微生物甲硫氨酸合成調(diào)控的綜合研究進(jìn)展與展望
2020-12-31 09:07:40趙嫚彭莉成浩應(yīng)向賢汪釗
食品與發(fā)酵工業(yè) 2020年24期
趙嫚,彭莉,成浩,應(yīng)向賢,汪釗
(浙江工業(yè)大學(xué) 生物工程學(xué)院,浙江 杭州, 310014)
甲硫氨酸(methionine,Met),包括L-甲硫氨酸和D-甲硫氨酸兩種構(gòu)型,L-甲硫氨酸是人和動(dòng)物必需的含硫氨基酸,在生物體內(nèi)具有重要的生理生化功能。具體功能包括參與DNA、蛋白質(zhì)的合成和蛋白結(jié)構(gòu)的穩(wěn)定;是精胺、亞精胺和乙烯等的前體,參與細(xì)胞分裂分化、凋亡、穩(wěn)態(tài)和基因表達(dá)等生物生長(zhǎng)發(fā)育的各個(gè)方面;并通過(guò)其主要代謝產(chǎn)物S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)間接調(diào)節(jié)各種代謝過(guò)程,為脂類(lèi)、蛋白質(zhì)、核酸、生物堿類(lèi)和植物固醇等多種化合物提供甲基[1]。近年來(lái),Met的需求量大幅度增加,預(yù)計(jì)2022年全球市場(chǎng)份額可達(dá)73億美元,其中飼料添加劑是最大的消費(fèi)市場(chǎng)。此外,Met在食品添加劑、醫(yī)藥和化妝品等領(lǐng)域的需求也呈長(zhǎng)期穩(wěn)定增長(zhǎng)趨勢(shì)[2-5]。
Met的合成主要包括化學(xué)法、酶法和微生物發(fā)酵法3種,目前工業(yè)上主要采用化學(xué)法和化學(xué)-酶法。盡管化學(xué)法和酶法技術(shù)成熟、成本低,但微生物發(fā)酵法因其綠色、高效和周期短等優(yōu)勢(shì),受到研究者的廣泛關(guān)注。Met發(fā)酵法合成的研究主要集中在大腸桿菌(Escherichiacoli)和谷氨酸棒狀桿菌(Corynebacteriumglutamicum)中,目前已經(jīng)報(bào)道的E.coli和C.glutamicum中Met的最高產(chǎn)量分別約為30 g/L和9.88 g/L[6-15],如表1所示,該水平距離工業(yè)化應(yīng)用還存在一定的差距。隨著Met市場(chǎng)需求的逐年增加,對(duì)提高M(jìn)et產(chǎn)量的研究迫在眉睫。代謝物產(chǎn)量的高低與代謝合成過(guò)程中關(guān)鍵基因的相互調(diào)控密不可分。本文將對(duì)近幾年Met的合成過(guò)程、調(diào)控機(jī)制和環(huán)境因素的研究進(jìn)展進(jìn)行全面綜述,以期為Met合成產(chǎn)量的提高提供基礎(chǔ)。
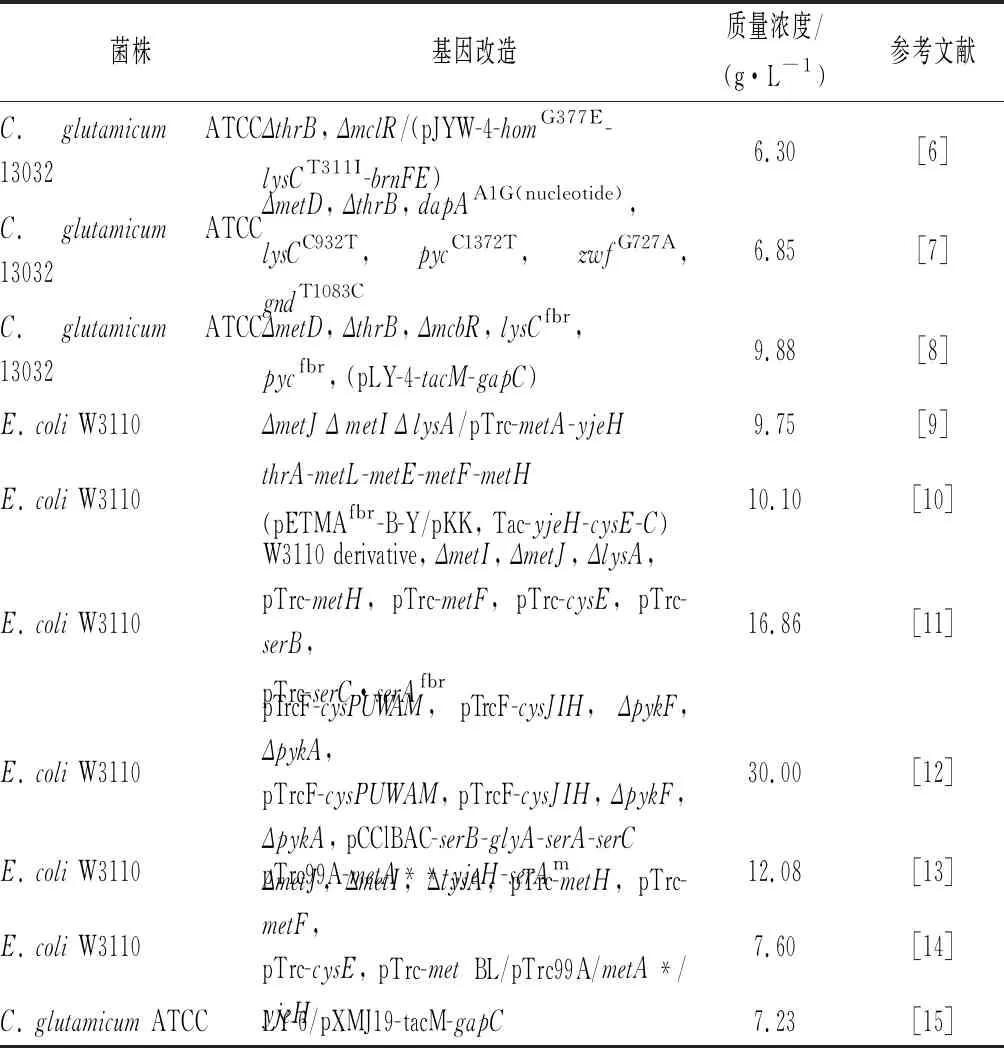
表1 甲硫氨酸產(chǎn)生菌構(gòu)建的典型實(shí)例Table 1 Representative examples of the methionine producing strains
1 Met的生物合成
Met能夠在大多數(shù)的植物和微生物中合成,其合成途徑已經(jīng)被完全解析,主要涉及兩個(gè)部分,即從頭合成途徑-天冬氨酸合成途徑和硫同化過(guò)程(圖1)[16-17]。
1.1 從頭合成途徑
