廣東石門臺假林線的物種組成及優勢種種群特征
2020-12-16 10:10:06張琪杜建會李遠球李升發邵佳怡
生態環境學報 2020年10期
關鍵詞:物種
張琪,杜建會, ,李遠球,李升發 ,邵佳怡
1. 中山大學地理科學與規劃學院,廣東 廣州 510275;2. 廣東省城市化與地理環境空間模擬重點實驗室,廣東 廣州 510275;3. 廣東石門臺國家級自然保護區管理局,廣東 英德 513000;4. 廣州地理研究所/廣東省地理空間信息技術與應用公共實驗室,廣東 廣州 510070;5. 廣東省遙感與地理信息系統應用重點實驗室,廣東 廣州 510070
在亞熱帶山區,矮曲林作為山頂重要的生態屏障,與其分布海拔之下的竹林、亞熱帶針葉林及常綠闊葉林等組成了相對穩定的生態系統,其物種組成及群落動態對于山地生態系統的水源涵養、水土保持、生物多樣性保護及碳儲存等具有非常重要的意義(Grafius et al.,2015;姚永慧等,2017)。但受自然或人為活動干擾,矮曲林被砍伐或燒毀,山頂因缺乏植被保護,加上坡度較陡和強降雨影響,使得該處土壤侵蝕加劇,土層變薄、養分流失。此外,山頂氣溫較低、輻射強烈、風力強勁、蒸發量大,進而使得森林的次生演替過程受到很大的抑制,原本發育森林的地方被灌叢或草甸所占據,森林則只能出現在距離山頂一定范圍的高度(Grubb,1971;姚永慧等,2017;張琪等,2018)。山頂灌草叢與森林接觸之處形極規則,視同橫線,前人將這種由山頂效應形成的突變,且分布在氣候林線之下的森林邊界稱為假林線(Gersmehl,1973;葉居新,1989)。
假林線在亞熱帶山地普遍存在,許多學者針對其起源、物種組成和土壤特性等展開調查,對于假林線的群落更新動態進行了初步研究,發現在人類活動中止后,受物種生物學特性和區域生境影響,假林線向上緣草坡均存在不同程度的入侵,但變化不顯著(徐捷等,2010;Bowman et al.,2017;張琪等,2018)。樹木的種子萌發、幼苗生長和幼樹的形成等受樹種組成、生境特征及干擾競爭等多個因素影響,任何一個環節出現障礙都可能會影響到森林的天然更新(趙高卷等,2016;張小鵬等,2018)。但假林線分布的區域地處偏遠、交通不便,在一定程度上限制了對其進行大范圍的深入調查,目前尚不清楚假林線附近的樹木在人類活動中止后能否持續更新,難以準確評估中國亞熱帶山地森林的恢復潛力(張金泉,2016)。種群特征可反映其個體數量的分配情況、形成過程和發展趨勢,也可表征樹種對生境的適應策略,這對揭示種群的更新和維持機制具有非常重要的意義(費世民等,2008;Li et al.,2020)。本文通過對廣東石門臺國家級自然保護區內的假林線典型分布區木龍頂樣地進行群落物種調查,并對其喬木層優勢種的樹高、胸徑和生物量等進行測量及采樣分析,有助于理解人類活動中止后,亞熱帶山地森林的次生演替過程,這對于準確評估亞熱帶山地生態系統的服務價值也具有重要的意義。
