渭河階地不同生境反枝莧種子萌發(fā)的熱量需求研究
2020-12-14 07:07:46王雅馨魏永勝劉嘉聰
西北農(nóng)業(yè)學(xué)報(bào) 2020年12期
王雅馨,楊 婷,魏永勝,劉嘉聰
(1.西北農(nóng)林科技大學(xué) 生命科學(xué)學(xué)院,陜西楊凌 712100;2.西北農(nóng)林科技大學(xué) 植物保護(hù)學(xué)院,陜西楊凌 712100;3.西北農(nóng)林科技大學(xué) 機(jī)械與電子工程學(xué)院,陜西楊凌 712100)
入侵雜草危害是全球重大生態(tài)問(wèn)題之一[1]。反枝莧(AmaranthusretroflexusL.)為莧科、莧屬1a生草本植物,是一種典型的C4植物,具有高光合效率、低CO2補(bǔ)償點(diǎn)的光合特性[2],以及生長(zhǎng)快、籽實(shí)多等生物學(xué)特性,一株成株的反枝莧種子產(chǎn)量高達(dá)1萬(wàn)~3萬(wàn)粒[3]。同時(shí),反枝莧具有較強(qiáng)的表型可塑性[4],可通過(guò)增加自身的生態(tài)幅和耐受力來(lái)適應(yīng)環(huán)境因子頻繁變化的生境,如反枝莧在晝夜溫度為23℃/13℃和20℃/10℃條件下單株結(jié)實(shí)量分別為5465粒和1655粒,單株干質(zhì)量分別為2.1g和0.7g[5]。反枝莧是世界廣布性入侵雜草[6],也是中國(guó)目前莧屬植物(共9種)中入侵?jǐn)?shù)量居首位和分布最為廣泛的一種[7],對(duì)農(nóng)田的危害面積在中國(guó)暖溫帶高達(dá)36%[8]。反枝莧可在棉花、玉米[9]、小麥[10]、煙草、大豆[11]等多種作物的田地生長(zhǎng),通過(guò)化感作用、與作物競(jìng)爭(zhēng)光照[12]、水分[13]和營(yíng)養(yǎng)物質(zhì)[14]以及作為病蟲(chóng)害的傳播介體等,造成作物大量減產(chǎn)[15],導(dǎo)致嚴(yán)重的經(jīng)濟(jì)損失。例如,棉花田中每平方米增加1株反枝莧可導(dǎo)致棉花單株鈴數(shù)減少20.81%~84.98%[16]。此外,反枝莧對(duì)家畜健康[17]、人類健康[18]以及生態(tài)環(huán)境都有一定的危害。
全球氣候變暖條件下,作為C4植物的反枝莧的生態(tài)適應(yīng)情況鮮有報(bào)道。種子萌發(fā)階段是反枝莧成功入侵多類型生境的關(guān)鍵階段,苗期則是實(shí)施防控的關(guān)鍵時(shí)期[19]。種子萌發(fā)所需的時(shí)熱累積模型參數(shù)具有一定的生理和生態(tài)意義,能夠有效預(yù)測(cè)不同溫度條件下的種子萌發(fā)時(shí)間及種苗出土?xí)r間[20]。但目前關(guān)于反枝莧萌發(fā)的熱量需求尚不清楚,無(wú)法準(zhǔn)確預(yù)測(cè)其在田間的萌發(fā)日期。