萱草糖轉運蛋白HfSWEET2a的克隆及低溫脅迫下的表達分析
2021-01-05 00:35:32黃東梅肖海濤張志國秦巧平
應用技術學報 2020年4期
黃東梅,肖海濤,張志國,白 露,秦巧平
(上海應用技術大學 生態技術與工程學院,上海 201418)
糖類物質不僅作為植物體內能源物質供植物生長發育,還參與植物體內的多種信號轉導途徑[1]。SWEET是一類新發現的具有7個跨膜結構域的糖轉運蛋白,該蛋白可作為糖轉運載體參與糖的運輸[2]。SWEET蛋白有著雙向轉運的功能,可調節葡萄糖跨膜吸收[3],其廣泛存在于原核生物、植物、人類及其他動物中[4]。目前SWEET蛋白在多種作物,如水稻(Oryzasativa)[3]、葡萄(Vitisvinifera)[5]、擬南芥(Arabidopsisthaliana)[6]、高粱(Sorghumbicolor)[7]、大豆(Glycinemax)[8]、番茄(Solanumlycopersicum)[9]等均有相關報道。研究表明,大多數物種SWEET基因家族主要分成4個分支[10],各分支成員之間互相作用共同促進糖的高效運輸和細胞生命活動,且對植物種子萌發、胚胎發育、花蜜產生以及花粉發育也起著十分重要的作用[11-12]。
低溫脅迫是植物生長過程中經常會遭遇的一種災害,它不僅影響著植物的生長和產量,而且容易引起植物細胞發生相應的生理變化,甚至有致使植株死亡的可能性[13]。但是也有部分植物在低溫脅迫下可以通過對自身生理和生化的相應調整,提高對低溫的耐受力,從而達到減輕甚至消除低溫脅迫帶來的傷害[14]。 由于糖的外排影響著糖的分布變化,植物中SWEET基因的調控對生物和非生物脅迫有著多種影響[15]。研究表明,SWEET基因參與調控植物花蜜分泌[16]、花粉發育[17]、赤霉素的反應[12]、衰老[18]、逆境脅迫[19]、種子和果實發育[20]等細胞生命活動。SWEETs以基因家族形式存在于不同植物中,水稻中鑒定了21個成員[3],大豆中52個[8],擬南芥中17個[6]。Xie等[21]通過對荔枝SWEET基因家族的全基因組鑒定和表達分析發現LcSWEET2a/3b參與了早期種子發育。馬鈴薯StSWEET2a在糖脅迫后顯著上調[22],在高溫脅迫下水稻葉片中的OsSWEET2a表達量有著明顯的變化[23],枇杷EjSWEET2a-1在8小時鹽處理后表達量上調顯著[24],以上均表明SWEET2a基因參與了非生物脅迫響應。因此,研究SWEET2a基因應對低溫脅迫的響應有著重要的意義。
