類烏齊牦牛RNF216基因的克隆及組織表達分析
2020-12-10 01:50:16柴志欣武志娟王吉坤王嘉博鐘金城信金偉
西南農業(yè)學報 2020年8期
關鍵詞:分析
陳 美,柴志欣,武志娟,王 會,王吉坤,王嘉博,鐘金城*,信金偉
(1.青藏高原動物遺傳資源保護與利用四川省、教育部重點實驗室,四川 成都 610041;2.省部共建青稞和牦牛種質資源與遺傳改良國家重點實驗室,西藏 拉薩 850000)
【研究意義】牦牛(Bosgrunniens)是我國青藏高原的特有畜種,能充分利用高寒高山草原進行動物性生產的優(yōu)勢牛種[1],主要分布在青海、西藏、四川、甘肅、云南、新疆及內蒙古等地區(qū)[2]。西藏自治區(qū)由于特殊的地理特征和生態(tài)環(huán)境形成了許多地方牦牛品種或類群,牦牛遺傳資源十分豐富[3]。牦牛是西藏畜牧業(yè)經濟不可缺少的畜種,牦牛業(yè)在自治區(qū)畜牧業(yè)發(fā)展中占有十分重要的地位。類烏齊牦牛主要分布在西藏自治區(qū)的昌都市類烏齊縣境內海拔4500 m以上的高山草甸草原地區(qū),屬以產肉為主肉乳兼用型牦牛。中心產區(qū)為類烏齊鎮(zhèn)、卡瑪多鄉(xiāng)、長毛嶺鄉(xiāng)和吉多鄉(xiāng)等鄉(xiāng)鎮(zhèn),是經長期自然選擇而形成的能適應當?shù)厣鷳B(tài)環(huán)境的優(yōu)良牦牛遺傳資源,類烏齊牦牛肉也因肉質鮮美,天然無公害,具有高蛋白質和礦物質含量,低脂肪、低能量、氨基酸含量豐富等特點[4-5],深受當?shù)剞r牧民的喜愛,也成為其增產增收的主要來源之一。環(huán)指蛋白216抗體(ring finger protein 216,RNF216),也稱含三聯(lián)域蛋白3(Triad3A)、泛素偶聯(lián)酶7-相互作用蛋白1或鋅指蛋白抑制NF-kappa-B(ZIN),是E3泛素連接酶的RING家族的一員,在其N末端包含一個鋅指結構域,C末端包含一個環(huán)指結構域。RNF216是含有RING結構域的E3泛素連接酶,在巨噬細胞、乳腺癌細胞、性腺發(fā)育以及肥胖等生物過程中均發(fā)揮一定作用[6-7]。【前人研究進展】Santens等[8]在人常染色體隱性遺傳疾病中發(fā)現(xiàn)RNF216突變可能導致Gordon Holmes綜合征,即一種以性腺功能低下和小腦共濟失調為特征的疾病,以及常染色體隱性遺傳性亨廷頓樣疾病;黃施倩等[7]采用野生型(WT)和RNF216基因缺陷型(RNF216-/-)小鼠,使用EO771乳腺癌細胞建立乳腺癌移植瘤模型,觀察小鼠乳腺癌細胞的成瘤情況,確定RNF216可通過調控Th1型免疫應答參與腫瘤的發(fā)生;Xu等[9]在人的自噬細胞中發(fā)現(xiàn)RNF216有強烈抑制巨噬細胞自噬的作用,RNF216與自噬作用中的關鍵調節(jié)因子BECN1相互作用,使BECN1泛素化,從而導致BECN1降解,且RNF216通過與三聯(lián)體(兩個RING和DRIL)結構域直接相互作用參與BECN1賴氨酸48的泛素化等過程;Wang等[10]發(fā)現(xiàn)RNF216在人結腸直腸癌(CRC)組織和細胞中有調節(jié)功能,并與CRC的進展相關。