除Met以外,必需氨基酸異亮氨酸、賴氨酸和蘇氨酸也是由該途徑的不同分支產(chǎn)生(圖1)。以C.glutamicum中從頭合成途徑為例,Met合成的具體過(guò)程是天冬氨酸在天冬氨酸激酶(Aspartate kinase, lysC)的催化下形成天冬氨酰-4-磷酸,然后被天冬氨酸半醛脫氫酶(Aspartaldehyde dehydrogenase, asd)催化形成天冬氨酸-4-半醛,后者被高絲氨酸脫氫酶(Homoserine dehydrogenase, hom)氧化成高絲氨酸;高絲氨酸在高絲氨酸-O-乙酰基轉(zhuǎn)移酶(Homoserine acetyltransderase, metX)的作用下生成O-乙酰-L-高絲氨酸,然后再依次在胱硫醚γ合成酶(Cystathinine γ-synthase, metB)和胱硫醚β裂解酶的作用下分別形成胱硫醚和高半胱氨酸,最后高半胱氨酸在甲硫氨酸合成酶(Methionine synthase, metEH)的作用下,以四氫葉酸(CH3-THF)作為甲基供體合成Met。此外,在C.glutamicum中O-乙酰-L-高絲氨酸還可以在O-乙酰-L-高絲氨酸硫化酶(O-acetylhomoserine sulfhydrylase, metY)的作用下直接合成高半胱氨酸,進(jìn)而合成Met。
在不同的生物中,合成途徑存在一定的差異(圖1, 表2),如高絲氨酸在C.glutamicum中經(jīng)過(guò)高絲氨酸乙酰轉(zhuǎn)移酶(Homoserine acetyltransferase, HAT)催化形成O-乙酰-L-高絲氨酸,而在E.coli和植物中則分別被高絲氨酸-O-琥珀酰基轉(zhuǎn)移酶(Homoserine succinyltransderase, metA)和高絲氨酸激酶(Homoserine kinase, HK)催化形成O-琥珀酰-L-高絲氨酸和O-磷酸-L-高絲氨酸[18]。在古細(xì)菌中,天冬氨酸-4-半醛可以在硫化酶的作用下,直接合成高半胱氨酸[17]。
1.2 硫同化


圖1 甲硫氨酸生物合成途徑與調(diào)控Fig.1 Biosynthesis pathway and regulation of Methionine
從頭合成途徑中天冬氨酸直接合成Met需要1個(gè)ATP和2個(gè)NADPH,無(wú)機(jī)硫的同化則需要2個(gè)ATP和4個(gè)NADPH。因此Met的合成除了受到從頭合成途徑和硫同化內(nèi)部途徑關(guān)鍵酶等合成過(guò)程的調(diào)控外,還受到能量、輔因子和外界環(huán)境等多種不同層次的綜合調(diào)控。
2 Met合成過(guò)程的調(diào)控
2.1 Met合成關(guān)鍵酶的調(diào)控
甲硫氨酸的合成受到多個(gè)關(guān)鍵酶的調(diào)控。首先,天冬氨酸激酶(Aspartate kinase,AK)是Met合成途徑的第一個(gè)限速酶,催化ATP依賴的天冬氨酸磷酸化形成天冬氨酰-4-磷酸。在E.coli中AK由thrA、metL和lysC三個(gè)基因編碼,這些基因的表達(dá)分別受蘇氨酸、Met和賴氨酸的反饋?zhàn)瓒粽{(diào)節(jié)[21]。而在C.glutamicum中AK僅有一個(gè)編碼基因lysC,其表達(dá)同時(shí)受到蘇氨酸和賴氨酸的反饋?zhàn)瓒鬧22]。過(guò)表達(dá)lysC以及不受賴氨酸反饋抑制的突變體可使最終產(chǎn)物的含量顯著提高。此外,在酶活水平上,AK還受其終端產(chǎn)物反饋抑制調(diào)節(jié)。