1 材料與方法
1.1 地理位置
廣東石門臺國家級自然保護區位于廣東省英德市西北部,北與韶關市的乳源、曲江縣相鄰,西起波羅鎮的赤泥嶺,東至沙口鎮的江溪,東西寬約43.66 km,南北長約15.48 km,總面積335.55 km2。地勢西北高、東南低,兼有平原、丘陵、山地和喀斯特地形的多樣性地貌,最高峰船底頂海拔1586.8 m(張琪等,2018)。早期因大煉鋼鐵,大面積常綠闊葉林被砍伐用于燃料,加上后續村民伐木建屋、毀林種果、種竹筍,使得原始森林遭到嚴重破壞。英德市人民政府首先在 1998年于石門臺成立縣級自然保護區,2012年將其升級為國家級自然保護區,并全面實行封山育林,嚴禁砍伐等措施,使得次生植被得到了良好的保護,除少量的戶外活動外,核心區無其它居民居住(張金泉,2017)。
1.2 數據來源及研究方法
1.2.1 植物群落調查
在線路踏查的基礎上,選取木龍頂樣地作為研究區(24°28′42.1″—24°28′43.0″N,113°14′15.0″—113°14′17.2″E),其海拔為 1118 m,土壤類型以山地草甸土為主。采用相鄰格子法,于2017年11月中旬沿著假林線邊緣設置6個10 m×20 m樣方,并在樣方內沿對角線設置2 m×2 m小樣格。考慮到研究區生境嚴酷,疏齒木荷主要以幼苗為主,且后勤難以補給,因此在保證數據測定精度的基礎上,以樹高≥2 m且胸徑≥2.5 cm作為喬木層起測標準,進行每木調查并記錄種名、高度、胸徑、枝下高、冠幅和林分郁閉度,胸徑小于2.5 cm的幼苗只統計其高度及數量;對于灌木層則記錄其種名、株數、平均高度和蓋度;草本層記錄其種名、平均高度和蓋度;藤本植物記錄其種名。用GPS測定海拔和經緯度,用坡度計測定坡向和坡度。
1.2.2 重要值
喬木層和灌木層重要值(IV)的計算基于相對多度(ra)、相對頻度(rf)和相對蓋度(rc);草本層以相對高度(rh)替換相對多度(ra)。各參數及植被蓋度(C)分別采用以下公式計算,其中真蓋度指喬木胸徑處的截面積,基蓋度指灌木基徑處的截面積(喬木層和灌木層重要值根據公式 (6) 計算;草本層重要值根據公式 (7) 計算):

上式中,W為實際測量的植物胸徑;

上式中,mi為第i個物種的株數,Σmj為全部物種的總株數;

上式中,fi為第i個物種的頻數,Σfj為全部物種的頻數之和;

上式中,ci為第i個物種的蓋度,Σcj為全部物種蓋度之和;灌木層直接用估算的蓋度進行計算;

上式中,hi為第i個物種的平均高度,Σhj為全部物種的平均高度。

