的SOD活性分別是不育系9A的1.18、1.58倍,均達到顯著性差異。在成花期,不育系9A的SOD活性驟降,推測成花期可能是發(fā)生敗育的時期。
不育系9A及保持系9B的POD活性在花器官發(fā)育時期均呈先升高后降低的變化趨勢,且二者同時在花蕾期達到最高值,但差異不顯著。在花芽期9B的POD活性比9A高9.5%,差異顯著。成花期9B的POD活性大幅降低,是9A的94.5%,差異顯著。不育系與保持系的POD活性在成花期較花蕾期均顯著降低,但在整個發(fā)育時期不育系和保持系表現(xiàn)出相同的倒U形變化規(guī)律。因此,推測POD活性與大豆雄性不育相關(guān)性不大。
不育系9A和保持系9B的CAT活性呈不同的變化趨勢,9A呈逐漸降低的變化趨勢,而9B呈倒U形的變化趨勢。隨著花器官的發(fā)育,9A、9B成花期CAT的活性均低于花芽期,且達到了差異顯著水平;其中,在花芽期9A比9B高1.75倍,而在成花期9B比9A高2.03倍,說明隨著花器官的發(fā)育,CAT活性在逐漸降低,且不育系9A比保持系9B顯著性降低。從花芽期至成花期,不育系9A的MDA含量呈先降低后升高的變化趨勢,而保持系9B呈逐漸下降的變化趨勢。在花芽期、花蕾期、成花期,不育系9A與保持系9B均達到了顯著性差異,并且不育系9A的MDA含量分別是保持系9B的1.22、1.23、1.41倍。在成花期不育系9A與保持系9B的MDA 含量差距最大, 說明成花期MDA含量對育性影響較大。
2.2 花器官不同發(fā)育時期的營養(yǎng)物質(zhì)含量比較
由圖2可以看出,不育系9A、保持系9B的淀粉含量均呈遞增的變化趨勢,均在成花期達到最高值,分別為11.41、21.76 mg/g。在花器官發(fā)育過程中,9B的淀粉含量始終高于9A,且在花蕾期和成花期差異顯著。在花蕾期、成花期,9B的淀粉含量比9A分別高出52.4%、90.7%,說明隨著花器官的發(fā)育,淀粉含量對大豆育性起關(guān)鍵作用。由于后期淀粉虧缺,花器官發(fā)育缺少能量和營養(yǎng)物質(zhì),使花器官發(fā)育異常,導(dǎo)致雄性不育。
在整個花器官發(fā)育過程中,不育系9A和保持系9B的可溶性蛋白含量在花器官發(fā)育時期均呈逐漸下降趨勢,且不育系9A與保持系9B的可溶性蛋白含量均在花芽期達最大值,分別為157.25、169.86 mg/g,且達到了顯著性差異。隨著花器官的發(fā)育,可溶性蛋白含量逐漸降低,在成花期無顯著性差異;但在花芽期、花蕾期,保持系9B的可溶性蛋白含量分別是不育系9A的1.08、1.09倍,且達到了顯著性差異,說明花芽期、花蕾期可能是發(fā)生不育的時期。