在C.glutamicum中,對(duì)基因lysC進(jìn)行定點(diǎn)突變?yōu)锳279T和G359D,結(jié)果表明,lysC的突變可部分解除AK的活性受蘇氨酸和賴氨酸的反饋?zhàn)瓒粢种芠21,23]。高絲氨酸脫氫酶是天冬氨酸合成途徑的關(guān)鍵雙功能酶,包含天冬氨酸激酶和高絲氨酸脫氫酶兩個(gè)功能。在E.coli中,高絲氨酸脫氫酶是由thrA和metL兩個(gè)基因進(jìn)行編碼。thrA的轉(zhuǎn)錄受到蘇氨酸和異亮氨酸的抑制,具體原因是thrA、thrB(在蘇氨酸合成途徑中編碼高絲氨酸激酶)和thrC(編碼蘇氨酸合成酶)在染色體上組成thrABC操縱子,蘇氨酸和異亮氨酸通過(guò)與該操縱子上游的一段前導(dǎo)序列作用,反饋抑制基因的表達(dá);而metL僅受到Met的反饋抑制。而在C.glutamicum中hom受到蘇氨酸的抑制,且其G378S定點(diǎn)突變可部分解除hom的活性受蘇氨酸和賴氨酸的反饋?zhàn)瓒粢种芠24-25]。在Met的合成過(guò)程中,除了Met以外,其下游產(chǎn)物S-腺苷甲硫氨酸(S- adenosylmethionine, SAM)也參與到Met的合成調(diào)控,比如metA和metB均受到SAM的反饋抑制,進(jìn)而影響Met的合成。

表2 L-甲硫氨酸生物合成相關(guān)的酶及其編碼基因Table 2 Enzymes and their encoding genes related to the methionine synthesis
在E.coli中,metA、metB和metC位于一個(gè)操縱子上,高效表達(dá)該操縱子基因是提高M(jìn)et常用的策略。USUDA等[26]在E.coliW3110 中過(guò)表達(dá)metA,使Met的含量從0.13 g/L提高到0.24 g/L。郭謙等[27]在一株經(jīng)過(guò)紫外誘變篩選選育出的Met高產(chǎn)菌株E.coliYB12的基礎(chǔ)上,過(guò)表達(dá)metA、cysE(絲氨酸合成半胱氨酸途徑中絲氨酸乙酰轉(zhuǎn)移酶編碼基因)和編碼Met轉(zhuǎn)運(yùn)蛋白的yeaS基因,使E.coliYB12中Met的含量從60 mg/L提高到251 mg/L。另外,在1株生產(chǎn)賴氨酸的C.glutamicum菌株中,對(duì)metX和metY基因過(guò)表達(dá)導(dǎo)致Met的產(chǎn)量提高到16 g/L[28]。因此,Met合成途徑的這些關(guān)鍵酶是調(diào)控其代謝途徑的關(guān)鍵限速酶。WEI等[29]在提高M(jìn)et重要中間體O-乙酰高絲氨酸的研究中,基于蛋白質(zhì)工程的進(jìn)化保守分析和結(jié)構(gòu)指導(dǎo)的工程化提高了關(guān)鍵酶高絲氨酸乙酰轉(zhuǎn)移酶(Homoserine acetyltransferase, metXlm)的活性且減少反饋抑制,使酶的活性增加了12.5倍,并通過(guò)守恒優(yōu)化,最終使產(chǎn)物O-乙酰高絲氨酸產(chǎn)量增加57.14%[29]。
2.2 Met合成競(jìng)爭(zhēng)途徑的阻斷