1.2.3 多樣性指數
本研究采用重要值作為優勢度指標以確定植物群落各層的優勢種(王伯蓀等,1996),以辛普松指數(Simpson Index,D)和香農-維納指數(Shannon-Wiener Index,H′)對喬灌層及群落的多樣性水平進行分析。兩種指數計算公式如下:

上述兩式中,Pi=Ni/N,其中Ni為樣方內第i個物種的個體數,N為樣方內所有物種的個體數之和。
1.2.4 疏齒木荷生物量測定
為避免對固定樣方內的植被造成干擾,在樣地附近選取不同徑級的疏齒木荷(Schima remotiserrata)作為標準株。考慮到絕大多數的植株胸徑均小于6.5 cm,且樣地位于保護區范圍內,無法進行大規模破壞性采樣,因此對胸徑≤6.5 cm的樣本,共選擇了 11株,使用小鐵鏟沿著其根系(主要分布在0—20 cm)的延伸方向,采用全挖法將其整株挖出。對于>6.5 cm的樣本,以胸徑間隔2 cm為徑級劃分標準,在樣地附近選擇標準木28株,現場砍伐,以充分代表疏齒木荷的種群特征。所有樣品按器官分為樹干、樹枝和樹葉,并在樣地迅速稱取鮮重,用于構建單株地上生物量與其樹高及胸徑之間的關系。由于山頂缺乏水源,胸徑≤6.5 cm的植株根系泥土粘連較多,因此將其全部帶回,仔細清理干凈后,測定其鮮質量,并計算根冠比,用于估算胸徑>6.5 cm的植株地下生物量。研究區山高路遠,運輸不便,因此將植株地上樣品分器官充分混合,依照比例選取部分樣品帶回實驗室。所有樣品均在烘箱內設置70 ℃烘干(24 h)至恒質量,分別記錄樹干、樹枝、樹葉和樹根生物量,并計算其干鮮重比。最后結合不同徑級的立木密度,估算木龍頂樣方內的疏齒木荷各個徑級的生物量及總生物量。
1.3 數據分析
相關數據采用SPSS 13.0(SPSS,Chicago,IL,USA)進行統計分析,作圖使用Origin 9.0(Origin,Northampton,USA)進行繪制。
2 結果與分析
2.1 群落組成及物種特征
2.1.1 群落外貌
木龍頂樣地植物群落主要以亞熱帶常綠闊葉林為主,外貌整齊,林冠濃密,郁閉度達0.85,極少林窗。群落組成中雖有少數落葉樹種,但并不構成明顯的落葉季相,常與一定數量的針葉樹種混交成林。林冠呈圓形或橢圓形,喬木分層不明顯,植株低矮,群落高度普遍在 3—5 m,平均樹高(3.5±0.9) m,平均胸徑 (4.7±2.8) cm,枝下高平均約(1.3±0.6) m,偶見少數針葉樹種高于林冠之上,但最高亦不超過7 m。本次調查的1200 m2樣地中,高度在5 m以上的樹木僅26株。群落內胸徑≥2.5 cm的立木平均密度為3900 plant·hm-2,林木生長稠密,多小型或中型葉、革質且多茸毛,植株通常分枝低矮且側枝發育扭曲,樹干多附生有苔蘚類植物。藤本種類較少,僅見少量草質和木質藤本。根據以上群落基本特征,可將該群落劃入山頂苔蘚矮曲林。