從花芽期到成花期,不育系9A和保持系9B的可溶性糖含量變化一致,均呈遞增變化趨勢。在花器官發(fā)育過程中,9B的可溶性糖含量均高于9A,且在花芽期和花蕾期差異不顯著;而在成花期達到最大值,分別高達11.33、16.15 mg/g,且9B是9A的1.43倍,差異顯著。說明花器官發(fā)育后期,可溶性糖對育性起著關(guān)鍵性作用,與發(fā)生不育的時期有關(guān)。
在花器官整個發(fā)育過程中,不育系9A與保持系9B的游離脯氨酸含量均呈倒U形變化趨勢。隨著花器官的發(fā)育,花蕾期的游離脯氨酸含量高于花芽期和成花期,且達到了顯著性差異;其中,在花芽期9A顯著高于9B,是9B的1.37倍;在花蕾期、成花期,9B則顯著高于9A,分別是9A的1.30、1.40倍,說明不育系中游離脯氨酸含量的顯著降低可能導(dǎo)致雄性不育。
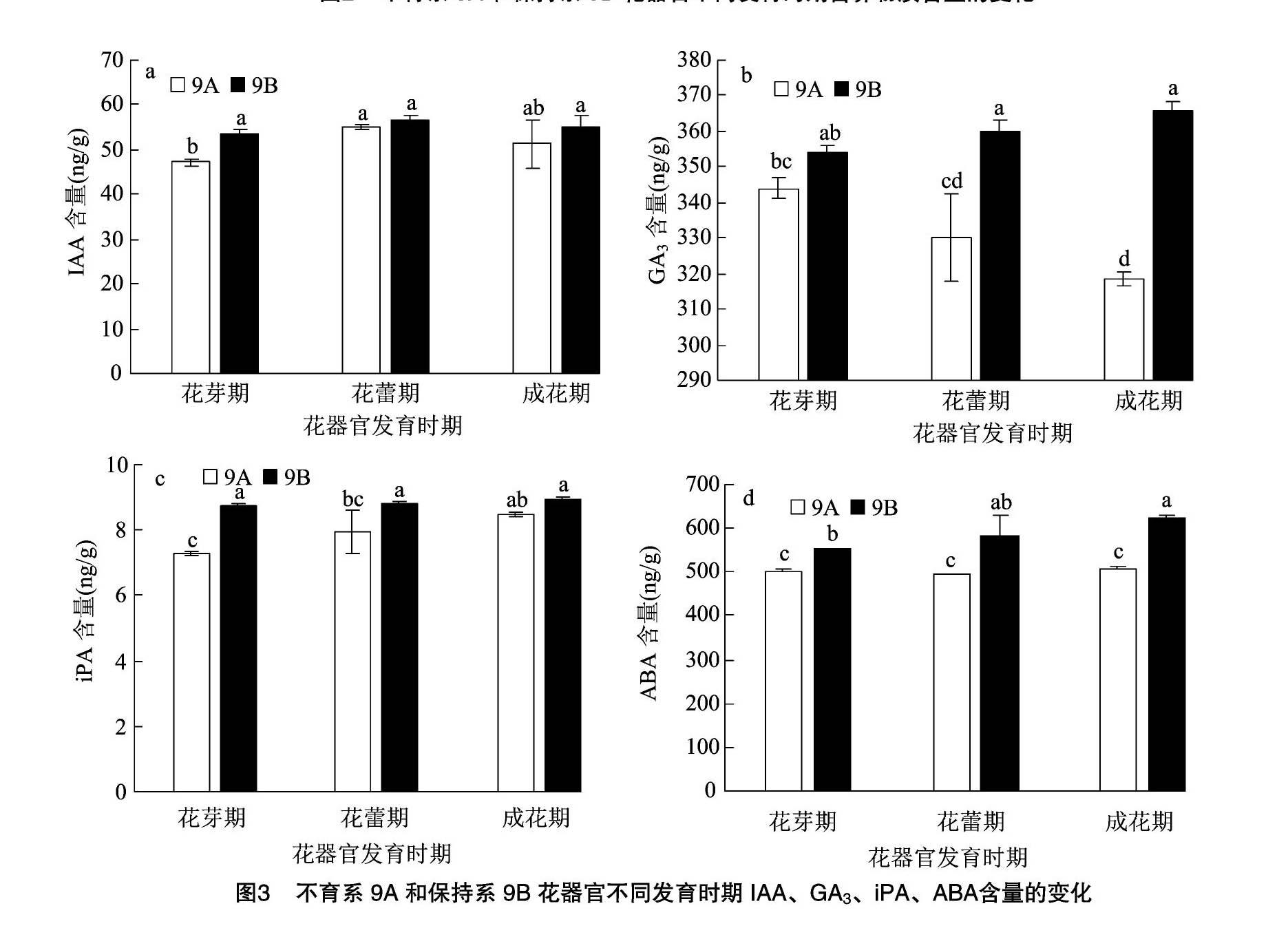
2.3 花器官發(fā)育不同時期內(nèi)源激素含量的變化
如圖3所示,不育系9A和保持系9B的IAA含量變化趨勢一致,均呈倒U形變化,在花蕾期均達到最大值,分別為55.12、56.73 ng/g,但差異不顯著。在花器官發(fā)育過程中,9B的IAA含量均高于9A,且在花芽期9B的IAA含量是9A的1.14倍,差異顯著;而在花蕾期、成花期均未達到顯著性差異。由此推斷,花芽期IAA的缺乏可能導(dǎo)致不育的發(fā)生。
在花器官發(fā)育過程中,不育系9A和保持系9B的GA3含量呈不同的變化趨勢,9A呈逐漸降低趨勢,9B呈逐漸升高趨勢。在花器官發(fā)育過程中,9B的GA3含量均高于9A,且在花蕾期、成花期,9B分別是9A的1.09、1.15倍,達到了顯著性差異。說明GA3的含量與大豆的育性關(guān)系密切,尤其在花器官發(fā)育的后期,這可能是不育發(fā)生的時期。
不育系9A與保持系9B的iPA含量在花器發(fā)育時期均呈逐漸上升趨勢,在成花期均達最大值,但是差異不顯著;在花芽期和花蕾期均達到了顯著性差異,其中,花芽期9B是9A的1.20倍,花蕾期達到1.11倍,說明大豆的雄性不育可能發(fā)生在花芽期或花蕾期。
由花芽期至成花期,不育系9A和保持系9B的ABA含量呈完全不同的變化趨勢,9A呈U形變化趨勢,而9B呈逐漸升高趨勢。在花器官發(fā)育過程中,9B的ABA含量均顯著高于9A,花芽期、花蕾期、成花期9B分別是9A的1.10、1.18、1.23倍,而不育系9A在花器官發(fā)育過程中均沒有顯著性差異。說明ABA含量的積累影響了不育系9A花器官后期的正常發(fā)育,從而導(dǎo)致雄性不育,育性的轉(zhuǎn)變影響植物體內(nèi)ABA含量的積累。
2.4 花器官發(fā)育不同時期內(nèi)源激素之間的平衡關(guān)系