天冬氨酸合成途徑中的賴氨酸和蘇氨酸合成途徑是Met合成的2個(gè)關(guān)鍵競(jìng)爭(zhēng)途徑,分別通過(guò)天冬氨酸-4-半醛和高絲氨酸實(shí)現(xiàn)競(jìng)爭(zhēng)。研究發(fā)現(xiàn),2個(gè)競(jìng)爭(zhēng)途徑的去除會(huì)為Met的合成提供更多的天冬氨酸-4-半醛和高絲氨酸底物,從而有效提高M(jìn)et的產(chǎn)量。研究中常用的是賴氨酸合成途徑中dapA(在賴氨酸合成途徑編碼二氫二吡啶甲酸合酶)和蘇氨酸合成途徑中thrB的敲除[16]。例如,在C.glutamicum13032中通過(guò)thrB的敲除和賴氨酸合成途徑的減弱(dapA中用稀有密碼子GTG代替起始密碼子ATG),Met的產(chǎn)量從0.89 g/L提高到2.99 g/L,增加了3倍[7]。在E.coliW3110中,通過(guò)thrBC的敲除獲得蘇氨酸營(yíng)養(yǎng)不良菌株,與野生型相比,其Met產(chǎn)量從無(wú)法檢測(cè)提高到0.008 g/L[26]。吳婷婷[30]在E.coliK-12/pKD46菌株中敲除dapA基因后,其Met的含量提高了2.61倍。
2.3 Met轉(zhuǎn)錄調(diào)節(jié)子的轉(zhuǎn)錄調(diào)控
Met合成的轉(zhuǎn)錄調(diào)控除了限速酶的表達(dá)受到產(chǎn)物的反饋抑制外,還包括各種調(diào)控蛋白的調(diào)控[31],如MetJ和MetR。MetJ是Met合成的關(guān)鍵阻遏蛋白,在E.coli中阻遏metL、metA、metB、metC和metE基因的表達(dá)[32]。研究發(fā)現(xiàn),將MetJ第54位的Ser替換為Asn,可以消除其反饋?zhàn)瓒糇饔茫岣進(jìn)et的產(chǎn)量[33]。與E.coli相對(duì)應(yīng)的,在C.glutamicum中Met的合成和攝入則是由McbR阻遏蛋白調(diào)控。McbR是C.glutamicum硫代謝的全局調(diào)控因子,涉及的基因包括hom、metX、metB、metE和metH等[34]。很多研究已經(jīng)表明,通過(guò)對(duì)細(xì)菌metJ或mcbR基因的敲除或修飾,可解除其對(duì)細(xì)胞生產(chǎn)Met能力的禁錮,使菌株產(chǎn)生更多的Met[6,9,26-27,35]。與MetJ相反,MetR是一類(lèi)轉(zhuǎn)錄激活因子,過(guò)表達(dá)后可大大促進(jìn)Met的合成,前期研究,通過(guò)共表達(dá)metE和metR的研究發(fā)現(xiàn),MetR是metE基因表達(dá)的反式激活因子,然而在MetJ蛋白和SAM的存在下,metR基因的表達(dá)被抑制[36]。
2.4 Met轉(zhuǎn)運(yùn)體的調(diào)控
菌株經(jīng)過(guò)一系列代謝工程改造后,胞內(nèi)氨基酸的積累會(huì)抑制其合成途徑關(guān)鍵酶的催化活性,增加其降解途徑的前體可利用性,甚至抑制細(xì)胞生長(zhǎng)。因此,Met的轉(zhuǎn)運(yùn)也成為其生產(chǎn)的限速步驟[31,37]。在E.coli中,過(guò)表達(dá)Met輸出蛋白YjeH、YeaS和敲除Met攝入蛋白MetD、MetP均顯著增加了Met的積累[38-39]。在基因工程菌C.glutamicum中敲除Met的攝取蛋白MetQP或過(guò)表達(dá)輸出蛋白BrnFE或MetT,Met的含量都顯著提高[40-41]。
3 硫同化對(duì)Met合成的調(diào)控
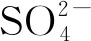
4 NADPH對(duì)Met合成的影響