2.1.2 群落組成與物種多樣性
(1)物種組成
木龍頂樣地內共有維管植物53種,隸屬33科,43屬(表1)。其中,蕨類植物1科1屬1種,裸子植物2科2屬2種,被子植物30科40屬50種。群落物種組成以杜鵑花科6種最多,其次是樟科4種,禾本科、冬青科和百合科各3種,茜草科、金縷梅科、山茶科、山礬科、木蘭科和野牡丹科各 2種,其余各科各1種。以亞熱帶表征科為數量優勢科,充分體現了植被的水平地帶性分布特征。
木龍頂樣方喬木層中的疏齒木荷重要值最高(26.01%),其次為五列木和華南五針松(表2)。樣方喬木層平均樹高普遍較小,華南五針松的平均高度(4.39 m)略高于五列木(3.44 m)和疏齒木荷(3.21 m),但其數量遠低于后兩者,相對多度較小。樣方喬木層中五列木數量最多,相對多度最大,但其相對蓋度及相對顯著度明顯小于疏齒木荷,因而后者成為樣方喬木層的優勢種。
灌木層中五列木分布均勻,生勢良好(表3),其數量雖略少于廣東杜鵑(Rhododendron kwangtungense),但總截面積是后者近10倍,故相對蓋度在木龍頂樣地的灌木層中位于首位,可判斷五列木為樣方內灌木層的優勢種,而其它物種在灌木層中的生勢則較為均衡。
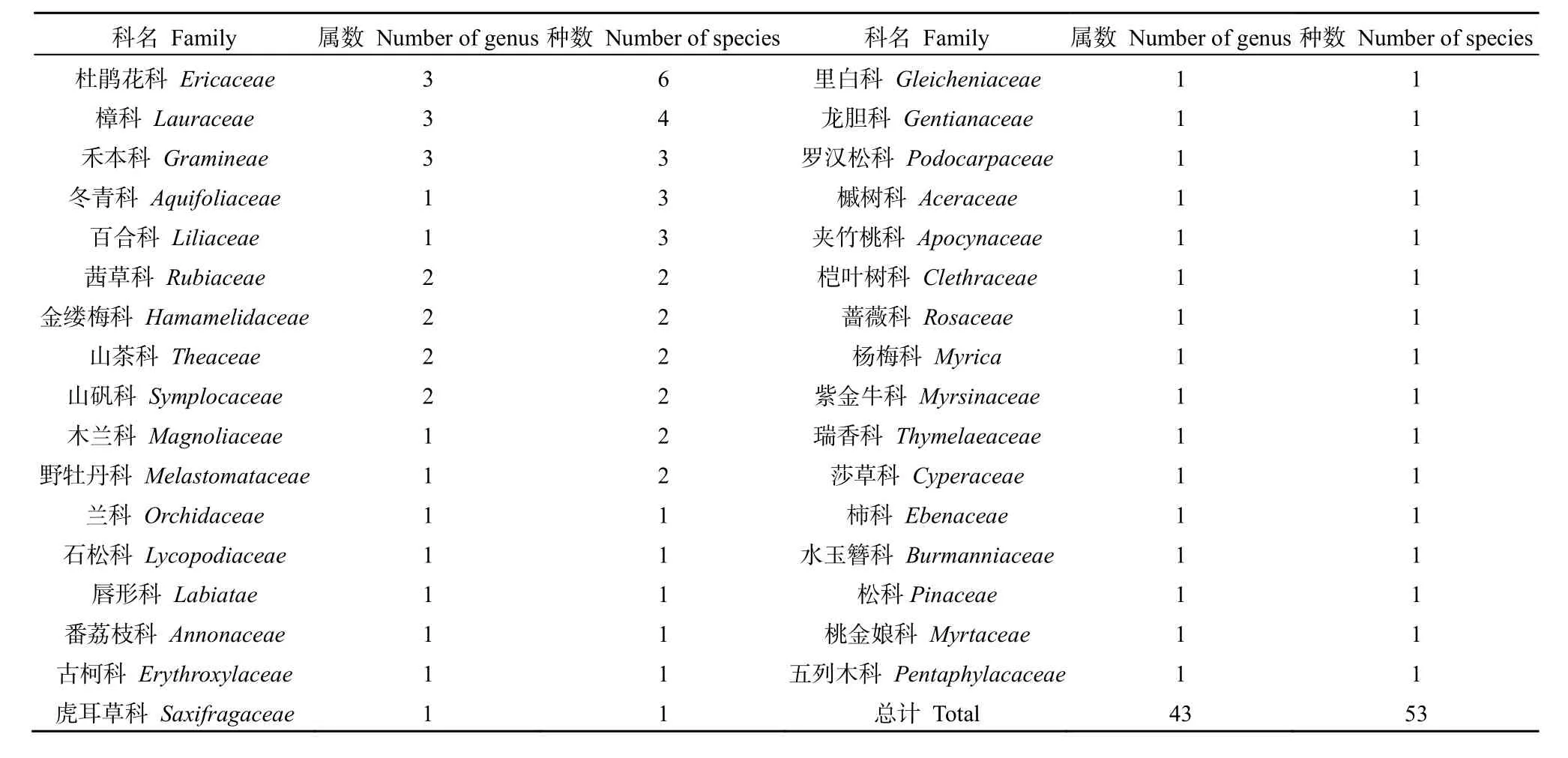
表1 群落維管植物分布特征Table 1 The distribution characteristics of community of vascular plants
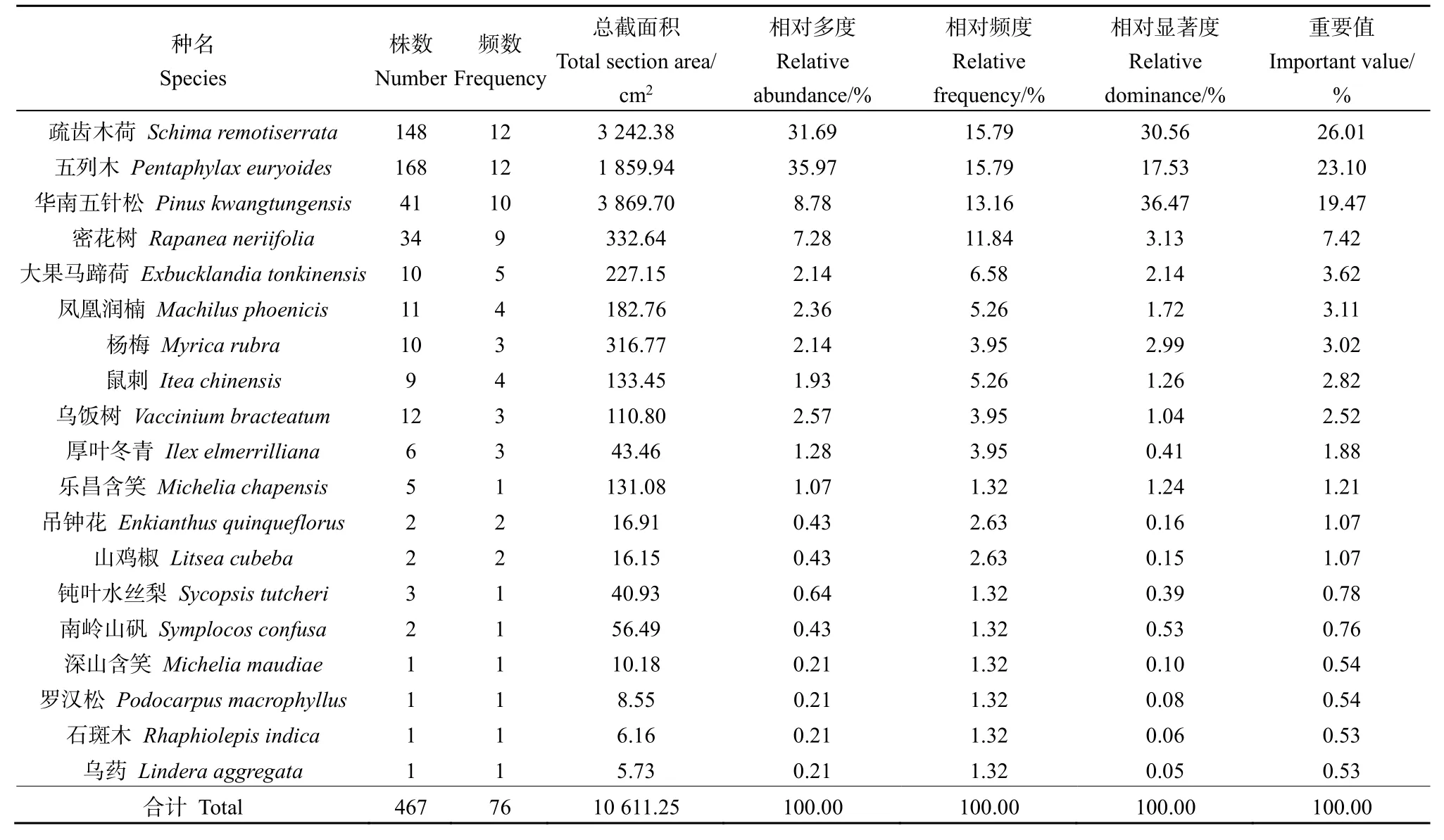
表2 木龍頂樣方喬木層特征值Table 2 Important value of tree layer plots at Mulongding Mountain
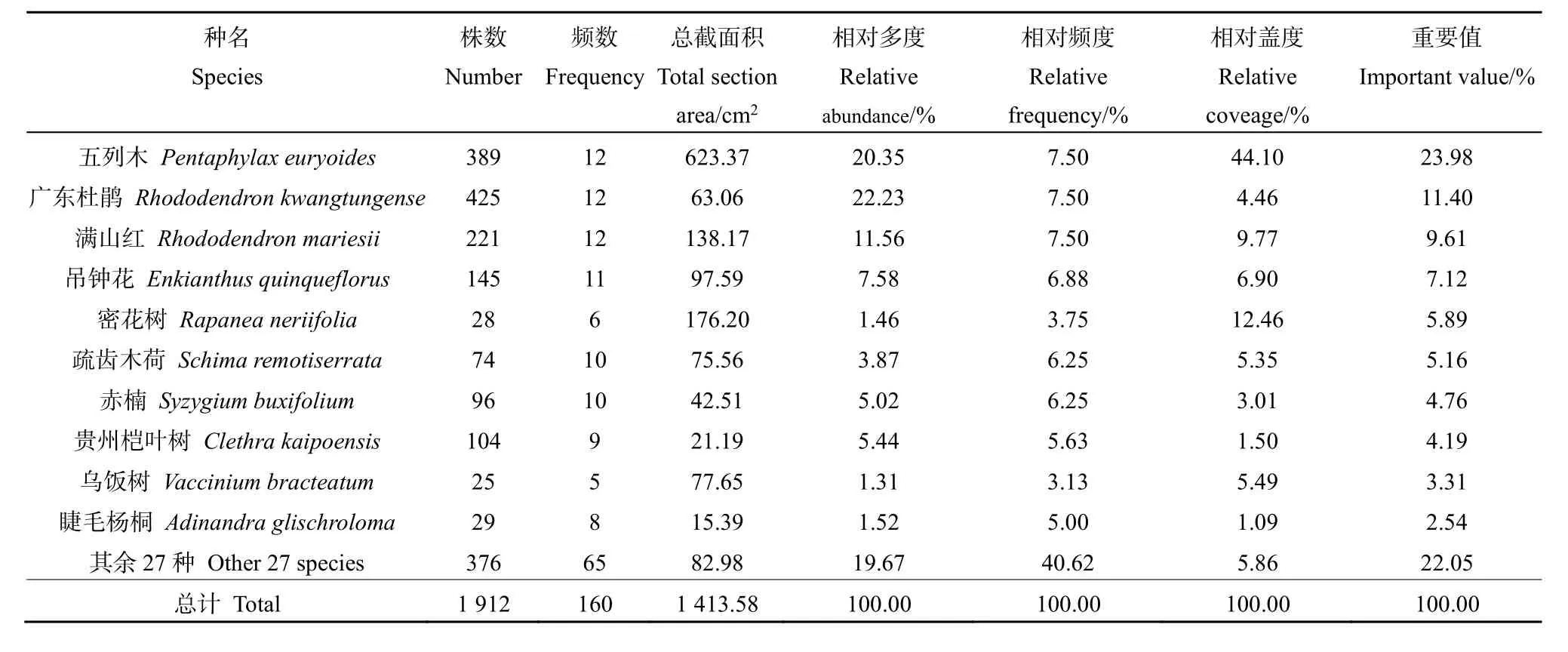
表3 木龍頂樣方灌木層特征值Table 3 Important value of shrub layer plots at Mulongding Mountain
草本層中的野古草(Arundinella anomala)分布均勻(表4),且生勢良好,其相對頻度與相對蓋度均居于樣方首位且遠大于其余物種,重要值達到47.84%,成為該樣方草本層的優勢種。
此外,木龍頂樣方內的層間植物還包括藤本灌木鏈珠藤(Alyxia sinensis)、土茯苓(Smilax glabra)、草質藤本藤石松(Lycopodiastrum casuarinoides)、攀援灌木菝葜(Smilax china)、暗色菝葜(Smilax lanceifolia)、羊角藤(Morindaumbellata)等6種。