萱草是深受大眾喜愛的花卉之一,‘阿斯隆’是萱草中抗寒性較好的品種,有著較高的利用價值。然而,目前尚未有萱草SWEET基因及其表達調控的有關報道。本研究采用同源克隆的方法從萱草‘阿斯隆’葉片中克隆出SWEET2a基因,利用生物信息學分析該基因的序列特征,同時研究其在低溫脅迫處理下的表達情況,旨在為萱草SWEET基因的研究與應用提供一定的參考價值,同時為萱草SWEET基因參與低溫脅迫反應的進一步研究奠定基礎。
1 材料與方法
1.1 試驗材料
萱草‘阿斯隆’(Hemerocallisfulva)植株栽植于上海應用技術大學植物園。將萱草栽植于富含腐殖質的濕潤土壤中,置于陽光充足、通風良好的地方培養。5~7天左右澆水一次,正常施肥,根據具體情況進行相應的除草、換盆換土等養護管理。采集萱草‘阿斯隆’長勢良好且一致的葉片,用蒸餾水洗凈晾干,液氮速凍并于-80 ℃下儲存用于后續試驗。
1.2 RNA的提取及cDNA的合成
使用改良的CTAB法[25]提取萱草葉片RNA,采用超微量分光光度法和凝膠電泳法檢測RNA的濃度、純度和完整性,-80 ℃保存備用。檢測合格的RNA采用M-MuLV 第一鏈 cDNA 合成試劑盒進行第一鏈cDNA的合成(上海生工),-20 ℃保存備用。
1.3 基因克隆
以萱草‘阿斯隆’葉片的cDNA為模板,根據轉錄組測序結果篩選到的SWEET2a基因序列在兩端設計引物(見表1),采用PCR技術擴增目的片段并克隆到pUCm-T Vector,進行測序驗證得到全長cDNA序列。實驗所需引物序列見表1,引物及測序委托生工生物工程(上海)有限公司完成。
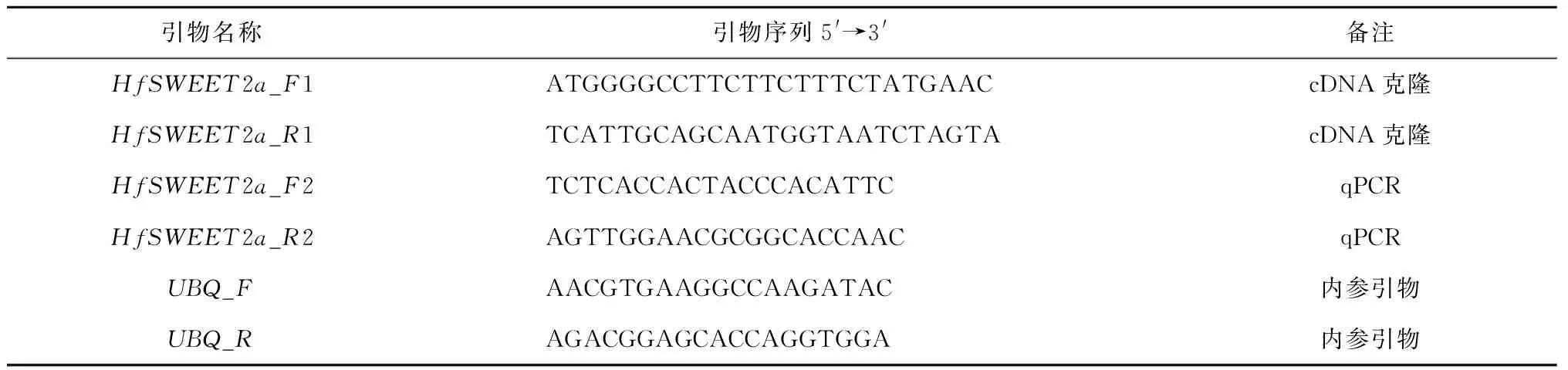
表1 實驗所需引物序列Tab.1 Sequence of primers used in this experiment
1.4 基因的生物信息學分析
利用ExPASY(https://web.expasy.org/translate/)網站對SWEET2a核苷酸序列進行翻譯,用ProtParam(http://web.expasy.org/protparam)網站進行蛋白質理化性質分析,用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)網站進行亞細胞定位預測,蛋白質跨膜結構分析采用TMHMM在線工具(http://www.cbs.dtu.dk/services/TMHMM/)進行。