全基因組預測哈茨木霉的效應子
2020-12-10 10:45:14陳欣瑜董章勇
西南農業學報 2020年9期
羅 梅,陳欣瑜,2,董章勇,2*
(1.仲愷農業工程學院 農業與生物學院,廣東 廣州 510225;2.仲愷農業工程學院植物健康創新研究院,廣東 廣州 510225)
【研究意義】哈茨木霉(Trichodermaharzianum)是木霉菌的一種,屬半知菌類(Fungi imperfecti),絲孢綱(Hyphomycetes),叢梗孢目,叢梗孢科,是一種目前廣泛應用的重要生防菌。該屬廣泛分布于全球,在土壤及有機質上均可發現[1]。哈茨木霉報道能夠與長孺孢屬(Helmisporiumspp.)、核盤菌屬(Sclerotiniaspp.)、輪枝孢屬(Verticilliumspp.)、黑星菌屬(Venturiaspp.)、蜜環菌屬(Armillariaspp.)、鐮刀菌(Fusariumspp.)、腐霉菌(Pythiumspp.)、疫霉菌(Phytophthoraspp.)、絲核菌(Rhizoctoniaspp.)、炭疽菌(Colletotrichumspp.)等多種植物病原菌競爭生長空間和營養物質[2-4],且能分泌產生一些抑菌物質,具有重寄生作用,并可以誘導植物產生抗性[5-7]。因此,其菌劑被廣泛應用在植物病害的防治上[6-7]。以色列的哈茨木霉T39和美國T22 菌株等均已經登記使用在多種植物病害的防治上。【前人研究進展】效應子(effector)也稱效應因子或效應蛋白,廣義上是指能夠選擇性地結合其他目標蛋白及調節其生物活性的一類小分子蛋白。隨著對效應子的研究,有學者凝煉出其具體的分析指標:①含有N-端信號肽;②無跨膜結構域;③定位于細胞質;④序列具有特異性;⑤富含半胱氨酸;⑥重復元件多;⑦氨基酸殘基數量大約在50~300 氨基酸[8-9]。哈茨木霉菌株的基因組測序的完成為高通量篩選效應子提供前提。許僖等[10]根據哈茨木霉T677菌株的基因組預測了其分泌蛋白,但并未進行效應子的分析。【本研究切入點】本研究在已公布的哈茨木霉CBS 226.95菌株全基因組信息的基礎上,根據效應子的典型特征,利用生物信息學軟件對其候選效應子進行了預測。【擬解決的關鍵問題】研究結果為哈茨木霉效應子的進一步篩選與驗證提供了重要的理論依據,并為探究效應子在生防菌與病原菌互作過程中的作用奠定基礎。
1 材料與方法
1.1 基因組信息
哈茨木霉CBS 226.95菌株的蛋白序列從NCBI上下載,具體網址為:ftp://ftp.ncbi.nlm.nih.gov/genomes/all/GCF/003/025/095/GCF_003025095.1_Triha_v1.0/GCF_003025095.1_Triha_v1.0_protein.faa.gz。該基因組共包含了14 065個蛋白序列(INSDC: MBGI00000000.1)。
1.2 分泌組蛋白預測
將14 065個蛋白序列先經過SignalP v4.1分析;隨后將有信號肽的蛋白進一步用TMHMM v2.0進行分析,選取跨膜結構域小于2 的蛋白;進一步用TargetP v1.1進行分析,篩選定位在胞外的蛋白;然后用ProtComp v9.0進行分析亞細胞定位。
1.3 分泌組蛋白序列分析
將1.2分析結果進行序列長度統計分析。
1.4 分泌組蛋白的半胱氨酸含量和multiple tandem repeats分析
將1.2分析結果的蛋白序列進行半胱氨酸數量及multiple tandem repeats分析。半胱氨酸數量采用perl工具進行計算。將具有高半胱氨酸的 (≥6) 的蛋白序列進行multiple tandem repeats分析。multiple tandem repeats采用T-Reks (http://bioinfo.montp.cnrs.fr/?r= t-reks/)進行預測。