因此,通過(guò)探究不同生境下反枝莧種子的萌發(fā)熱量需求,探討反枝莧對(duì)全球氣候變暖的響應(yīng),并預(yù)測(cè)其萌發(fā)日期,對(duì)進(jìn)一步掌握反枝莧的生物學(xué)特性,優(yōu)化反枝莧的防控方法,以提高防效有重要意義。為此,本研究擬利用積溫模型(thermal time model),定量估計(jì)不同生境下反枝莧種子的萌發(fā)熱量需求,預(yù)測(cè)反枝莧對(duì)全球氣候變化的響應(yīng)及萌發(fā)日期,進(jìn)而為反枝莧的適時(shí)防除提供理論 依據(jù)。
1 材料與方法
1.1 材 料
反枝莧種子于2019年9月采自陜西省咸陽(yáng)市楊凌區(qū)渭河階地(圖1),該區(qū)位于西北農(nóng)林科技大學(xué)北校區(qū)(北緯 34°15′,東經(jīng) 108°03′,海拔:519 m,即頭道塬,以N表示)、南校區(qū)(北緯 34°17′,東經(jīng) 108°04′E,海拔:459 m,即二道塬,以S表示)以及渭河河岸旁(北緯34°13′,東經(jīng) 108°07′,海拔:438 m,即河灘地,以R表示),每個(gè)地隨機(jī)取3個(gè)樣點(diǎn)(間距>300 m),每樣點(diǎn)各3株。種子去雜、自然風(fēng)干后保存于牛皮紙信封,室溫通風(fēng)干燥處儲(chǔ)藏,待用。
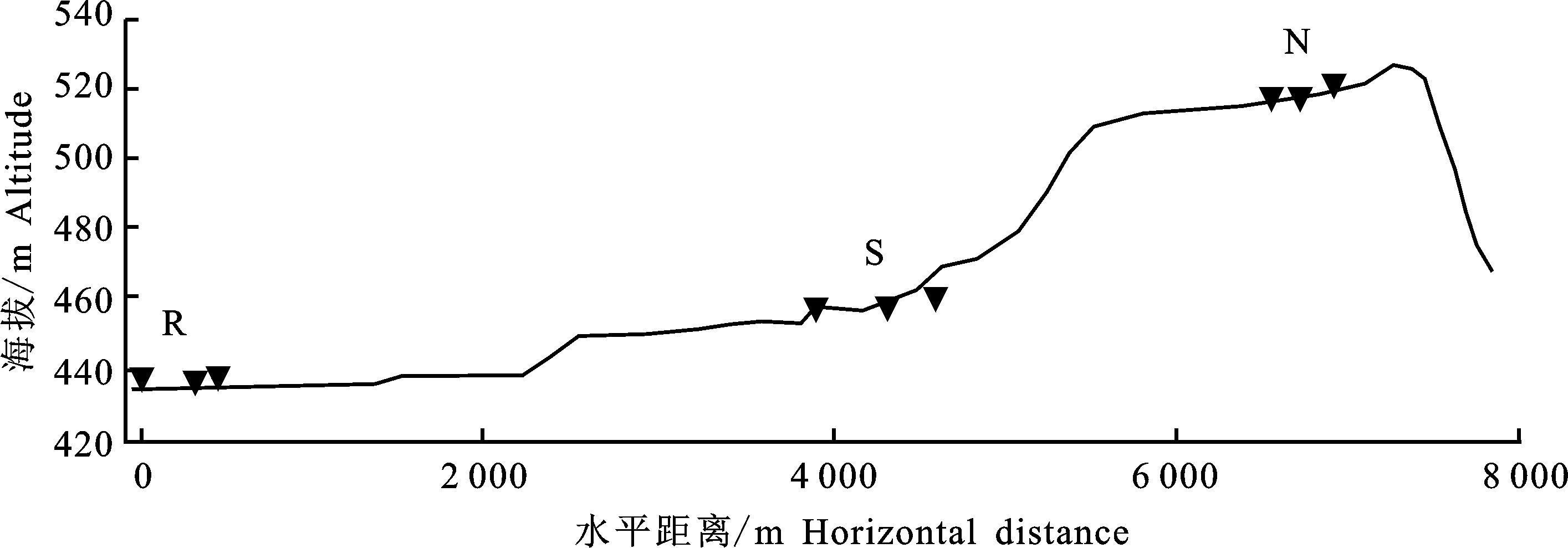
R.河灘地;S.二道塬;N.頭道塬;下同
1.2 方 法
1.2.1 種子質(zhì)量評(píng)價(jià) 種子千粒質(zhì)量與含水量的測(cè)定:隨機(jī)選取200粒種子進(jìn)行測(cè)量,重復(fù)3次(共600粒),稱量后計(jì)算千粒質(zhì)量。另取3份各100粒種子,稱量后放入稱量瓶,置于80 ℃烘箱烘24 h后取出,于干燥器內(nèi)冷卻30 min至室溫,稱取干質(zhì)量后計(jì)算含水量。
種子活力評(píng)估:活力測(cè)定用TTC染色法[21],輕壓種子使種皮破裂,置于培養(yǎng)皿中,加入w=0.5%TTC溶液10 mL,置35 ℃黑暗條件下4 h,之后檢查種子的胚,統(tǒng)計(jì)染色結(jié)果。胚有光澤的鮮紅色且組織狀態(tài)正常的為有活力的種子。并計(jì)算活力,具體為:V=n/N×100%,式中V為種子活力,n為染色種子數(shù),N為供試種子總數(shù)。試驗(yàn)重復(fù)3次,每次50粒種子。
