里氏木霉與黏綠木霉中非核糖體肽合成酶的生物信息學(xué)分析
2020-12-09 05:31:47祝友朋韓長志
江蘇農(nóng)業(yè)學(xué)報 2020年5期
祝友朋 韓長志

摘要:為了更好地解析里氏木霉和黏綠木霉在次生代謝產(chǎn)物關(guān)鍵基因方面的差異,以非核糖體肽合成酶(NRPS)為例,通過antiSMASH預(yù)測網(wǎng)站對上述2種木霉開展次生代謝產(chǎn)物基因找尋。結(jié)果表明,里氏木霉中含有87個NRPS蛋白,而黏綠木霉中含有214個NRPS蛋白,后者明顯高于前者;同時,利用ProtComp v9.0、PHD等生物信息學(xué)預(yù)測網(wǎng)站對上述木霉中NRPS蛋白進行亞細胞定位、理化性質(zhì)以及二級結(jié)構(gòu)等特征分析,明確2種木霉中NRPS蛋白具有以下共同特征:理論等電點多集中在5.01至7.00之間,不穩(wěn)定性系數(shù)多集中在30.01至50.00之間,高達75%以上的蛋白質(zhì)屬于親水性蛋白質(zhì),而二級結(jié)構(gòu)組成特征則以α螺旋為主,轉(zhuǎn)運肽情況尚未明確,保守結(jié)構(gòu)域較多,亞細胞定位多集中在線粒體等。
關(guān)鍵詞:里氏木霉;黏綠木霉;非核糖體肽合成酶(NRPS);理化性質(zhì);生物信息學(xué)分析
中圖分類號:S435.1文獻標識碼:A文章編號:1000-4440(2020)05-1119-07
Abstract:In order to better analyze the differences of the key genes of secondary metabolites between Trichoderma reesei and Trichoderma virens, the secondary metabolite genes of T. reesei and T. virens were searched through prediction website of antiSMASH, taking non-ribosomal peptide synthetase (NRPS) as example. The results showed that there were 87 NRPS proteins in T. reesei and 214 NRPS proteins in T. virens, the number of NPRS proteins in T. virens was significantly higher than that in T. reesei. Meanwhile, ProtComp v9.0, PHD and other websites for bioinformatics prediction were used to analyze the characteristics of NRPS proteins in T. reesei and T. virens, including subcellular localization, physicochemical properties and secondary structure, etc. The following common characteristics of NRPS proteins in T. reesei and T. virens are clear: the theoretical isoelectric points are mostly concentrated between 5.01 and 7.00, the instability indices are mostly concentrated between 30.01 and 50.00, more than 75% proteins are hydrophilic, while the compositional characteristics of secondary structures are dominated by α-helix. Besides, the information of transit peptides is not clear, there are many conserved domains and the subcellular localizations are mostly concentrated in mitochondria and so on.
Key words:Trichoderma reesei;Trichoderma virens;non-ribosomal peptide synthetase(NRPS);physical and chemical properties;bioinformatics analysis
里氏木霉(Trichoderma reesei)與黏綠木霉(Trichoderma virens)均屬于木霉屬真菌中的重要種類,分別在工業(yè)、農(nóng)業(yè)生產(chǎn)方面發(fā)揮著重要作用[1-2]。里氏木霉QM6a作為重要的工業(yè)菌株,在實現(xiàn)植物組織降解及利用方面發(fā)揮著重要作用,黏綠木霉GV29-8作為木霉屬中重要的生防菌株之一,其在抑制植物病原菌生長發(fā)育方面具有重要的作用。2003年,國外學(xué)者首次通過轉(zhuǎn)錄組學(xué)研究對里氏木霉野生型QM6a開展了全局性分析[3],之后里氏木霉QM6a以及黏綠木霉GV29-8的全基因組序列得以釋放[2,4],為開展深入分析提供了重要的數(shù)據(jù)支撐。前人研究發(fā)現(xiàn)黏綠木霉對立枯絲核菌(Rhizoctonia solani)[5]、終極腐霉(Pythium ultimum)[6]等植物病原菌均有較好的抑制作用。針對里氏木霉的突變菌株開展了大量研究工作[4,7-8],目前,里氏木霉己經(jīng)成為世界上研究和應(yīng)用最廣泛的纖維素酶工業(yè)生產(chǎn)菌種,世界各國科學(xué)家最初采用物理化學(xué)誘變的方法進行了大量的菌株改良工作,研發(fā)出數(shù)十種的工業(yè)菌株[9]。