Goyenechea等[11]選擇兩組肥胖受試者,通過qRT-PCR在3個時間點(第0周,第8周和第32周)評估外周血單核細胞(PBMC)中RIPK3和RNF216的身體組成和mRNA水平,發(fā)現(xiàn)肥胖受試者在接受低熱量飲食(LCD)的過程中,體重會明顯減輕,且這些基因的mRNA水平可用于預測肥胖癥治療結果的營養(yǎng)基因組生物標志物。常染色體隱性遺傳疾病的發(fā)生,涉及RNF216,OTUD4和STUB1的泛素化及基因的編碼突變,如Jonatha等[12]對CHIP-/-小鼠的運動缺陷進行了表型分析,在STUB1突變患者中觀察到性腺機能減退,說明RNF216突變患者也出現(xiàn)相同現(xiàn)象,且通過透射電子顯微鏡對骨骼肌進行觀察,發(fā)現(xiàn)CHIP -/-小鼠具有形態(tài)學變化,其與股四頭肌和肌肉痙攣的肌漿網(wǎng)室增加一致腓腸肌(有毒低聚物和管狀聚集體)有關,說明CHIP與常見的肌肉和心臟疾病具有相關性。【本研究切入點】以上研究顯示,RNF216除在人的疾病中發(fā)揮作用外,其在肥胖或者肌肉相關疾病等方面也發(fā)揮作用。目前,針對RNF216基因的相關研究主要集中于人和小鼠,【擬解決的關鍵問題】本研究選取類烏齊牦牛通過克隆獲得RNF216基因的cDNA序列,并利用熒光定量PCR方法檢測該基因在不同組織器官中的表達情況,為牦牛的肌肉發(fā)育分子調控機制提供重要理論依據(jù),以期為優(yōu)化牦牛選育體系提供數(shù)據(jù)支撐。
1 材料與方法
1.1 材料
1.1.1 試驗樣本采集 于西藏自治區(qū)昌都市類烏齊縣選取3頭健康的0.5歲齡類烏齊牦牛,采集其肝臟、肺臟、心臟、脾臟、臀脂和臀肌等組織,DEPC水沖洗干凈后,錫箔紙包裝迅速置于液氮中保存?zhèn)溆谩?/p>
1.1.2 實驗試劑 YBR Premix ExTaqTMⅡ、PrimeScriptTMRtreagent Kit with gDNA Eraser(Perfect Real Time)RNA 反轉錄試劑盒、pGEM-T Vector cloning kit、DH5α感受態(tài)細胞、2000 bp Marker和TB Green Premix ExTaqII (TaKaRa公司);組織RNA提取試劑TRIzol(美國 Invitrogen公司);DNA 純化回收試劑盒(北京 天根生化科技有限公司)。
1.2 方法
1.2.1 引物設計與合成 根據(jù)GenBank中公布的黃牛RNF216基因mRNA序列(NM_001193258.1),利用Primer Premier 5.0軟件設計PCR引物(表1);將克隆獲得的類烏齊牦牛CDS區(qū)序列用于設計熒光定量PCR引物,引物由成都擎科梓熙生物技術有限公司合成。
1.2.2 組織總RNA提取及cDNA鏈的合成 采用Trizol法提取組織總RNA,用1 %瓊脂糖凝膠電泳檢測所提RNA樣品的完整性,分光光度計測定 RNA濃度及D260nm/D280nm值。按照反轉錄試劑盒說明書將其反轉錄成cDNA,置于-20 ℃保存。