1.2.2 萌發(fā)試驗(yàn) 取3個(gè)生境的種子各50粒,用φ=75%的酒精消毒0.5 h,蒸餾水沖洗3次,置于鋪有雙層濾紙、直徑為9 cm的培養(yǎng)皿中,加5 mL蒸餾水后加蓋分別置于10、20、25、30、35和40 ℃的恒溫培養(yǎng)箱(上海福瑪,SPX-250B)內(nèi)進(jìn)行萌發(fā)試驗(yàn)。試驗(yàn)重復(fù)5次。每個(gè)培養(yǎng)箱中置1支UT330ABC溫度計(jì),采樣間隔0.5 h,測(cè)得實(shí)際溫度分別為10.2、19.2、24.7、28.9、36.5和39.6 ℃。自吸水后計(jì)時(shí),以種子胚根突破種皮1 mm為萌發(fā),每間隔2 h觀察并記錄萌發(fā)率。計(jì)數(shù)后的種子取出,至連續(xù)1周無(wú)種子萌發(fā)時(shí)結(jié)束試驗(yàn)。最終計(jì)數(shù)未萌發(fā)的種子,并評(píng)估種子萌發(fā)率(G)。G=ni/(N×V) ×100%,ni為i時(shí)刻萌發(fā)種子數(shù),N為供試種子總數(shù),V為活力。
1.3 數(shù)據(jù)分析
1.3.1 萌發(fā)過(guò)程的擬合 利用sigmaplot 14.0以經(jīng)過(guò)活力校正的累積萌發(fā)率(y)對(duì)種子吸水時(shí)間(x)做散點(diǎn)圖,利用Chapman 3 Parameter方程y=a(1-e-bx)c擬合,估計(jì)參數(shù)a、b、c。其中a為估計(jì)的最高萌發(fā)率,有生物學(xué)意義,因此本模型為半理論模型。
利用上述方程分別計(jì)算,當(dāng)累積萌發(fā)率y=20%、30%、…70%時(shí),所需萌發(fā)時(shí)間x的值,以1/x(萌發(fā)速率)對(duì)溫度作圖,最適溫度之前的各萌發(fā)率線性回歸方程的斜率為相應(yīng)的1/θT(g)。θT(g)為調(diào)整累積萌發(fā)率達(dá)到g%(相應(yīng)的萌發(fā)率,即y)所需要的積溫,所有θT(g)呈正態(tài)分布[22]。
1.3.2 調(diào)整累積萌發(fā)率達(dá)到50%所需積溫 θT(50)及積溫標(biāo)準(zhǔn)差σθT的評(píng)估 利用正態(tài)分布函數(shù)進(jìn)行評(píng)估:對(duì)應(yīng)的種群相對(duì)頻率呈正態(tài)分布,對(duì)其進(jìn)行標(biāo)準(zhǔn)正態(tài)離差轉(zhuǎn)換。以θT(g)為橫坐標(biāo),達(dá)到不同萌發(fā)率的概率密度Probg為縱坐標(biāo)做散點(diǎn)圖,并進(jìn)行線性回歸方程擬合Probit(θT(g))=(θT(g)- θT(50))/ σ θT(g)。擬合曲線的截距為 [-θT(50)]/ σθT(g),斜率為1/σθT。故θT(50)為負(fù)的截距除以斜率,σθT為斜率的倒數(shù)。
1.3.3 萌發(fā)時(shí)間預(yù)測(cè) 氣象資料由西北農(nóng)林科技大學(xué)旱區(qū)農(nóng)業(yè)研究院提供。利用公式y(tǒng)=y0+asin(2πx)/b+c(式y(tǒng)0為年平均溫度,x為儒略日,a、b和c為待估參數(shù))對(duì)楊凌地區(qū)2008-2018年的月平均溫度的散點(diǎn)圖進(jìn)行回歸擬合,估計(jì)參數(shù)y0,其中y0為年平均溫度。以歷年擬合方程中的y0(年均溫度)對(duì)年份進(jìn)行線性回歸,預(yù)測(cè)2020的平均溫度的y,進(jìn)而估計(jì)萌發(fā)時(shí)間。