根據以上調查結果可知,樣方內共記錄有 19種喬木,群落優勢種為疏齒木荷,其截面積為88427 cm2·hm-2。灌木層記錄物種數 37種,平均高度(1.48±1.33) m。草本層記錄物種數8種,平均高度(0.38±0.24) m。依據各層物種的重要值(表2—4),可將該樣地植物群落命名為疏齒木荷-五列木-野古草群叢(Ass.S.remotiserrata-Pentaphylax euryoides-Arundinella anomala)。
(2)物種多樣性指數
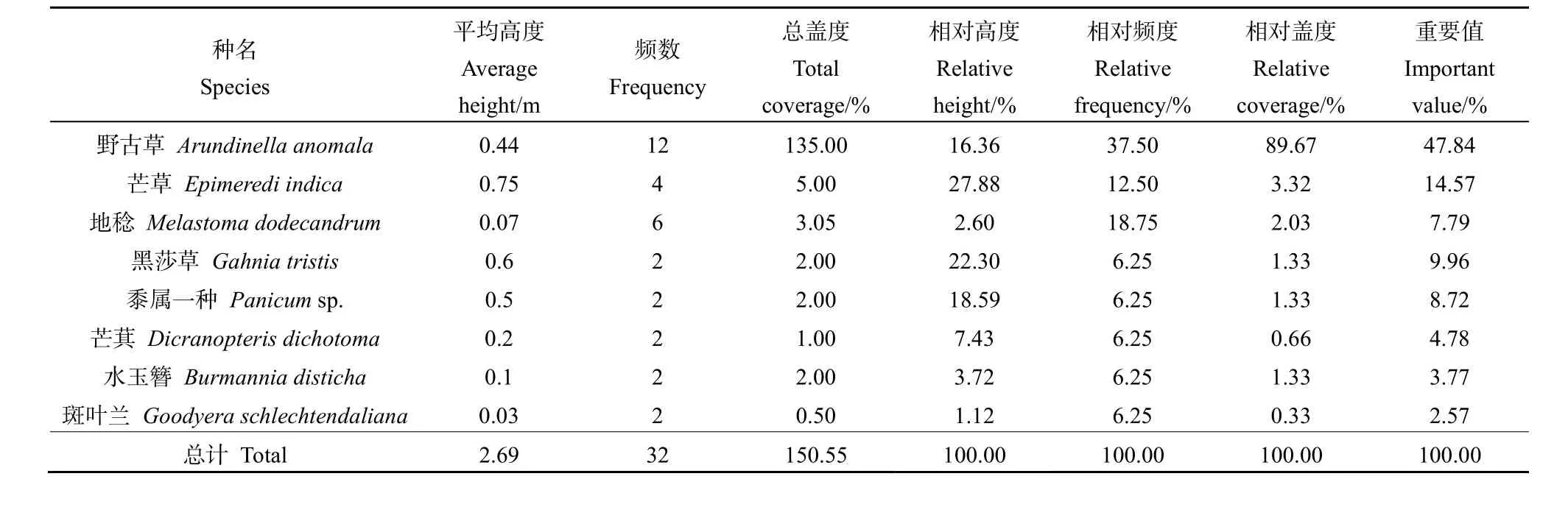
表4 木龍頂樣方草本層特征值Table 4 Important value of herb layer plots at Mulongding Mountain
樣方植物群落的物種多樣性指數D為0.882,H′為 3.652,表明群落物種多樣性水平較高。其中喬木層D為0.754,H′為2.624;灌木層D為0.879,H′為 3.679。兩種多樣性指數在喬木層和灌木層變化趨勢一致,均表現為灌木層的物種多樣性水平高于喬木層。
2.2 疏齒木荷的種群特征
2.2.1 疏齒木荷種群的結構特征
樣地內疏齒木荷種群的平均高度為 (3.22±0.84)m,以小于1.0 m的植株為主,其數量占疏齒木荷種群的88.8%,高度在5 m以上的僅5株,最高為7.0 m,群落整體矮化,分枝較低(圖 1A)。疏齒木荷種群的平均胸徑為 (4.60±2.25) cm,以胸徑≤2.5 cm的植株為主,密度達到15417 plant·hm-2,占疏齒木荷種群的92.3%。隨著胸徑增長,其個體數量呈現出顯著的下降趨勢,胸徑>2.5 cm的植株較少,其密度僅有1283 plant·hm-2。其中胸徑在2.5—6.5 cm范圍內的植株占6.8%,而胸徑大于6.5 cm以上的植株數量僅占總數的0.9%(圖1B)。
2.2.2 疏齒木荷種群的生物量特征
不同徑級的疏齒木荷生物量差異較大,除樹葉外,其余器官的生物量均隨著徑級的增加呈現出先上升后下降的趨勢(表5)。胸徑≤6.5 cm的疏齒木荷總生物量為4.73 t·hm-2,占總生物量的51.88%,而胸徑>6.5 cm的疏齒木荷盡管株數較少,但單株生物量大,占到總生物量的48.12%。疏齒木荷各器官的生物量占比也有所不同,從大到小依次為樹干、樹根、樹枝和樹葉。樹干生物量占總生物量的比例達52.20%,而樹葉生物量僅占總生物量的6.01%。本文采樣時間為11月中旬,恰好對應了疏齒木荷的換葉期,可能低估了樹葉的生物量。依據樣方的立木密度調查數據,基于胸徑和樹高模型估算可得研究區的疏齒木荷地上生物量為7.11 t·hm-2。疏齒木荷根冠比波動范圍在 0.15—0.48之間,且隨著胸徑的增大先快速減小,后逐漸穩定,依據其根冠比可估算疏齒木荷的總生物量為9.10 t·hm-2。