1.2.3RNF216基因克隆與測序 PCR擴增體系(25 μl):2×TaqGreen PCRMasterMix 12.5 μl,上、下游引物各1 μl, cDNA模板1 μl,ddH2O 9.5 μl。PCR擴增程序:95 ℃預變性5 min,95 ℃變性30 s,54 ℃退火30 s,72 ℃延伸90 s,35個循環(huán),72 ℃再延伸5 min,4 ℃保存。將PCR擴增產物用1 %瓊脂糖凝膠電泳檢測其條帶清晰度及完整性,利用DNA純化試劑盒進行純化回收,將純化回收后的目的片段與pGEM-T載體在16 ℃的條件下連接過夜,后轉化到 DH5α感受態(tài)細胞(大腸桿菌)中冰浴30 min,取出于超凈工作臺中,加入900 μl不含Amp+的無菌LB培養(yǎng)液,在37 ℃、150 r/min的搖床中培養(yǎng)45 min,復蘇后將其涂布于含有Amp+的LB培養(yǎng)基上,37 ℃倒置培養(yǎng)10~14 h,挑取合適的克隆菌置于含Amp+的LB培養(yǎng)液中37 ℃、180~200 r/min震蕩培養(yǎng)10~12 h,經菌液PCR鑒定后送成都擎科梓熙生物技術有限公司測序。

表1 引物序列
1.2.4 熒光定量PCR檢測 將克隆得到的RNF216基因目的片段進行拼接,用于設計實時熒光定量特異性引物。以GAPDH為內參基因,cDNA為模板,進行牦牛組織的熒光定量PCR,分析RNF216基因在心臟、肝臟、脾臟、肺臟、臀脂和臀肌等組織的相對表達量。反應體系(10 μl):SYBR Premix ExTaqTM(2×)5 μl,上、下游引物各0.4 μl,cDNA模板1 μl,ddH2O 3.2 μl。反應程序:95 ℃預變性30 s,95 ℃變性5 s,55 ℃退火30 s,共39個循環(huán)。用 2-ΔΔCt法計算各個樣本的相對表達量,利用SPSS 19.0軟件進行顯著性分析。
1.2.5RNF216基因序列的生物信息學分析 利用NCBI中ORF Finder程序分析牦牛RNF216基因開放閱讀框;ExPASy中ProtParam預測其蛋白分子質量、氨基酸組成、親水性等理化性質;NetPhos預測蛋白磷酸化位點;TMHMM、PSORT II和SignaLP分別預測其蛋白的跨膜結構、信號肽和亞細胞定位;SOPMA及SWISS MODEL預測蛋白的二級、三級結構;Clustal X和MEGA軟件分析基因序列一致性并構建系統(tǒng)進化樹。
2 結果與分析
2.1 牦牛RNF216基因PCR擴增結果
按照克隆引物序列,以牦牛肝臟組織cDNA為模板進行PCR擴增,PCR產物經1 %瓊脂糖凝膠電泳檢測,其擴增條帶清晰單一,分別為500、1262和1242 bp,與預期擴增片段大小吻合(圖1)。
利用NCBI中ORF Finder在線程序對克隆得到的牦牛RNF216基因序列進行開放性閱讀框分析,獲得RNF216基因CDS區(qū)序列長為2223 bp,共編碼740個氨基酸,起始密碼子為ATG,終止密碼子為TGA(圖2)。
2.2 牦牛RNF216基因的理化性質分析