1.3.4 數(shù)據(jù)處理 數(shù)據(jù)用R軟件進(jìn)行方差分析及多重比較(Duncan’s法),結(jié)果以“平均值±標(biāo)準(zhǔn)誤”的形式表示。用Sigmaplot 14.0軟件制圖。
2 結(jié)果與分析
2.1 反枝莧種子質(zhì)量評(píng)估
3個(gè)不同來(lái)源的反枝莧種子均呈黑色圓形,直徑1~2 mm,種皮較硬,表面光滑且有光澤。不同生境種子的千粒質(zhì)量、含水量及活力均有顯著差異(表1)。河灘地、二道塬和頭道塬三地的千粒質(zhì)量分別為0.348 5、0.311 7、0.322 5 g,含水量分別為30.29%、25.32%、24.38%,活力分別為87.61%、80.37%、87.83%。

表1 不同生境反枝莧種子質(zhì)量Table 1 Quality of Amaranthus retroflexus seeds collected from three habitats
2.2 反枝莧萌發(fā)過(guò)程
3個(gè)不同來(lái)源的種子萌發(fā)過(guò)程存在明顯差異(圖2),在10.2 ℃時(shí)三地的種子萌發(fā)均極少。在19.2 ℃時(shí),頭道塬反枝莧種子萌發(fā)率即可達(dá)到40%以上,而其他兩地的種子萌發(fā)不足5%。頭道塬反枝莧種子萌發(fā)最適溫度為28.9 ℃,低于二道塬和河灘地的最適溫度36.5 ℃。頭道塬、二道塬、河灘地相應(yīng)的最高萌發(fā)率分別為70.83%、90.66%和34.68%。表明低海拔地區(qū)種子更喜高溫。海拔越低,累積萌發(fā)曲線變化越緩,表明其萌發(fā)速率越低,種子萌發(fā)不整齊(圖3)。
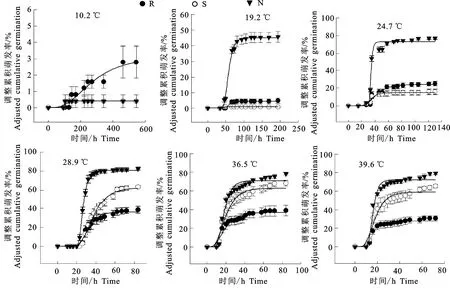
圖2 反枝莧種子不同溫度條件下種子的調(diào)整累積萌發(fā)率Fig.2 Adjusted cumulative germination of Amaranthus retroflexus seeds at different temperatures

圖3 不同溫度條件下頭道塬和二道塬反枝莧種子的萌發(fā)速率直線Fig.3 Germination rate against constant temperatures for Amaranthus retroflexus seed collected from first and second tiered terraceland
2.3 反枝莧時(shí)熱模型參數(shù)評(píng)估
由于本試驗(yàn)河灘地反枝莧種子的最高萌發(fā)率為34.68%,無(wú)法評(píng)估θT(50),故只討論二道塬、頭道塬兩地種子萌發(fā)的θT(50)及標(biāo)準(zhǔn)誤σθT。