由表1可以看出,隨著花器官的發(fā)育,不育系9A的IAA/ABA的比值先升后降,保持系9B的IAA/ABA比值與9A明顯不同,呈逐漸降低趨勢。9B的IAA/ABA比值除了在花芽期高于9A,其余2個時期均小于9A。
對于IAA/GA3,不育系9A與保持系9B的比值均在花蕾期達最大值,且二者的變化趨勢相同,均為先升后降。花芽期不育系9A的IAA/GA3的值低于保持系9B,花蕾期與成花期不育系9A的 IAA/GA3 的值均高于保持系9B,差距較小,2個時期二者比值均差0.01。
不育系9A與保持系9B在花器官發(fā)育過程中IAA/iPA的比值均呈先升后降趨勢,且二者在花蕾期的比值明顯上升;不育系9A的IAA/iPA的比值在成花期最低,而保持系9B則在花芽期的值最小,不育系9A花芽期與成花期IAA/iPA的值的差距明顯,保持系9B的比值的差距較小,為0.02。花芽期與花蕾期不育系9A的IAA/iPA的值明顯高于保持系9B,在成花期不育系9A的IAA/iPA的值驟降,明顯低于保持系9B。
隨著花器官的發(fā)育,不育系9A、保持系9B的ABA/GA3的值均在成花期達到最大,分別為1.60、1.71,兩者的ABA/GA3的值均隨著花器官的發(fā)育逐漸升高。各時期內(nèi)不育系9A的ABA/GA3的值均低于保持系9B。
3 結(jié)論與討論
3.1 抗氧化酶活性與雄性不育的關(guān)系
研究報道指出,SOD、POD、CAT具有清除植物體內(nèi)毒性氧的作用,可以維持植物體內(nèi)的動態(tài)平衡,使植物能進行正常的生理生化活動和正常發(fā)育[22-24]。蔣會兵等對厚軸茶不育株的生理生化特性進行研究發(fā)現(xiàn),花蕾發(fā)育過程中不育株的POD活性呈上升趨勢,均高于可育株,不育株中的SOD、CAT活性先升后降,顯著低于可育株[25]。王永琦等研究發(fā)現(xiàn),西瓜不育株中SOD、POD活性均高于可育株,而CAT活性則低于可育株[26]。楊龍樹等通過研究大豆不育系的花芽發(fā)現(xiàn),不育系的POD活性顯著上升,且顯著高于同型保持系[27]。張勤等對玉米不育系研究發(fā)現(xiàn),不育株體內(nèi)的SOD、POD活性均顯著高于可育株,CAT活性低于可育株[28]。本試驗結(jié)果顯示,在大豆花器官發(fā)育過程中,不育系的SOD、CAT活性均呈下降趨勢,其同型保持系的SOD、CAT活性均呈先升后降趨勢,不育系的SOD、CAT活性在花芽期均高于其同型保持系,在花蕾期與盛花期均低于保持系;不育系的POD活性在花芽期低于保持系,在花蕾期、成花期顯著高于保持系。該結(jié)果與蔣會兵等對厚軸茶的研究結(jié)果相似,蔣會兵研究發(fā)現(xiàn)在花蕾發(fā)育過程中,厚軸茶不育株的POD活性均低于可育株[25]。本研究結(jié)果表明,在花蕾期和成花期,不育系的POD活性高于保持系,推測可能與不同作物的不同育性有關(guān),具體原因有待進一步研究。