利用ExPASy在線軟件分析類烏齊牦牛 RNF216 蛋白質序列的理化性質。結果顯示,共編碼740個氨基酸,其中Glu含量最多(10.8 %),不含Pyl和Sec。負電荷殘基總數(shù)(Asp + Glu)134個,正電殘基總數(shù)(Arg + Lys)82個。該蛋白分子式為C3710H5721N1019O1172S45,其分子量為84 785.06 D,原子總數(shù)11 667,理論等電點PI為4.79,親水性平均值(GRAV)為-0.722,脂肪指數(shù)為68.19,不穩(wěn)定指數(shù)(II)為60.19,說明該蛋白為不穩(wěn)定親水酸性蛋白。

圖2 牦牛RNF216基因編碼的蛋白序列Fig.2 Protein sequence encoded by the yak RNF216 gene
2.3 牦牛RNF216蛋白的親水性/疏水性及亞細胞定位預測
利用Expasy中Prot Scale程序分析RNF216蛋白疏水性/親水性,發(fā)現(xiàn)信號蛋白的最小值為-3.300,位點位于432位,其親水性最強;最大值為1.967,位點位于322位,其疏水性最強(圖3),說明此蛋白為親水性蛋白。利用PSORTII Prediction 在線軟件進行牦牛RNF216蛋白的亞細胞定位,發(fā)現(xiàn)該蛋白在細胞核中的分布最多,高達91.3 %;在囊泡和細胞骨架內分布較少,均為4.3 %。
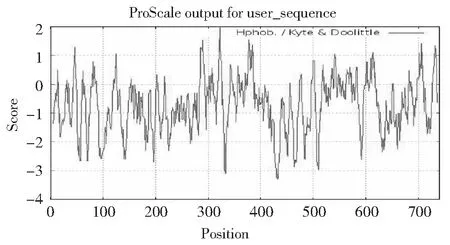
正值表示疏水;負值表示親水 Positive values indicate hydrophobicity; negative values indicate hydrophilic圖3 牦牛RNF216蛋白的疏水性/親水性預測Fig.3 Hydrophobic/hydrophilic prediction of RNF216 protein in yak
2.4 牦牛RNF216基因編碼蛋白跨膜區(qū)與信號肽預測
采用TMHMM在線軟件分析RNF216基因編碼蛋白跨膜區(qū),發(fā)現(xiàn)類烏齊牦牛RNF216蛋白不存在跨膜區(qū)域,不屬于膜上受體或無法定位于細胞膜上,說明該蛋白質非跨膜蛋白。
SignalP在線分析發(fā)現(xiàn)RNF216蛋白無信號肽區(qū)域,說明RNF216屬非分泌蛋白。
2.5 牦牛RNF216蛋白磷酸化分析
運用Net Phos2.0軟件對RNF216蛋白磷酸化位點進行預測,發(fā)現(xiàn)牦牛RNF216基因編碼的氨基酸中共發(fā)現(xiàn)24個絲氨酸(Ser)、8個蘇氨酸(Thr)和9個酪氨酸(Tyr)磷酸化位點(圖4)。
2.6 牦牛RNF216二級結構和三級結構預測
采用SOPMA在線軟件對牦牛RNF216蛋白二級結構進行預測,RNF216蛋白主要結構為無規(guī)卷曲和α-螺旋,分別占總鏈48.11 %和41.32 %;延伸鏈和β-轉角,分別占總鏈的7.74 %和2.83 %(圖5)。SWISS-MODEL軟件預測RNF216蛋白三級結構,發(fā)現(xiàn)該蛋白是基于ARIH1原子結構5tte.1的A鏈進行模建,兩者序列一致性可達25.13 %。