1.5 候選致病相關效應因子的確定
一般的效應子都是小分子蛋白,因此,將1.4獲得的蛋白序列去除大于300個氨基酸的蛋白序列,剩余的為候選效應子。將獲得的蛋白序列和PHI數據庫(Pathogen-Host Interaction database,http://www.Phi-bas e.org/downloadLink.htm)進行比對分析,分析其可能的功能。
2 結果與分析
2.1 分泌組蛋白預測
將14 065個哈茨木霉蛋白序列用SignalP v4.1進行分析,共獲得1288個蛋白具有信號肽;隨后進一步用TMHMM v2.0進行跨膜結構分析,跨膜結構域小于2的共有1186個蛋白;將這1186個蛋白用TargetP v1.1進行定位分析,共得到1138個蛋白在胞外表達;再進一步用ProtComp v9.0進行分析,得到709個蛋白。這709個蛋白為哈茨木霉CBS226.95菌株的分泌蛋白,占總蛋白的5.04 %。
2.2 分泌組蛋白序列分析
將709個蛋白氨基酸殘基序列特征進行分析,獲得序列長度統計數據(perl程序)。小于200個氨基酸的有199個,占28 %;201~400個氨基酸的有275個序列,占39 %;401~600個氨基酸的有149個序列,占21 %;601~800個氨基酸的有57個序列,占8 %;801~1000個氨基酸的有23個序列,占3 %;大于1000個氨基酸殘基的有6個序列,占1 %;沒有大于2000個氨基酸殘基的序列(圖1)。
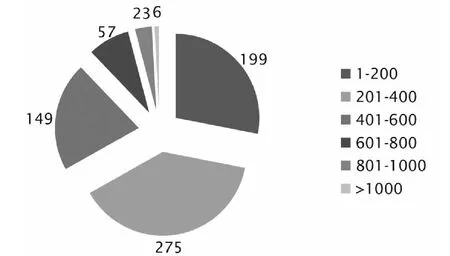
圖1 709個哈茨木霉分泌蛋白的長度統計分析Fig.1 Sequence length statistics of 709 Trichoderma harzianum secretome genes
2.3 分泌組蛋白的cysteine含量和multiple tandem repeats分析
將709個蛋白序列進行cysteine含量及multiple tandem repeats分析。Cysteine含量采用perl工具進行計算。得到396個具有高cysteine含量的 (≥6) 的蛋白。其中6個半胱氨酸的有93個序列,占23.5 %,其次為8個半胱氨酸,有72個序列,占18.2 %;再次為7、10個,分別為45和47個,占11.4 %和11.9 %;其中1個序列(XP_024772708.1)擁有98個半胱氨酸位點(圖2)。
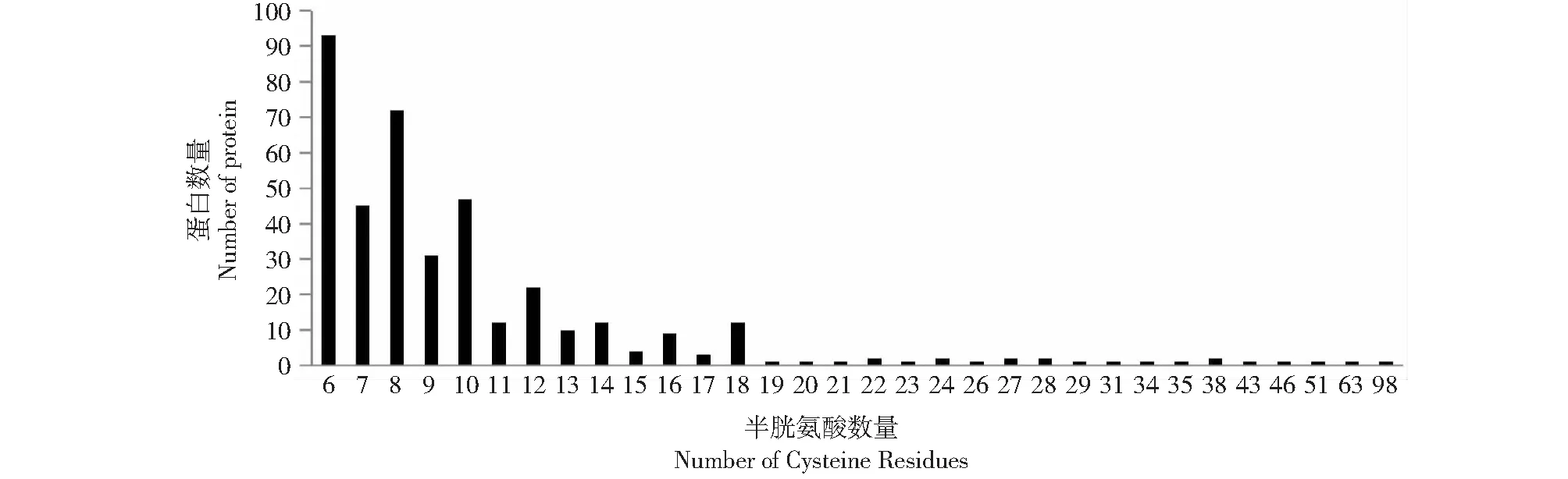
圖2 396個哈茨木霉分泌蛋白的半胱氨酸殘基數量分布Fig.2 Number of cysteine residues of the 396 Trichoderma harzianum secretome genes
將396個具有高cysteine含量的 (≥6) 的蛋白序列進行multiple tandem repeats分析。multiple tandem repeats采用T-Reks (http://bioinfo.montp.cnrs.fr/?r=t-reks/)進行預測。得到86個蛋白具有multiple tandem repeats (≥9)。
2.4 候選致病相關效應因子的確定
一般效應因子都是小分子蛋白,因此,將上述獲得的86個基因序列進行長度篩選,去除大于300個氨基酸的蛋白序列。剩余24個基因序列,被認為是候選致病相關效應因子(表1)。其中有3個是碳水化合物結合模塊家族蛋白(carbohydrate-binding module family protein),其余21個為假定蛋白。在PHI數據庫比對結果顯示,其中9個蛋白可能跟真菌的毒力相關(reduced virulence),3個為效應子(effector),2個蛋白是致死蛋白(lethal),6個為非致病相關(Unaffected pathogenicity),1個蛋白為功能未知,1個蛋白為混合型(可能為非致病相關或引起毒力減弱)。