3 討論
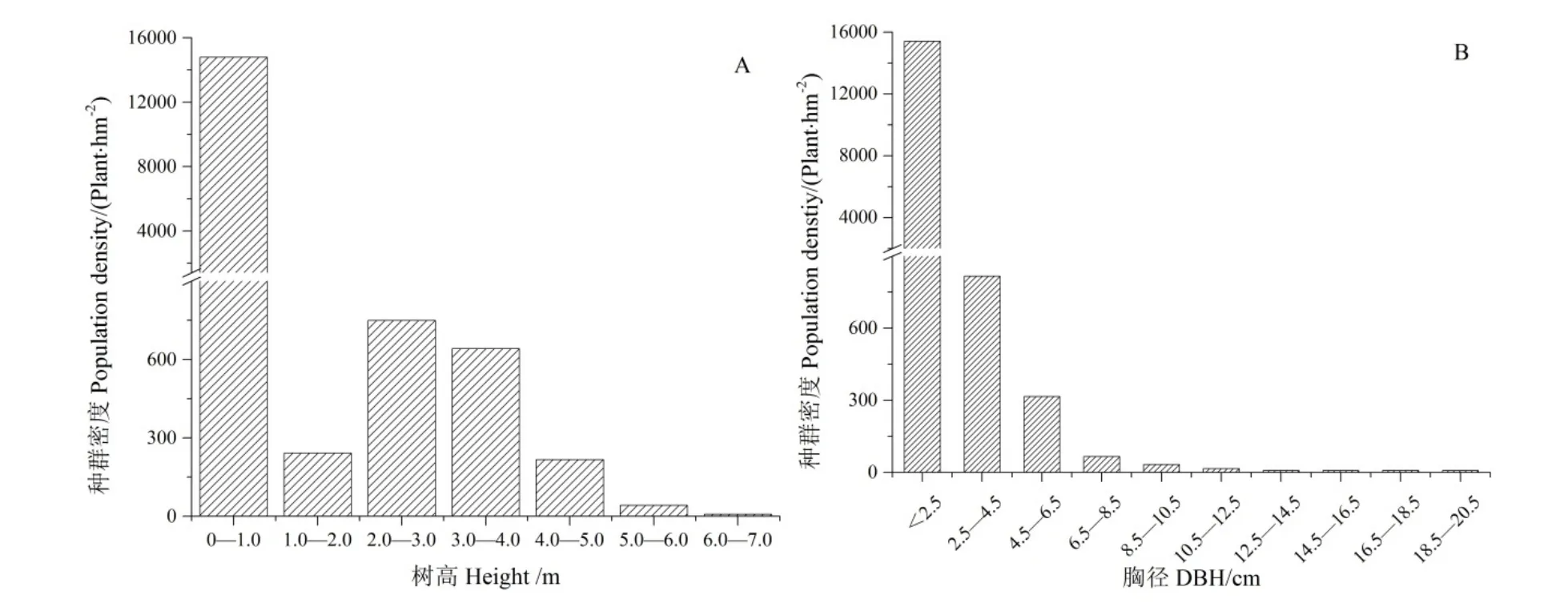
圖1 木龍頂樣方中的疏齒木荷種群胸徑及樹高頻數分布特征Fig. 1 Population density in height (A) and DBH-class (B) of S. remotiserrata in Mulongding Mountain
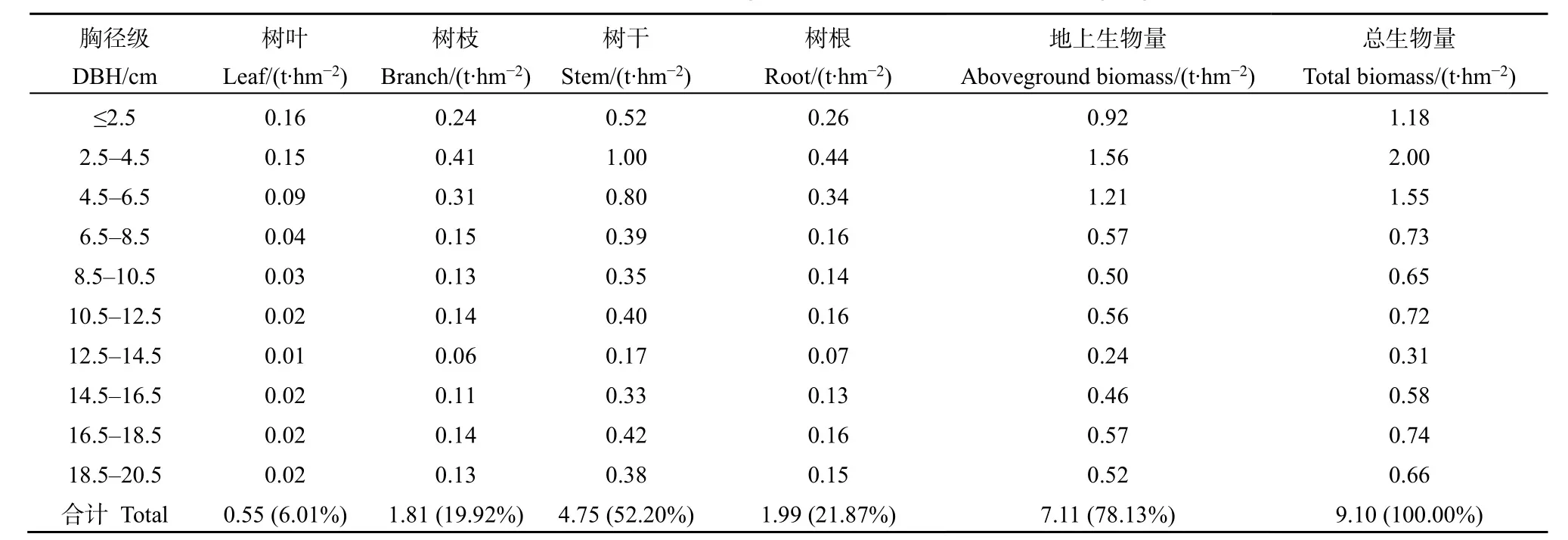
表5 木龍頂樣方不同徑級的疏齒木荷各器官生物量特征Table 5 Biomass of S. remotiserrata among different diameter classes in Mulongding Mountain
森林群落的物種組成受生境影響明顯(陳林等,2010)。對木龍頂假林線樣方的調查發現,其物種組成主要以喬灌木為主,草本和藤本植物相對較少。這與王厚麟等(2008)對中國亞熱帶山頂矮林的研究結果比較一致。假林線林冠層的郁閉度較高,往往造成林下光照不足,且枯枝落葉多,不利于草本和藤本植物種子萌發及根系下扎,從而抑制其后續生長(陳林等,2010)。樣方內的喬木層高度普遍較矮,且扭曲生長,其平均樹高為(3.5±0.9) m。這與陳林等(2010)對南嶺國家自然保護區的山頂矮林調查結果較為接近,其喬木層的平均高度為3.23 m,以5 m以下的個體為主。Shiels et al.(2001)對科羅拉多Goliath山地的刺果松(Pinus aristata)和恩格曼云杉(Picea engelmannii)矮曲林研究也發現,其高度大小在0.5—3 m之間。假林線多分布在保護區的山頂或山脊部位(張琪等,2018),該處風力強勁、濕冷多霧、溫差大,致密且低矮的林冠層可有效降低風力造成的機械損傷,并能減少輻射加熱和土壤熱通量交換,林冠層下的熱量散失較慢,其葉片組織和根際溫度常高于直立生長的高大樹木,有利于提高矮曲林的光合作用水平,緩解極端氣溫對樹木生長的影響(Grubb,1971;McIntire et al.,2016;Ka?par et al.,2017)。