圖4 牦牛RNF216蛋白磷酸化分析Fig.4 Phosphorylation analysis of RNF216 protein in yak

圖5 牦牛RNF216蛋白二級結構預測Fig.5 Prediction of secondary structure of RNF216 protein in yak
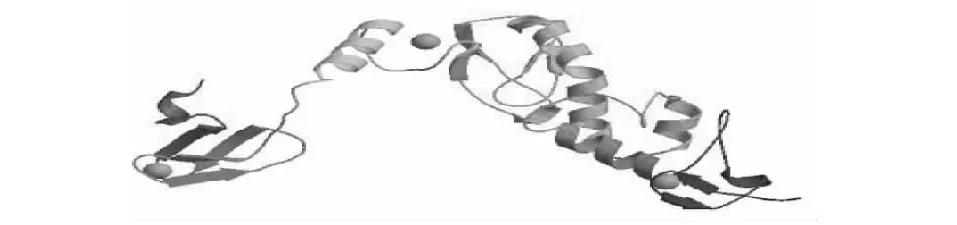
圖6 牦牛RNF216蛋白三級結構預測Fig.6 Prediction of tertiary structure of RNF216 protein in yak
2.7 物種一致性比較與系統(tǒng)進化樹分析
利用Lasergene軟件中MegAlign程序對類烏齊牦牛RNF216基因CDS區(qū)序列和其他物種該基因的CDS進行比對,并用MEGA.5軟件構建系統(tǒng)進化樹。由表3可知,類烏齊牦牛RNF216基因序列與GenBank中黃牛、白海豚、豬的序列一致性較高,分別為93.5 %、91.3 %和90.3 %;與人和鼠類的一致性較低。依據(jù)牦牛RNF216基因構建進化樹,發(fā)現(xiàn)牦牛與黃牛的遺傳距離最近,與鼠類親緣關系最遠,符合物種進化規(guī)律(圖7)。
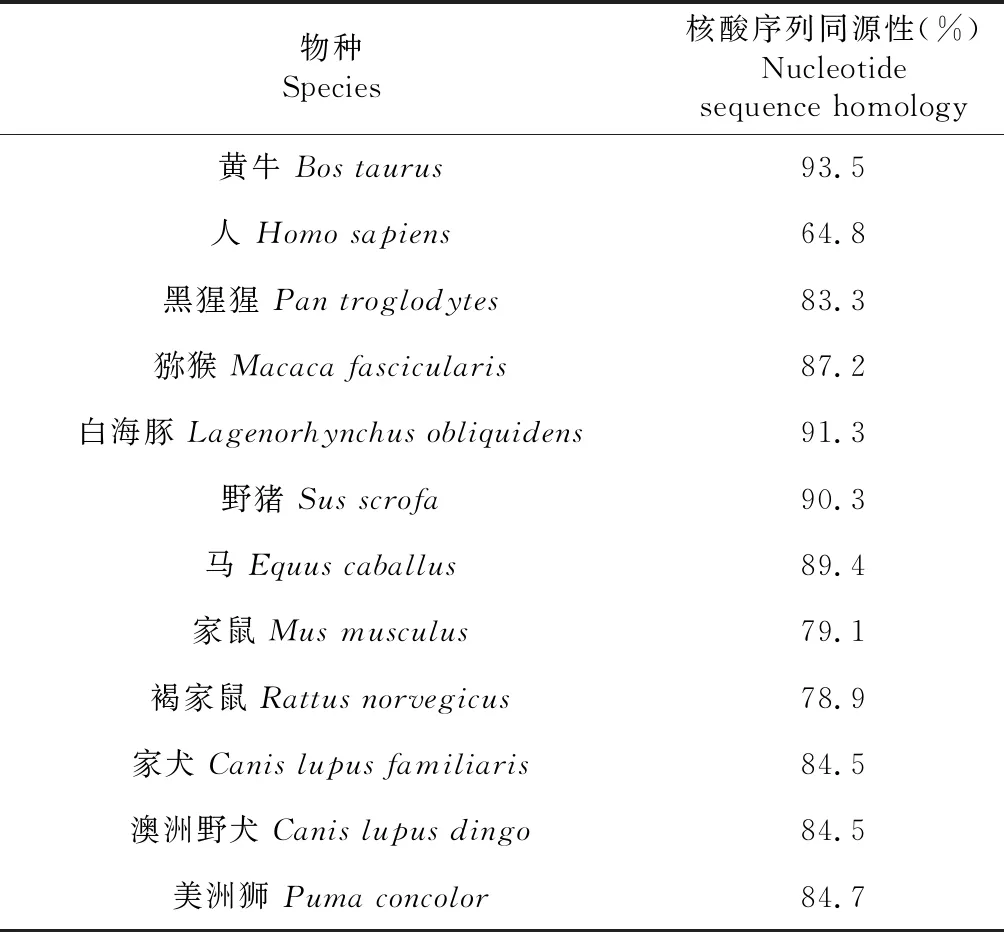
表2 牦牛與12個物種RNF216基因的序列同源性比對

圖7 牦牛RF216基因的系統(tǒng)進化樹Fig.7 Phylogenetic tree of RF216 gene in yak
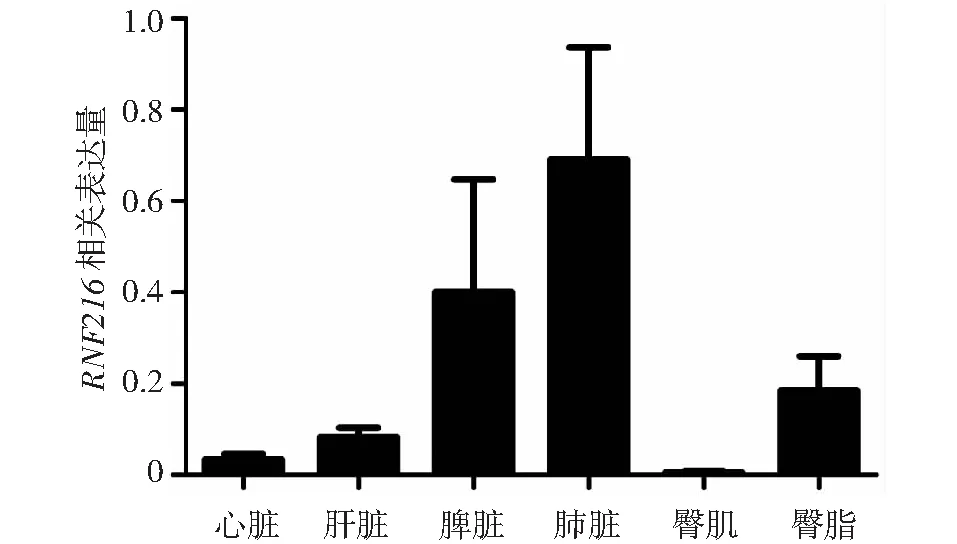
圖8 牦牛RNF216基因的mRNA組織表達分析Fig.8 mRNA expression analysis of RNF216 gene in yak
2.8 牦牛RNF216基因不同組織差異表達分析
以GAPDH基因為內參基因,利用實時熒光定量PCR對類烏齊牦牛的心臟、肝臟、脾臟、肺臟、臀肌和臀脂等組織進行差異分析,發(fā)現(xiàn)牦牛RNF216基因在以上組織中均有表達 (圖8),其中在肺臟中的表達水平最高,其次是脾臟,在臀肌的表達水平最低。
3 討 論
中國是世界上牦牛遺傳資源最多的國家,雖牦牛生長在高寒地區(qū),肌纖維較粗,肉質嫩度中,較其它品種牛肉屬于中等偏韌[13-14],但牦牛肉是西藏自治區(qū)特有的肉類資源,富含豐富的蛋白質和氨基酸,從而受到廣大消費者青睞[15]。因此,隨著人們生活水平不斷提高,提高牦牛肉品質及產量成為研究的熱點之一,近年來,國內眾多學者在提高牦牛生產性能研究的基礎上開始對其肉質性狀進行研究和開發(fā)[16]。通過篩選影響牦牛肉品質及產肉量的關鍵基因,并對其進行定量表達分析成為目前研究基礎。