Met的合成從硫同化到從頭合成途徑共需要6個(gè)NADPH和3個(gè)ATP。因此能量和還原力的提高是保證Met高效合成的關(guān)鍵。NADPH在C.glutamicum和E.coli中分別是以不同的方式進(jìn)行提供。在C.glutamicum中,NADPH主要是通過(guò)戊糖磷酸途徑中的葡萄糖-6-磷酸脫氫酶(G6PDH, zwf)和6-磷酸葡萄糖酸脫氫酶(6PGDH, gnd),而在E.coli中主要是通過(guò)膜結(jié)合的轉(zhuǎn)氫酶合成[48-49]。此外,前人的研究已經(jīng)嘗試了不同策略來(lái)增加NADPH的供應(yīng),最常用的是調(diào)整糖酵解途徑和戊糖磷酸途徑之間的代謝流量比。C.glutamicum戊糖磷酸途徑中的zwf和gnd進(jìn)行突變以消除其反饋抑制,構(gòu)建了zwfA243T和gndS361F突變體后,氨基酸的產(chǎn)量呈現(xiàn)了不同程度的增加[50]。此外,通過(guò)過(guò)表達(dá)果糖-1,6-二磷酸酶基因(fbp),加速碳源從糖酵解轉(zhuǎn)向戊糖磷酸途徑,也顯著增加了還原力的提供[49,51]。此外,BOMMAREDDY等[52]不依賴于戊糖磷酸途徑的想法,僅通過(guò)改變天然NAD+依賴的甘油醛-3-磷酸脫氫酶(Glyceraldehyde-3-phosphate dehydrogenase, gapA)的輔酶專一性來(lái)生成NADP+,在提高甘油醛-3-磷酸脫氫酶對(duì)NADP+的催化效率的同時(shí),所有受試突變體的氨基酸產(chǎn)量都得到了顯著提高(60%)。因此,糖酵解、戊糖磷酸途徑和三羧酸循環(huán)等初級(jí)代謝途徑中能量和輔助因子的通量變化通常會(huì)直接影響氨基酸的生物合成,因此,它們?yōu)樯锕こ剔D(zhuǎn)化提供了一個(gè)潛在的目標(biāo),包括與能量和還原力形成相關(guān)的反應(yīng)。
5 環(huán)境營(yíng)養(yǎng)元素對(duì)Met合成的影響
5.1 營(yíng)養(yǎng)元素對(duì)Met合成的影響
碳源是微生物生長(zhǎng)的物質(zhì)基礎(chǔ)。在工業(yè)氨基酸發(fā)酵中,葡萄糖、乳糖、可溶性淀粉、甘油和蔗糖是目前最常使用的碳源,其濃度過(guò)高或過(guò)低均會(huì)影響Met的合成。比如,劉俊琦等[53]向培養(yǎng)基中添加高濃度的葡萄糖,導(dǎo)致菌體濃度和Met產(chǎn)量都有所下降。進(jìn)一步的研究發(fā)現(xiàn),不同的菌株對(duì)不同碳源的利用具有顯著的順序作用,如酵母菌、E.coli和枯草芽孢桿菌(Bacillussubtilis)順序利用較為明顯,而C.glutamcum很少表現(xiàn)出對(duì)底物碳源的共利用[54]。可見(jiàn)碳源的種類(lèi)和比例對(duì)Met的產(chǎn)量有明顯的影響,因此可通過(guò)合適的碳源配比來(lái)提高M(jìn)et生產(chǎn)水平。
氮源也是微生物生長(zhǎng)的必需營(yíng)養(yǎng)要素,為生物體合成蛋白質(zhì)、核酸及其他含氮化合物提供原料。牛肉膏、蛋白胨及尿素是微生物常利用的氮源。不同的氮源對(duì)Met產(chǎn)量和菌體濃度均有影響,和其他氮素相比較,尿素的效果最好,適宜的尿素濃度對(duì)Met的產(chǎn)量至關(guān)重要[53]。