表1 哈茨木霉候選效應因子

續表1 Continued table 1
3 結論與討論
分泌的胞外蛋白除了作為效應子參與侵染寄主過程外,還可以作為毒力因子或者毒素改變寄主細胞的結構或者功能,或者作為無毒因子或者激發子引發寄主防衛反應[11-12]。本研究通過對哈茨木霉CBS 226.95菌株的14 065個蛋白進行分析,共分析得到709個分泌蛋白,占總蛋白的5.04 %。而許僖等[10]根據哈茨木霉T677菌株基因組的11 498個蛋白中共分析得到分泌蛋白503個,占總蛋白的4.37 %。不同的植物真菌所含分泌蛋白占的比列不同,多數半活體營養型和死體營養型真菌編碼的分泌蛋白占總基因的比例高于活體營養型真菌[13]。本研究分析到的分泌蛋白的比例比許僖等[10]分析的略高,可能2個菌株來源不一樣的差異引起的。
本研究基于哈茨木霉CBS 226.95菌株基因組共篩選獲得24個效應子,其中有3個是碳水化合物結合模塊家族蛋白(carbohydrate-binding module family protein,CBM),其余21個為假定蛋白。雖然通過PHI數據庫對其功能進行了初步的注釋,但是相應蛋白在哈茨木霉的具體功能還需要進一步進行確認。3個碳水化合物結合模塊家族蛋白中有1個是CBM50 亞家族,2個是CBM1亞家族。CBM作為碳水化合物酶類的一大類,在真菌的作用機制上發揮了重要作用。其中CBM50亞家族屬于CBM的一個亞家族,通常結合糖基水解酶家族(glycoside hydrolases,GH)發揮降解幾丁質或者肽聚糖的作用(http://www.cazy.org/CBM50.html)。柳少燕等[14]分析表明拮抗菌中的CBM50相比病原菌來說可能發生了顯著的擴增,且有部分拮抗菌中的CBM50蛋白單獨聚為一支。本研究所分析獲得的CBM50亞家族基因具有1個LysM結構域。LysM結構域最早是在芽孢桿菌噬菌體溶菌酶中被發現其通過降解N-乙酰胞壁酸和N-乙酰葡糖胺之間的糖苷鍵來降解細胞壁[15]。LysM結構域偏好出現在蛋白質的N端和C端,在中心位置出現較少[16]。本研究所分析的CBM50蛋白的LysM結構域出現在C端。由于拮抗菌與病原菌的細胞組分相似,拮抗菌需要解決如何識別“自我”與“非我”并避免自身受到影響的關鍵問題。LysM結構域作為一類古老而又普遍存在的結構域,對該類結構域的研究有助于揭示拮抗真菌、病原真菌及植物防御之間的復雜聯系。Chen等[25]報道效應子LysM幫助逃避昆蟲的免疫防御而致病昆蟲。
隨著基因組學以及生物信息學的快速發展,大數據分析成為現代生物學研究的重要研究手段之一[10, 17-19]。通過高通量的方法進行效應子的篩選,雖然可能不完全,但可以大大的縮小研究范圍,找到研究的切入口。而本研究篩選到的潛在的效應子在哈茨木霉的拮抗作用以及在植物環境中的作用還需要進行進一步的功能研究才能得以驗證。
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
電子制作(2018年18期)2018-11-14 01:48:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
山東工業技術(2016年15期)2016-12-01 05:31:22
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