3.2 營養(yǎng)物質(zhì)含量與雄性不育的關(guān)系
蛋白質(zhì)、氨基酸、丙二醛、淀粉與糖類作為植物花器官發(fā)育過程中的重要物質(zhì),對不育系的形成有重要影響,植物花器官中花藥與花粉的異常、敗育均與此類營養(yǎng)物質(zhì)缺乏有關(guān)[29]。本試驗結(jié)果表明,大豆不育系的可溶性蛋白含量低于其同型保持系,與馮文鵬等的研究結(jié)果[30]一致;游離脯氨酸含量在花芽期高于保持系,而在花蕾期、成花期則是低于保持系,推測可能是與作物類型和取樣時期有關(guān)。張勤等研究發(fā)現(xiàn),玉米不育株的可溶性糖含量顯著低于可育株[28],本試驗結(jié)果與其一致。王瑩等研究發(fā)現(xiàn),紫花苜蓿不育株中的淀粉含量在花蕾發(fā)育過程中均低于可育株[31],本試驗結(jié)果與其一致,說明淀粉含量的虧缺能引起花器官發(fā)育異常,從而導(dǎo)致不育。據(jù)此可推測,花器官發(fā)育過程中淀粉含量虧缺可導(dǎo)致大豆花器官發(fā)生敗育,從而導(dǎo)致不育的發(fā)生。MDA作為脂質(zhì)發(fā)生過氧化的終產(chǎn)物,會引起蛋白質(zhì)、核酸等生命大分子發(fā)生聚合反應(yīng),使細胞具有毒性,從而影響花器官的育性[32]。本試驗發(fā)現(xiàn),在花芽期、花蕾期、成花期,不育系中MDA的積累量較多,隨著花器官的發(fā)育先降后升,且均高于保持系;在成花期MDA含量是保持系的1.41倍,說明MDA的含量與花器官的育性密切相關(guān),且高積累量更容易引發(fā)花器官的敗育。
3.3 內(nèi)源激素含量與雄性不育的關(guān)系
生長素、赤霉素與異戊烯基腺嘌呤核苷均具有促進細胞生長與調(diào)控細胞體內(nèi)養(yǎng)分的作用,脫落酸則會抑制細胞生長,以上幾種激素的含量均對植物雄性不育起著重要作用[33]。王永琦等研究發(fā)現(xiàn),西瓜不育株的IAA、GA3、iPA含量均發(fā)生嚴重缺損,在小孢子發(fā)育過程中均顯著低于可育株[26],在白菜[34]、小麥[35]等雄性不育的研究中也得到了相似的結(jié)果。本試驗結(jié)果與王永琦等的研究結(jié)果[26]相似,在大豆花器官的發(fā)育過程中,不育系的IAA、GA3、iPA含量均低于其同型保持系,因此導(dǎo)致9A的敗育。王學(xué)德研究發(fā)現(xiàn),棉花不育株中IAA、GA3含量偏低,而ABA含量偏高[36]。本試驗結(jié)果表明,不育系體內(nèi)的ABA含量在各時期內(nèi)均低于其同型保持系,推測可能與不育發(fā)生的時期不同有關(guān)。