以HfSWEET2a蛋白序列在NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)網站搜索同源蛋白序列,下載ONK69846.1,蘆筍asparagus;XP_008775459.1,海棗date;XP_020701878.1,鐵皮石斛dendrobe;THU64444.1,野蕉banana;XP_026394894.1,罌粟opium;OVA00198.1,博落回macleaya;XP_004968893.1,小米millet;XP_021313692.1高粱sorghum;NP_001146103.1,玉米maize;OAY69557.1,菠蘿pineapple;XP_020595974.1,蝴蝶蘭orchid;MQL94069.1,芋頭taro;RWR97356.1,沉水樟cinnamomun;XP_031490470.1,藍星睡蓮lotus;XP_012848541.1,猴面花guttata;XP_024020545.1,川桑mulberry;XP_011623118.1,無油樟amborella;TXG47757.1,漾濞槭acer;XP_017234459.1,黃胡蘿卜carrot;XP_006467563.1,橙orange;XP_022715681.1,榴梿durian;QHT64202.1,荔枝Litchi;KAF3666069.1,辣椒pepper;AMQ35580.1,馬鈴薯potato;QIE48602.1,茉莉花jasmine;XP_028118815.1,茶camellia;XP_007025233.2,可可樹cacao;XP_020553914.1,胡麻flax等28種SWEET2a蛋白序列進行后續分析。利用Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)和jalview2.10.2軟件進行多序列比對,再用MEGA7.0軟件輸出基于neighhor-joining法構建的系統進化樹,bootstrap值設置為500次重復。用MEME(http://meme-suite.org/tools/meme)進行motif比對,再用NCBI的CD-Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)進行保守結構域查詢。
1.5 低溫處理及基因定量表達分析