頭道塬和二道塬種子的Tb為13.98和 23.14 ℃;θT(50)為398.74和326.79 ℃·h,(由于標(biāo)準(zhǔn)差較大,因此二者差異達(dá)不到顯著水平,表2)反枝莧調(diào)整累積萌發(fā)率隨著積溫的增大而升高(圖 4)。頭道塬種子萌發(fā)所需積溫變幅窄,為334.04 ~463.44 ℃·h;而二道塬的較寬,為177.76~ 475.82 ℃·h。高海拔種源的種子萌發(fā)所需積溫明顯高于低海拔種源的種子(表2和圖4)。頭道塬種子的萌發(fā)積溫標(biāo)準(zhǔn)差為64.70 ℃·h,不足二道塬(149.03 ℃·h)的一半(表2),表明頭道塬種子萌發(fā)時(shí)間更為集中,自然條件下的適應(yīng)性弱于二道塬的種子。
2.4 反枝莧種子萌發(fā)日期預(yù)測(cè)
對(duì)楊凌地區(qū)2008—2018年各月平均溫度的散點(diǎn)圖進(jìn)行回歸擬合,結(jié)果表明,楊凌多年平均溫度(y0)分別為8.9、11.6、13.1、8.8、11.4、12.1、13.2、13.3、12.9、14.3和12.3 ℃,其對(duì)年份的線性回歸方程為y=0.348 6x-689.714 4,R2= 0.424 3。預(yù)測(cè)2020年平均溫度為14.5 ℃。可以得到2020年的日平均溫度數(shù)據(jù)方程:y= 14.5+14.04sin(2πx/405.21+4.84),利用以上公式找到積溫達(dá)到θT(50)及達(dá)到其95%的置信限時(shí)的日期。預(yù)測(cè)萌發(fā)日期以及最早、最晚萌發(fā)日期如表3所示。二道塬的反枝莧種子萌發(fā)率達(dá)50%的日期是6月6日,晚于頭道塬的4月18日,但萌發(fā)時(shí)期變幅為15 d,而頭道塬的則只用 4 d。因此,水熱條件好時(shí),反枝莧種子萌發(fā)時(shí)間區(qū)間更寬,防控更加困難。

表2 反枝莧種子萌發(fā)時(shí)熱模型參數(shù)估計(jì)Table 2 Model parameters estimation of Amaranthus retroflexus seed germination

圖4 頭道塬和二道塬反枝莧種子萌發(fā)的積溫模型曲線Fig.4 Thermal time diagram of Amaranthus retroflexus seeds collected from first and second tiered terraceland
3 討 論
3.1 反枝莧萌發(fā)的熱量需求