3.4 激素含量比值與雄性不育的關(guān)系
植物體內(nèi)的激素并不是單獨存在的個體,激素之間具有相互促進與拮抗作用,這種激素間的相互作用會導(dǎo)致植物發(fā)生不育[37]。本試驗結(jié)果表明,不育系中IAA/ABA的比值在花器官發(fā)育的花蕾期、成花期均高于其同型保持系;在花芽期不育系IAA/GA3的比值比保持系低,在花蕾期、成花期正好相反;在花芽期、花蕾期不育系IAA/iPA的比值均高于保持系,成花期則相反;隨花器官的發(fā)育,不育系和保持系A(chǔ)BA/GA3的比值均呈遞增變化趨勢,且不育系均低于保持系。不育系及其同型保持系各時期內(nèi)ABA/GA3的比值變化均為逐漸上升趨勢,且各時期內(nèi)不育系的比值均低于其同型保持系。孫希祿等對蘿卜不育株花蕾激素之間的比值進行分析發(fā)現(xiàn),不育株與可育株的IAA/ABA、IAA/GA3、GA3/ABA之間的比值變化不一致[38]。劉紅艷等對芝麻不育株進行研究后發(fā)現(xiàn),其不育株和可育株的IAA/ABA、IAA/SA、IAA/JA比值的變化趨勢差異較大[33]。由此可知,激素比值與作物種類有關(guān),不同的作物其比值的變化趨勢各不相同。
綜上所述,大豆不育系花器官發(fā)育過程中SOD、CAT活性比保持系低,而POD活性隨花器官的形成先升后降;不育系中MDA積累量較保持系高;可溶性糖、可溶性蛋白、淀粉含量均顯著性低于保持系;游離脯氨酸含量先升后降;不育系的IAA、GA3、iPA、ABA含量均顯著低于保持系,且激素比值差異較大,引起激素平衡失調(diào)。因此,不育系9A的敗育與花器官發(fā)育過程中激素含量的缺失有關(guān),據(jù)此推測以上結(jié)果受到大豆質(zhì)核互作雄性不育的影響。花器官發(fā)育至花蕾期后,SOD、CAT活性,MDA、淀粉、可溶性糖、游離脯氨酸、GA3、ABA含量等多項生理生化指標在不育系9A與保持系9B間差距較大,花蕾期可能是大豆質(zhì)核互作雄性不育系生理生化指標發(fā)生異常的關(guān)鍵時期。本研究結(jié)果僅反映了大豆質(zhì)核互作雄性不育系花器官發(fā)育過程中的生理生化變化趨勢,旨在為進一步研究大豆質(zhì)核互作雄性不育的遺傳機制提供理論基礎(chǔ)。
參考文獻:
[1]王曙明,王躍強,李建平,等.田間開放條件下大豆不育系制種技術(shù)研究[J]. 大豆科學(xué),2010,29(3):385-389.