樹木在不同的生長階段受限因素存在差異,進而影響到其種群的結構特征(王樹力等,1993)。對疏齒木荷種群的樹高特征研究發現,其呈現為反“J”型,以高度小于1 m的植株為主,1—2 m高度的植株占比急劇減小。說明疏齒木荷的天然更新存在瓶頸現象,其小于1 m的植株死亡率較高,只有少部分植株能夠存活,并進入到下一生長階段。這與宋立奕等(2013)對云南元謀干熱河谷膏桐(Jatropha curcas)種群的天然更新研究較為一致,其認為膏桐幼苗可能與高度近1 m的多年生草本植物扭黃茅(Heteropogon contortus)產生了強烈的競爭,導致高度小于1 m的膏桐幼苗生長受到了很大的抑制。木荷屬植物的更新主要與自然或人類活動形成的林窗有關(Tang et al.,2020)。對研究區的調查發現,草本植物中重要值較高的如芒草等平均高度可達到0.75 m,灌木層平均高度為1.48 m,其與高度小于1 m的疏齒木荷幼苗共同競爭空間和光照等,在一定程度上可能抑制了疏齒木荷的幼苗成長為幼樹。但一旦突破這一高度,其在與草本及灌木的競爭過程中將占據優勢,且生長力明顯提高,進而更新形成相對穩定的種群(胡爾查等,2013)。對樣方內的疏齒木荷調查也證實,高度在 2—3 m的植株比例再次顯著增加(圖 1A)。因此從幼苗向幼樹階段的轉變就成為假林線附近疏齒木荷更新的關鍵,未來需要通過適當的幼苗撫育,促使其持續更新(Tesfaye et al.,2002)。5 m以上的個體占比僅有0.25%,主要在于隨著幼樹逐漸成長為成熟林,其對資源的需求及對環境脅迫的敏感程度均在增加,加上逐漸衰老,導致大樹存活的數量極為稀少(宋立奕等,2012)。
幼苗更新的早期受種源制約,而之后的更新過程則與生物和非生物要素均有關(邵佳怡等,2019)。對疏齒木荷種群的胸徑研究發現,樣方中92.6%的疏齒木荷胸徑小于2.5 cm,極少有植株胸徑大于 10.5 cm,這與尹愛國等(2004)和陳林等(2010)對粵北山頂矮林的研究結果較為一致,其喬木層大部分植株的胸徑均在10 cm以下。這主要與疏齒木荷的生物學特性有關,其單株結實量大,且隨風擴散能力強(Tang et al.,2020),可確保疏齒木荷幼苗更新充分,也可能是其成為假林線森林群落優勢種的關鍵(宋立奕等,2013)。但隨著幼苗生長,個體對資源的需求也不斷增加,在資源有限的情況下,進而導致其個體數量隨著胸徑增加出現銳減(趙欣鑫等,2017),其中胸徑小于等于2.5 cm 的植株密度達到 15417 plant·hm-2,而大于 2.5 cm的植株密度僅有1283 plant·hm-2。疏齒木荷的種群結構因而表現為基部較寬、頂部極窄的金字塔型結構,樹木的更新和死亡率在短時間尺度上達到了平衡,種群增長性較低(宋立奕等,2012;Chhetri et al.,2015;吳語嫣等,2019)。