不同生境反枝莧種子萌發(fā)的基礎(chǔ)溫度、熱量需求均存在較大差異(表2)。雖然頭道塬、二道塬兩地海拔相差只有60 m,但多年物候表明,二道塬植物開(kāi)花早于頭道塬1~2周。位于低海拔的二道塬反枝莧種子的萌發(fā)基礎(chǔ)溫度比頭道塬的高9.16 ℃。表明在水熱條件相對(duì)較好的條件下,反枝莧種子萌發(fā)啟動(dòng)雖然晚,但萌發(fā)周期長(zhǎng);而處于高海拔的反枝莧種子,萌發(fā)啟動(dòng)溫度低,但萌發(fā)所需積溫較高,種子不會(huì)因短暫適宜的溫度而全部萌發(fā),在溫度積累到較高水平才萌發(fā),這是一種防止高海拔地區(qū)溫度條件對(duì)其幼苗不利、造成損害的萌發(fā)策略。高海拔的反枝莧種子的σθT小,表明高海拔來(lái)源的種子萌發(fā)積溫范圍窄,萌發(fā)相對(duì)整齊,會(huì)在短時(shí)間內(nèi)萌發(fā),為后期的營(yíng)養(yǎng)生長(zhǎng)和生殖生長(zhǎng)留出更長(zhǎng)的時(shí)間。
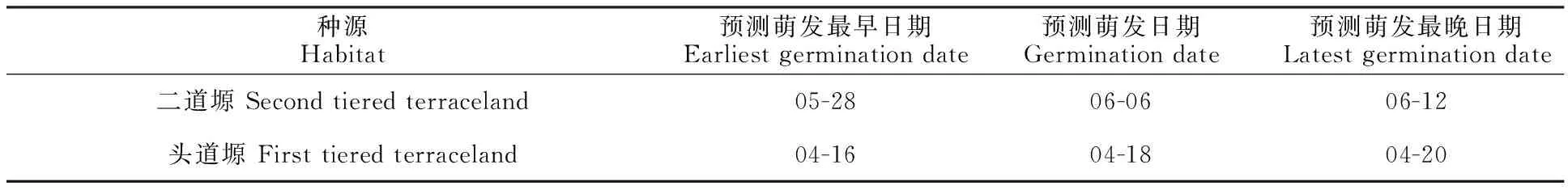
表3 積溫模型預(yù)測(cè)二道塬和頭道塬兩地反枝莧萌發(fā)的日期Table 3 Prediction of germination date of Amaranthus retroflexus seeds based on thermal time model
通過(guò)對(duì)比幾種經(jīng)濟(jì)作物萌發(fā)的基礎(chǔ)溫度與積溫需求發(fā)現(xiàn),反枝莧種子萌發(fā)對(duì)基礎(chǔ)溫度要求較高,而對(duì)積溫要求較低。如大豆種子萌發(fā)的基礎(chǔ)溫度為3.33 ℃,積溫需求為216.7 ℃·d;花生萌發(fā)的基礎(chǔ)溫度為7.5 ℃,積溫需求為87.5 ℃·d[23]。可以看出,反枝莧萌發(fā)基礎(chǔ)溫度高于大豆和花生,所需積溫遠(yuǎn)遠(yuǎn)小于大豆、花生等經(jīng)濟(jì)作物。相較于其他雜草,如野燕麥種子萌發(fā)的基礎(chǔ)溫度為-3.27 ℃,積溫需求為2 146.70 ℃·h等,同樣具有明顯差異。對(duì)于萌發(fā)基礎(chǔ)溫度低、積溫高的種子來(lái)說(shuō),種子萌發(fā)需要較長(zhǎng)的時(shí)間,以保幼苗躲過(guò)不利的溫度條件正常生長(zhǎng)[24]。而反枝莧萌發(fā)基礎(chǔ)溫度較高,可確保幼苗于適宜的溫度條件下生長(zhǎng),故對(duì)積溫沒(méi)有較高要求。
3.2 反枝莧對(duì)未來(lái)氣候變暖的可能響應(yīng)
反枝莧種子的萌發(fā)率隨溫度的升高增大,在10.2 ℃時(shí)幾乎不萌發(fā);19.2 ℃時(shí)高海拔種源的種子最先萌發(fā),24.5 ℃以上時(shí),3種來(lái)源的種子僅需40 h都可達(dá)到其最高萌發(fā)率(圖2)。馬鵬里等[25]對(duì)1961-2014年氣候變暖背景下中國(guó)干旱變化的研究表明,中國(guó)西北地區(qū)干旱程度明顯減輕,因此西北地區(qū)有向暖濕氣候發(fā)展的趨勢(shì)。在這一背景下,反枝莧達(dá)到種子萌發(fā)基礎(chǔ)溫度的日期將會(huì)提前[26],其萌發(fā)也將提前。