[2]馬小定,邢朝柱.棉花雄性不育研究和應(yīng)用進展[J]. 棉花學(xué)報,2006,18(5):309-314.
[3]陳 秒,尹 超,劉 燁,等.水稻溫敏核不育系育性轉(zhuǎn)換因素及生理生化特性[J]. 作物研究,2013,27(2):164-168.
[4]魯美宏,孫萬倉,孔德晶,等.白菜型冬油菜不育系LRCMS花器生理生化特性及其雄蕊發(fā)育特征研究[J]. 西北植物學(xué)報,2014,34(3):509-515.
[5]劉齊元,劉飛虎,何寬信,等.煙草雄性不育的細胞形態(tài)學(xué)及生理生化研究進展[J]. 亞熱帶植物科學(xué),2004,33(1):69-72.
[6]湯偉華,張蜀寧,孔艷娥,等.不同倍性不結(jié)球白菜Pol CMS及保持系生理生化特性比較[J]. 西北植物學(xué)報,2009,29(1):80-84.
[7]孔艷娥,張紅梅,韓業(yè)飛,等.莖芥菜胞質(zhì)四倍體白菜雄性不育系及保持系生理生化特性研究[J]. 江西農(nóng)業(yè)學(xué)報,2011,23(7):12-15.
[8]王書強,崔輝梅,石國亮.大白菜Ogura雄性不育系及保持系生理生化分析[J]. 新疆農(nóng)業(yè)科學(xué),2011,48(1):110-115.
[9]鄒 佳,藺萬煌,羅紅兵,等.玉米C型胞質(zhì)雄性不育系POD、CAT、SOD活性及POD酶譜分析[J]. 玉米科學(xué),2009,17(6):45-49.
[10]胡小娟.幾種活性物質(zhì)與超氧化物歧化酶相互作用的研究[D]. 合肥:安徽大學(xué),2012.
[11]馮小磊,范光宇,蘇 旭,等.植物雄性不育生理生化研究進展[J]. 作物雜志,2012(3):6-11.
[12]何長征,蕭浪濤,劉志敏,等.植物激素與雄性不育關(guān)系的研究進展[J]. 中國農(nóng)學(xué)通報,2002,18(3):65-69.
[13]余建章,薦 立.沈農(nóng)雄性核不育大豆 L-78-387等位性測驗研究[J]. 沈陽農(nóng)學(xué)院學(xué)報,1985,16(4):19-24.
[14]楊守萍,曾維英,段美萍,等.大豆雄性不育突變體NJ89-1核不育基因的SSR標記和定位[J]. 大豆科學(xué),2006,25(4):344-348.
[15]趙團結(jié),蓋鈞鎰.大豆不育性自然變異的發(fā)現(xiàn)與鑒定[J]. 中國農(nóng)業(yè)科學(xué),2006,39(9):1756-1764.
[16]衛(wèi)保國.大豆光溫敏感型雄性不育系發(fā)現(xiàn)初報[J]. 作物品種資源,1991(3):12.
[17]彭玉華,楊國保,袁建中,等.一個對播種期反應(yīng)敏感的不育大豆特征分析[J]. 作物學(xué)報,1998,24(6):1010-1013.
[18]王 芳,衛(wèi)保國,李貴全,等.大豆光敏雄性不育株88-428BY-827小孢子母細胞的細胞學(xué)觀察[J]. 中國農(nóng)業(yè)科學(xué),2004,37(8):1110-1113,1251.
[19]孫 寰,趙麗梅,黃 梅.大豆質(zhì)-核互作不育系研究[J]. 科學(xué)通報,1993,38(16):1535-1536.
[20]張 磊,戴甌和.大豆質(zhì)核互作不育系W931A的選育研究[J]. 中國農(nóng)業(yè)科學(xué),1997,30(6):90-91.