假林線所在的矮曲林生長受生境影響較大,其普遍低矮,且扭曲生長,因此基于胸徑和樹高的生物量估測模型能夠更好地估算疏齒木荷的生物量(Poorter et al.,2012;Hunter et al.,2013)。本文的估算結果表明,疏齒木荷的地上生物量為 7.11 t·hm-2,其遠低于付甜等(2014)對三峽庫區八種亞熱帶森林喬木層地上生物量的估算結果,說明人類活動中止后,假林線所在的植物群落長勢較差,樹木的天然更新不容樂觀。疏齒木荷的根系生物量占總生物量的比例為 21.87%,略大于陳章和等(1996)采用全挖法對黑石頂南亞熱帶常綠闊葉林地下部分生物量的測定結果(20.54%)。這主要與山頂土層淺薄和土壤貧瘠有關,疏齒木荷的幼苗一開始傾向于將更多地生物量分配到根系,以確保水分和養分的供應,但同時也削弱了其與相鄰草灌木競爭空間和光照的能力,不利于其后續的更新(張文輝等,2008)。依據根冠比估算可知,本研究區疏齒木荷的總生物量為9.10 t·hm-2,也遠低于Fang et al.(1998)對亞熱帶森林典型樹種的總生物量估算結果,主要與當前疏齒木荷種群以幼苗為主,成熟林占比相對較少有關,其碳儲存相對于同氣候區偏低,碳匯潛力較大。未來若能對現有樹木進行適當的人工撫育和科學管理,促進其幼苗向成熟林的演替,則有望增強這一地區的碳匯功能(胡海清等,2015)。Bastin et al.(2019)對全球森林恢復的評估中也指出,因早期退化而以稀疏灌叢或草地為主的亞熱帶山地可能是未來全球森林恢復的潛在區域之一,并有望緩解當前的氣候變化。
4 結論
本文通過對廣東石門臺國家級自然保護區假林線群落的物種組成及喬木層優勢種疏齒木荷的種群特征進行研究,結果表明:
(1)受山頂效應影響,假林線的群落結構簡單、總體矮化,形成了典型的亞熱帶山頂常綠苔蘚矮曲林,其喬灌草的優勢種分別為疏齒木荷、五列木和野古草。
(2)疏齒木荷的高度分布表現為反“J”型,胸徑以中小徑級為主,其幼苗更新充分,但死亡率較高,大徑級植株極少,形成了典型的底部較寬、頂部極窄的金字塔型結構,種群增長性較低,其中從幼苗到幼樹的過渡成為研究區假林線附近疏齒木荷種群更新的關鍵。
(3)疏齒木荷的地上生物量和總生物量均低于同氣候區的森林,其林分質量較差,未來需要通過適當的人工撫育,提高成熟林所占比重,從而增強這一地區的碳匯功能。
氣候變暖背景下,不同物種的響應存在差異,進而會影響到假林線附近的物種組成及其種群特征。未來需要在亞熱帶山地選擇典型樣地,采用固定樣方對于假林線群落的物種組成、種群特征和非生物要素等開展長期系統的定位觀測,研究不同撫育措施下,假林線優勢種疏齒木荷幼苗的更新潛力,這些都有助于理解亞熱帶山頂矮林的次生演替過程,并對準確評估其生態系統的服務價值也尤為重要。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