有研究表明,萌發(fā)早的反枝莧生活史周期較長(zhǎng),春季物候期提前,秋季物候期延遲[26],生育期更長(zhǎng),有更充分的營(yíng)養(yǎng)生長(zhǎng),因而種子的質(zhì)量與數(shù)量將會(huì)更大,全球變暖將會(huì)增加其繁殖優(yōu)勢(shì)。此外,反枝莧作為一種典型C4植物,具有高光合效率、低CO2補(bǔ)償點(diǎn)的光合特性,全球氣候變暖將更有利于其生長(zhǎng),因此會(huì)對(duì)田間生產(chǎn)造成更大影響。本研究中,低海拔暖濕條件下的種子的σθT大,表明低海拔來(lái)源的種子萌發(fā)積溫范圍廣,自然適應(yīng)能力強(qiáng),田間防除更加困難。因此,在全球氣候變暖的條件,反枝莧的防控難度將增加。
3.3 反枝莧防控建議
反枝莧在適宜的條件下可達(dá)到90%的萌發(fā)率(圖2),表明若不及時(shí)防除,將對(duì)田間生產(chǎn)造成極大危害。反枝莧對(duì)多種除草劑都產(chǎn)生了抗性[27],但幼苗期抗性較弱,施用除草劑可以達(dá)到較好的防治效果。因此,定量估計(jì)反枝莧的萌發(fā)日期,可以為適時(shí)化學(xué)除草提供可靠依據(jù),減少用藥,提高效率。由于正在萌發(fā)的種子被土壤覆蓋,噴灑莖葉處理劑對(duì)其進(jìn)行防除沒(méi)有明顯效果。該時(shí)期可采取翻土、施用苗前土壤處理除草劑,例如氯溴隆、利谷隆、溴谷隆等對(duì)其進(jìn)行防治[28]。本研究中,反枝莧萌發(fā)后3 d,幼苗能長(zhǎng)到15 mm左右,5 d長(zhǎng)到25 mm左右并進(jìn)入2葉期,此時(shí)使用450.0 g a.i./hm250%溴苯腈、350.0 g a.i./hm270%麥草畏或600.0 g a.i./hm240%甲基磺草酮/特丁津混劑可有效防治反枝莧[29]。若在4~5葉期防控,可考慮施用孢子濃度107個(gè)/mL的Alternariaalternate,持續(xù)6 h的濕度,導(dǎo)致葉片壞死,植株萎蔫死亡[28]。小根蒜、臭椿、角茴香根等多種植物的提取液都能夠抑制反枝莧種子發(fā)芽,降低幼苗鮮質(zhì)量,并且隨著提取物濃度的提高,抑制效果更加明顯。
4 結(jié) 論
反枝莧種子的萌發(fā)的基礎(chǔ)溫度和最適溫度較高,萌發(fā)熱量需求少,是典型喜溫雜草,萌發(fā)周期短,在全球變暖的大背景下,其萌發(fā)日期將會(huì)提前,萌發(fā)期和生活史周期將會(huì)延長(zhǎng),其生長(zhǎng)繁殖都會(huì)處于優(yōu)勢(shì)地位,防控難度增加。楊凌地區(qū)正常年份的萌發(fā)日期為4月16日至6月12日(因COVID-19疫情影響,原計(jì)劃的野外調(diào)查無(wú)法實(shí)施),此后1~2周內(nèi)實(shí)施化學(xué)除草效果最好。