為進一步了解基因對低溫脅迫的響應,選長勢一致的萱草幼苗移入低溫人工氣候箱中,分別置于15 ℃、10 ℃、5 ℃、0 ℃下處理,24 h后進行取樣,取樣部位為完全展開的功能葉片,設置3次生物學重復。利用Primer5(https://sg.idtdna.com/pages/tools/primerquest)網站,根據HfSWEET2a基因cDNA序列設計定量表達分析所用引物HfSWEET2a_F2/HfSWEET2a_R2,內參為UBQ(見表1)。提取葉片RNA、反轉錄獲得cDNA后,進行qPCR,反應條件為95 ℃預變性5 min;95 ℃變性10 s,60 ℃退火30 s,循環40次;融解曲線采集程序為95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。采用2-ΔΔCt法計算基因相對表達量,同時計算其標準差,再用Excel和SPSS22.0對數據進行處理。
2 結果與分析
2.1 萱草SWEET2a基因的克隆及序列分析
以萱草‘阿斯隆’葉片的cDNA為模板,通過PCR技術克隆得到長度為 1 045 bp的核苷酸序列,命名為HfSWEET2a,該基因已經提交GenBank,登錄號為MT379659。序列分析顯示,HfSWEET2a含有690 bp的完整開放式閱讀框,編碼229個氨基酸(圖1)。蛋白質理化性質分析表明,HfSWEET2a編碼的蛋白分子量25.531 kDa,等電點9.16,定位在細胞膜。蛋白質跨膜結構分析顯示,HfSWEET2a含有典型的7個跨膜結構域,其位置分別為:10-32,45-67,71-93,105-127,137-159,166-188,193-215(見圖2)。

下劃線表示PCR引物序列,黑色標出字母分別表示起始密碼子和終止密碼子灰色部分代表兩個保守結構域,框出部分代表7個跨膜結構域(TM1-7)圖1 萱草SWEET2a基因的核苷酸序列及編碼氨基酸序列Fig.1 Nucleotide sequence and encoding amino acid sequence of HfSWEET2a
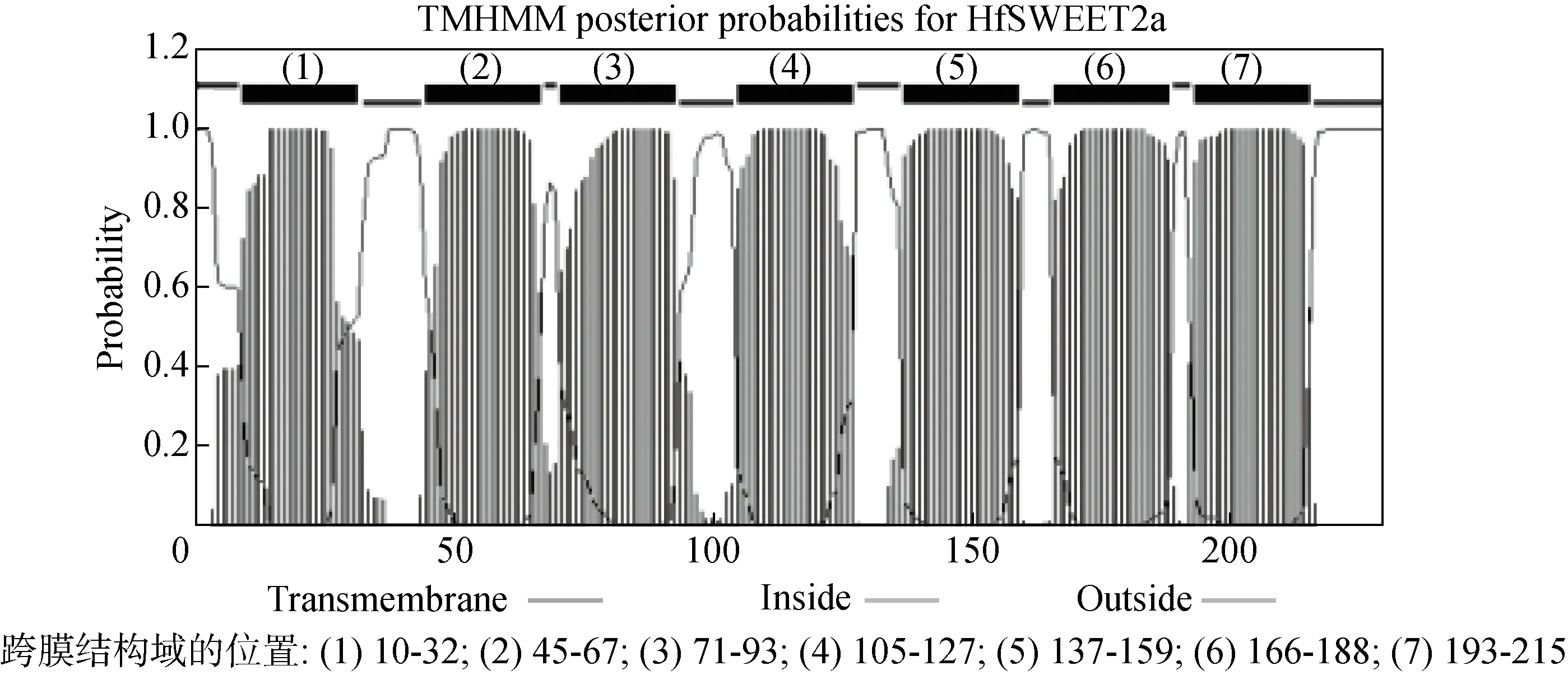
圖2 HfSWEET2a蛋白的跨膜結構預測Fig.2 Transmembrane probabilities for HfSWEET2a
2.2 萱草HfSWEET2a基因編碼蛋白與其他植物進化關系分析
BLASTX比對結果表明,HfSWEET2a的蛋白序列與蘆筍、菠蘿、野蕉等SWEET2a蛋白序列相似性較高,分別為75.58%、74.6%、73.09%。將HfSWEET2a蛋白與蘆筍ONK69846.1等10條蛋白質序列進行序列比對,結果顯示HfSWEET2a與這些蛋白的保守結構域高度相似。motif比對結果顯示,它們都有3個相同的motif,且均為PQ-loop家族(見圖3中的PQ-loop 1、PQ-loop 2和PQ-loop 3)。