RNF216是一種RBR型E3泛素蛋白連接酶,其與TLR的Toll /白細胞介素-1受體結構域相互作用并促進其蛋白水解降解[17-18]。研究表明,通過RNF216上調可抑制TLR4表達,而TLR4的異常激活在肥胖誘導的炎癥中具有關鍵作用,且與高胰島素血癥、高甘油三酯血癥和心血管疾病等疾病相關[19],說明RNF216可能與脂肪和糖代謝相關。RNF216在自噬中也發(fā)揮重要作用,自噬是細胞穩(wěn)態(tài)的基礎[10]。它還控制受體相互作用蛋白1(RIP1)的泛素化和蛋白酶體降解,RIP1是一種絲氨酸/蘇氨酸蛋白激酶,在Hsp90結合被破壞后,該蛋白激酶與腫瘤壞死因子受體1(TNF-R1)誘導的NF-kappa B活化密切相關[20-21]。此外,RNF216與人類免疫缺陷病毒1型(HIV-1)病毒粒子感染因子(Vif)蛋白相互作用,而Vif蛋白對初級人類CD4T淋巴細胞和巨噬細胞的生產性感染至關重要[22]。
本研究成功克隆了類烏齊牦牛RNF216基因的CDS區(qū)序列,對亞細胞定位進行分析,發(fā)現(xiàn)RNF216基因編碼蛋白主要存在細胞核中,而細胞核是遺傳物質儲存和復制的場所及細胞遺傳性和細胞代謝活動的控制中心,說明RNF216蛋白可能參與細胞內的代謝調控,且在維持細胞脂質穩(wěn)態(tài)等方面發(fā)揮一定作用[23]。對牦牛RNF216蛋白進行磷酸化分析,發(fā)現(xiàn)存在24個絲氨酸、8個蘇氨酸和9個酪氨酸磷酸化位點,其中絲氨酸位點的數(shù)量最多,而絲氨酸在脂肪和脂肪酸新陳代謝及肌肉生長過程中發(fā)揮作用,并在肌肉組織和包圍神經細胞的鞘的合成等過程均發(fā)揮作用,推測RNF216基因可能在類烏齊牦牛的脂肪代謝和肌肉生長過程中發(fā)揮一定作用,接下來我們將進一步開展該基因功能的相關研究。因RNF216基因在核苷酸和氨基酸序列中與哺乳動物的一致性較高,同時類烏齊牦牛RNF216與黃牛的遺傳距離最近,并與鼠類遺傳距離較遠,說明該基因在進化過程中保守性強。
運用qPCR 技術檢測牦牛心臟、肝臟、脾臟、肺臟、臀肌和臀脂等組織中RNF216基因的表達情況,發(fā)現(xiàn)該基因在肺臟中的表達水平最高,其次是脾臟,然后是臀脂,而在臀肌中的表達水平最低,根據(jù)以上研究結果可知RNF216基因主要在牦牛肺臟組織中作用,而肺作為機體重要的呼吸器官,牦牛主要生活在海波3500 m以上的高寒草場,具有耐低氧、寒冷、強紫外線等優(yōu)良特性,推測該基因在牦牛低氧適應性的生理特性中發(fā)揮一定調控作用。牛肉感官品質的指標包括牛肉色澤、肌肉內脂肪含量及肉質柔韌性等[24],而肌內脂肪(IMF)含量是影響肉質的主要因素[25],牛肉肌肉內的脂肪含量越高,肉質越是滑嫩,品質也越好。而牦牛肌肉蛋白質含量高,而脂肪含量非常低。本研究表明,牦牛RNF216基因在臀脂中的表達水平也較高,且在臀肌中也有分布,推測該基因在牦牛的肌肉脂肪發(fā)育過程中也發(fā)揮一定的作用。
4 結 論
本研究成功克隆了牦牛RNF216基因的CDS序列,編碼 740 個氨基酸殘基。該基因編碼蛋白不存在信號肽和跨膜結構,為不穩(wěn)定親水酸性蛋白,主要分布在細胞核中。牦牛RNF216蛋白存在 24個絲氨酸、8個蘇氨酸和9個酪氨酸磷酸化位點。其與黃牛的遺傳距離是最近,與鼠類的遺傳距離最遠,RNF216基因在肺臟組織中的表達水平最高。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