無(wú)機(jī)鹽是維持細(xì)胞穩(wěn)態(tài)的重要因子,在Met的合成中不僅提供必需的環(huán)境和營(yíng)養(yǎng)成分,而且還可以合成Met螯合物。Met螯合物因其具有較高的生物價(jià)值,增加牲畜的免疫力,現(xiàn)被廣泛用于飼料生產(chǎn)[55],如甲硫氨酸硒[56]、甲硫氨酸銅[57-59]、甲硫氨酸鋅[60]和甲硫氨酸酪[61]。此外,在培養(yǎng)基中添加半胱氨酸乙硫氨酸等化合物均可使Met的產(chǎn)量顯著調(diào)高,如KUMAR等[62]在培養(yǎng)基中添加半胱氨酸,可使Met產(chǎn)量從2.34 g/L提高到3.39 g/L。金利群等[63]的研究發(fā)現(xiàn)通過(guò)控制發(fā)酵培養(yǎng)基中Na2S2O3的添加量,可顯著提高M(jìn)et的產(chǎn)量;當(dāng)Na2S2O3的添加量在3 g/L時(shí),Met產(chǎn)量可提高41.3%。
5.2 培養(yǎng)條件對(duì)Met合成的影響
在發(fā)酵過(guò)程中,除了合理的營(yíng)養(yǎng)元素外,培養(yǎng)條件也是影響Met產(chǎn)量的關(guān)鍵,包括pH、溫度及攪拌速率等。在微生物合成Met的過(guò)程中,pH值會(huì)直接影響代謝途徑中酶的活性。ZHOU等[13]研究發(fā)現(xiàn),pH值為7時(shí),E.coli發(fā)酵生產(chǎn)Met產(chǎn)量達(dá)到最高,分別比pH值6.5和7.5時(shí)提高37.78%和18.10%,表明中性pH值有助于E.coli發(fā)酵生產(chǎn)Met。溫度影響微生物發(fā)酵生長(zhǎng)主要涉及兩個(gè)方面即影響菌體自身生長(zhǎng)活力、誘導(dǎo)溫度影響代謝途徑蛋白質(zhì)表達(dá)水平和相應(yīng)酶活性[64]。有研究指出,用E.coli作為生產(chǎn)菌株生產(chǎn)Met最適宜溫度是28 ℃,此時(shí)參與Met生物合成的關(guān)鍵酶活性增強(qiáng),Met產(chǎn)量為1.24 g/L,較33 ℃和25 ℃時(shí)高1.09倍和2.21倍。攪拌速率是影響細(xì)胞生長(zhǎng)和代謝產(chǎn)物生物合成的重要因素。攪拌速度在200~400 r/min最為適宜,當(dāng)攪拌速率為300 r/min時(shí)Met的產(chǎn)量最高[13]。
6 總結(jié)和展望
Met具有重要的工業(yè)價(jià)值。近幾年,隨著發(fā)酵工程、代謝工程和合成生物學(xué)的不斷發(fā)展和應(yīng)用,研究者們利用基因工程菌生產(chǎn)Met已經(jīng)成為Met開(kāi)發(fā)的戰(zhàn)略方向,但是基于目前的研究進(jìn)展,距離工業(yè)化生產(chǎn)仍然有待研究。Met合成途徑已經(jīng)被完全解析,合成途徑的代謝調(diào)控尤其是從頭合成途徑的調(diào)控多數(shù)已經(jīng)被揭示,但是在前體物質(zhì)H2S和天冬氨酸,以及輔因子NADPH和ATP的提供等方面的調(diào)控也是限制Met合成的關(guān)鍵因素,還需要進(jìn)一步探究。本課題組致力于微生物高產(chǎn)Met的探究,前期的研究發(fā)現(xiàn)對(duì)各個(gè)前體物質(zhì)合成綜合調(diào)控的解析和工程化,是提高M(jìn)et產(chǎn)量的關(guān)鍵。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國(guó)衛(wèi)生(2016年3期)2016-11-12 13:23:26