系統進化分析結果顯示單子葉和雙子葉SWEET2a蛋白分別聚為兩類,HfSWEET2a與蘆筍的系統進化關系最近,聚在同一分支,其次為菠蘿、野蕉和海棗,與雙子葉植物荔枝、胡麻以及茉莉花等的親緣關系較遠(見圖4)。

圖3 萱草HfSWEET2a編碼蛋白與其他10種植物SWEET2a蛋白的motif比對Fig.3 The motif comparison of HfSWEET2a and SWEET2a proteins from other 10 plants
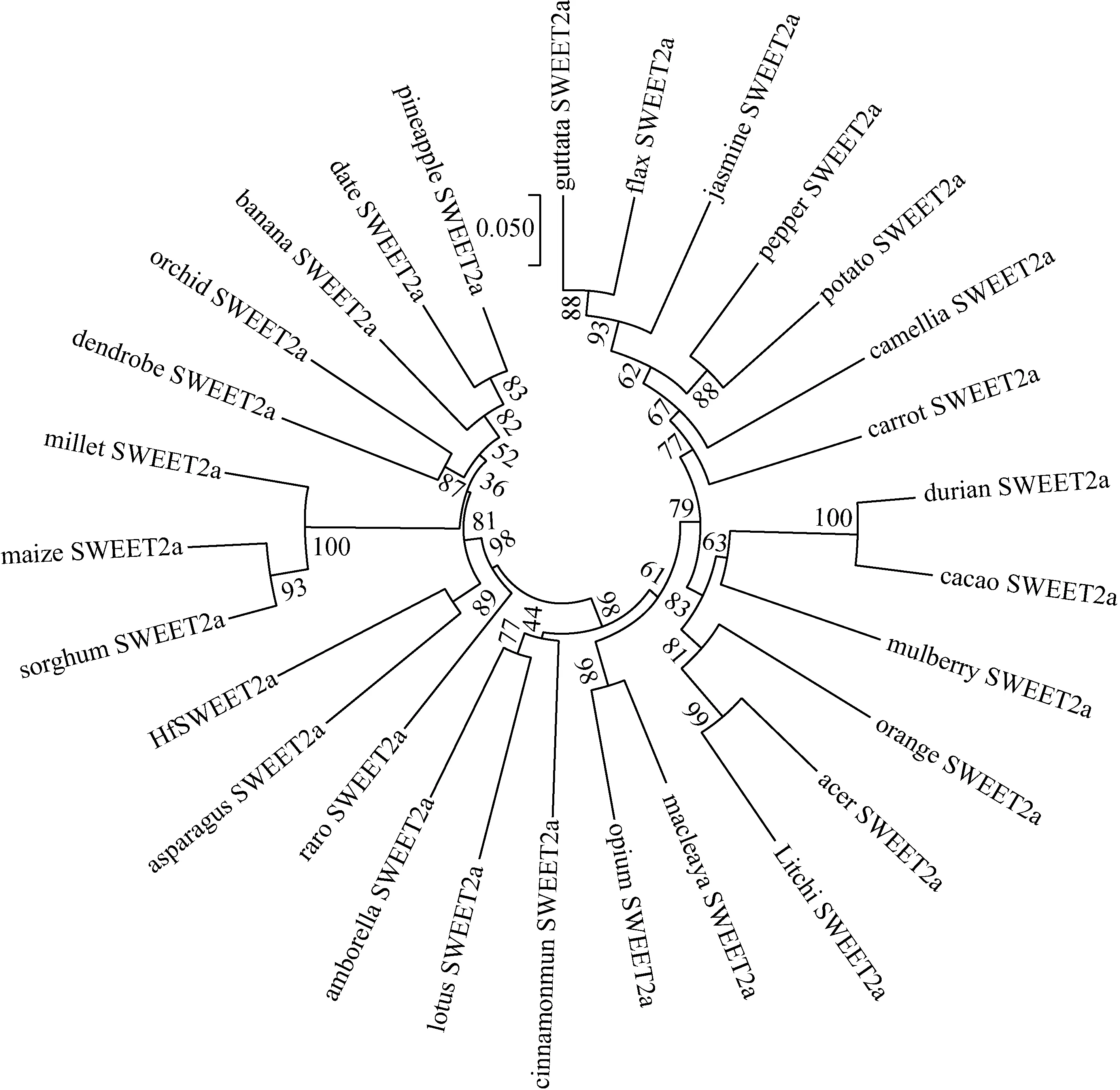
圖4 萱草HfSWEET2a與其他28種植物SWEET2a蛋白的系統進化分析Fig.4 The phylogenetic analysis HfSWEET2a and SWEET2a proteins from other 28 plants
2.3 低溫對萱草HfSWEET2a基因表達的影響
基因定量表達分析顯示,在不同溫度處理下,HfSWEET2a的表達呈現出不同程度的變化。HfSWEET2a基因在0 ℃下表達量最高,0 ℃相對表達量是15 ℃下的3.3倍、10 ℃下的1.7倍、5 ℃下的3.7倍(見圖5)。統計分析顯示,5 ℃和15 ℃下的HfSWEET2a基因表達差異不顯著,其他溫度下的基因表達均達到顯著差異水平(P<0.05)。

柱形表示平均值±標準誤,相同字母表示差異不顯著圖5 不同溫度處理后HfSWEET2a基因的表達變化Fig.5 The expression of HfSWEET2a under different temperature
3 討論
糖是植物光合作用的主要產物,也是植物體內的主要能源物質[1]。SWEET蛋白廣泛存在于各種植物中,參與植物生長發育和逆境響應。目前已有大豆[8]、擬南芥[6]、小麥(Triticumaestivum)[26]、陸地棉(Gossypiumhirsutum)[27]等植物上均有關于SWEET蛋白的研究報道,但萱草作為我國傳統花卉之一,目前未見有關SWEET基因的相關報道。本研究通過PCR技術從萱草‘阿斯隆’葉片中克隆出HfSWEET2a的全長cDNA序列,同源序列比對結果表明,HfSWEET2a基因所編碼的蛋白質序列與蘆筍、菠蘿以及海棗等植物SWEET蛋白質序列同源性較高,具有高度相似的保守結構域,且具有均為PQ-loop家族的3個相同的motif。