[21]蓋鈞鎰,丁德榮,崔章林,等.大豆質(zhì)核互作雄性不育系NJCM S1A的選育及其特性[J]. 中國農(nóng)業(yè)科學(xué),1999,32(5):23-27.
[22]張文靜,劉 亮,黃正來,等.低溫脅迫對稻茬小麥根系抗氧化酶活性及內(nèi)源激素含量的影響[J]. 麥類作物學(xué)報,2016,36(4):501-506.
[23]劉淑娟,朱 祺,幸學(xué)俊,等.植物雄性不育影響因素研究進展[J]. 中國農(nóng)學(xué)通報,2014,30(34):46-50.
[24]O’Brien J A,Daudi A,Butt V S,et al.Reactive oxygen species and their role in plant defence and cell wall metabolism[J]. Planta,2012,236(3):765-779.
[25]蔣會兵,許 燕,宋維希,等.厚軸茶雄性不育株花蕾發(fā)育過程中的生理生化變化[J]. 植物生理學(xué)報,2020,56(9):1807-1817.
[26]王永琦,楊小振,莫言玲,等.西瓜雄性不育系‘Se18’抗氧化酶活性和內(nèi)源激素含量變化分析[J]. 園藝學(xué)報,2016,43(11):2161-2172.
[27]楊龍樹,李佳佳,賀亭亭,等.大豆質(zhì)核互作雄性不育系NJCMS1A與其保持系NJCMS1B的生理生化特性比較分析[J]. 大豆科學(xué),2017,36(3):391-398.
[28]張 勤,金圣浩,方 芳,等.玉米光溫敏雄性不育系CB1208-82的生理生化代謝研究[J]. 玉米科學(xué),2019,27(3):48-53.
[29]盛云燕,常 薇,矯士琦,等.甜瓜雄性不育植株雄蕊發(fā)育結(jié)構(gòu)及生理生化特征[J]. 植物生理學(xué)報,2016,52(7):1028-1034.
[30]馮文鵬,周書棟,楊博智,等.辣椒反向溫敏雄性核不育突變體E6421S不育特征及遺傳規(guī)律研究[J]. 園藝學(xué)報,2019,46(6):1112-1122.
[31]王 瑩,王英哲,徐安凱,等.紫花苜蓿細胞質(zhì)雄性不育系及其保持系花蕾的生理生化特性[J]. 中國草地學(xué)報,2018,40(1):24-28,34.
[32]Zheng B B,F(xiàn)ang Y N,Pan Z Y,et al.iTRAQ-based quantitative proteomics analysis revealed alterations of carbohydrate metabolism pathways and mitochondrial proteins in a male sterile cybrid pummelo[J]. Journal of Proteome Research,2014,13(6):2998-3015.
[33]劉紅艷,吳 坤,楊敏敏,等.芝麻顯性細胞核雄性不育系內(nèi)源激素、可溶性糖和淀粉含量變化[J]. 中國油料作物學(xué)報,2014,36(2):175-180.
[34]王永勤.白菜核不育兩用系育性基因表達特征分析及其小孢子發(fā)育相關(guān)基因的分離[D]. 杭州:浙江大學(xué),2003.
[35]張自剛,馬小飛,張紅霞,等.小麥光溫敏雄性不育系BNS育性轉(zhuǎn)換與內(nèi)源激素的關(guān)系研究[J]. 植物遺傳資源學(xué)報,2016,17(5):913-919.
[36]王學(xué)德.棉花細胞質(zhì)雄性不育的研究與利用[J]. 中國農(nóng)業(yè)科學(xué),2019,52(8):1341-1354.
[37]叢 慶,張 琪,宋麗莉,等.激素在植物冷脅迫應(yīng)答中的角色[J]. 核農(nóng)學(xué)報,2016,30(3):614-619.
[38]孫希祿,許小勇,張魯剛.蘿卜雄性不育系花蕾發(fā)育過程中內(nèi)源激素分析[J]. 北方園藝,2011(19):11-15.
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年13期)2021-11-26 11:50:54
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