SWEET家族基因一般都具有典型的6~7個跨膜結構域,菠蘿中39個SWEET蛋白中大部分包含7個ɑ-螺旋的跨膜結構域[10],水稻中有18條OsSWEET蛋白具有6-7個跨膜結構域[28],雷蒙德氏棉SWEET基因家族跨膜結構域預測結果顯示有22個SWEET蛋白含有7個跨膜結構域,7個SWEET蛋白含有6個跨膜結構域[29]。這些跨膜結構域使其具有保守的跨膜糖轉運功能以及介導植物對疾病的感病功能[30],本研究分離到的HfSWEET2a與其他植物SWEET蛋白相似,具有7個保守的跨膜結構域,故HfSWEET2a基因屬于SWEET基因家族,可能參與糖的跨膜運輸。
低溫逆境脅迫是植物中常見的一種非生物脅迫,在低溫逆境脅迫下,會有很多結構及生理生化方面的變化,其中可溶性糖含量變化是重要指標之一[31]。可溶性糖是植物在低溫逆境下的呼吸作用底物,對保護物質起滲透作用,可增強植物的耐寒性[32-33]。當遭遇低溫脅迫環境時,為減少逆境脅迫帶來的傷害,植物體內糖類含量會有明顯的增長[34-35]。SWEET蛋白作為糖轉運載體參與糖的運輸、分配和貯藏,被運輸到植物體內不同組織、細胞器中,參與植物生長發育的重要生理過程[36],不僅可以響應生物脅迫,而且也是非生物脅迫響應的重要蛋白,SWEETs可以通過調控糖的運輸和分配來參與非生物脅迫的響應[37]。
許多植物基因低溫響應的順式元件LTR位于SWEET基因的啟動子區域[38],SWEET蛋白的中斷對擬南芥的耐寒性有較大的負面影響,耐寒力遠不如正常植株[39],而AtSWEET16過量表達植物卻比平常表現出更好的抗凍性[40]。CsSWEET16也被證明在改善擬南芥的耐寒性方面了發揮了重要作用[19]。在本研究中,萱草葉片在不同低溫情況處理,HfSWEET2a基因表達發生了不同程度的變化,這與茶樹、番茄、甘藍等研究中SWEET基因的表達較為一致。茶樹在冷訓化過程中,不同SWEET基因的表達量有著不同的變化,CsSWEET2、CsSWEET3和CsSWEET16的表達受到了廣泛的抑制,而CsSWEET17和CsSWEET1的表達量卻有著顯著地提高[41]。番茄中9種SWEET基因在低溫處理下葉片中的表達量上調了好幾倍,而在根中則顯著下調[42]。甘藍中BoSWEET16a和BoSWEET17的表達在低溫脅迫后迅速下降,并在12 h至48 h保持低水平狀態[43]。本研究初步表明,HfSWEET2a基因可能參與了低溫脅迫應答反應并發揮著重要作用,具體作用機制有待進一步研究。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