非核糖體肽合成酶(NRPS)作為真菌合成次生代謝物的關(guān)鍵酶,是由多個催化結(jié)構(gòu)域組成的高度模塊化的肽合成酶,包括腺苷酰化結(jié)構(gòu)域(A結(jié)構(gòu)域)、巰基化結(jié)構(gòu)域(T結(jié)構(gòu)域)和縮合結(jié)構(gòu)域(C結(jié)構(gòu)域)等[10]。在非核糖體肽(Non-ribosomal peptides, NRP)合成過程中,首先A結(jié)構(gòu)域篩選和激活底物氨基酸,其次底物氨基酸被轉(zhuǎn)運并連接到T結(jié)構(gòu)域,然后經(jīng)過C結(jié)構(gòu)域催化形成肽鍵,最后經(jīng)過多次循環(huán)延伸形成多肽鏈,在硫酯酶結(jié)構(gòu)域(TE結(jié)構(gòu)域)催化下通過水解的方式從NRPS中釋放出來。NRP種類繁多、結(jié)構(gòu)復(fù)雜,在真菌生長發(fā)育、信號傳導(dǎo)及致病等過程中都發(fā)揮著重要的作用[11]。前人研究發(fā)現(xiàn),由蘋果斑點落葉病病菌(Altenaria alternata)產(chǎn)生的AM-毒素[12]、玉米圓斑病病菌(Cochliobolus carbonum)產(chǎn)生的HC-毒素[13],以及玉米小斑病病菌(Cochliobolus heterostrophus)[14]、水稻胡麻斑病病菌(Cochliobolus miyabeanus)[15]、擬南芥黑斑病病菌(Alternaria brassicicola)[15]、柑橘褐斑病病菌(Alternaria alternata)[16]和禾谷鐮刀菌(Fusarium graminearum)中的NPS6基因、蘋果樹腐爛病病菌(Valsa mali)的NPS12基因[11]均與上述病原真菌的致病性有關(guān)[17];同時,前人對于硬枝樹花(Ramalina conduplicans)地衣型真菌中的NRPS基因開展了克隆和鑒定工作[18]。然而,學(xué)術(shù)界尚未見有關(guān)里氏木霉和黏綠木霉中NRPS的數(shù)量和理化性質(zhì)的報道,也未見有關(guān)具有不同功能的木霉中NRPS與其功能關(guān)系的研究報道。
本研究通過SMURF網(wǎng)站預(yù)測里氏木霉和黏綠木霉的次生代謝基因分布情況,獲得編碼聚酮化合物合酶(PKS)、非核糖體肽合成酶以及萜烯合酶(TPS)等基因簇情況,進一步利用antiSMASH對其開展基因組數(shù)據(jù)分析,鑒定和注釋潛在的編碼PKS、NRPS以及TPS的基因,從而得到上述木霉中NRPS蛋白,同時,通過ProtComp v9.0、expasy、PHD等軟件對上述NPRS蛋白進行生物信息學(xué)分析,明確木霉中NRPS的亞細胞定位、理化性質(zhì)以及二級結(jié)構(gòu)等特征,為今后深入研究里氏木霉和黏綠木霉NRPS的功能及其作用機制提供理論依據(jù)。
1材料與方法
1.1木霉中NRPS蛋白找尋
首先,通過SMURF(Secondary Metabolite Unknown Regions Finder)[19](網(wǎng)址為http://smurf.jcvi.org/upload.php)預(yù)測里氏木霉QM6a(全基因組序列來源于http://genome.jgi-psf.org/Trire2/Trire2.home.html)和黏綠木霉GV29-8(全基因組序列來源于http://genome.jgi.doe.gov/TriviGv29_8_2/TriviGv29_8_2.home.html)的次生代謝基因分布情況,獲得編碼PKS、NRPS、TPS等基因簇情況,然后利用antiSMASH(antibiotics & Secondary Metabolite Analysis SHell)[20](網(wǎng)址為https://fungismash.secondarymetabolites.org/#!/start)對上述2種木霉進行基因組數(shù)據(jù)分析,鑒定和注釋潛在的PKS、NRPS、TPS等基因。
1.2NRPS蛋白理化性質(zhì)分析
運用蛋白質(zhì)數(shù)據(jù)庫(網(wǎng)址為http://www.expasy.ch/tools/protparam.html)分析NRPS蛋白的理化性質(zhì),包括理論等電點、不穩(wěn)定性系數(shù)、總平均親水性等,采用Protscale(網(wǎng)址為http://web.expasy.org/protscale/)分析NRPS蛋白的疏水性[21]。
1.3NRPS蛋白二級結(jié)構(gòu)分析
采用PHD(網(wǎng)址為http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)對2種木霉中的NRPS蛋白開展二級結(jié)構(gòu)預(yù)測及分析[22]。
1.4NRPS蛋白轉(zhuǎn)運肽分析
采用TargetP 1.1 Server(網(wǎng)址為http://www.cbs.dtu.dk/services/TargetP-1.1/index.php)分析NRPS蛋白的轉(zhuǎn)運肽[23]。
1.5NRPS蛋白保守結(jié)構(gòu)域分析
采用SMART(網(wǎng)址為http://smart.embl-heidelberg.de/)分析NRPS蛋白的保守結(jié)構(gòu)域[24]。
1.6NRPS蛋白亞細胞定位分析
采用ProtComp v9.0(網(wǎng)址為http://phobius.sbc.su.se/)分析NRPS蛋白的亞細胞定位情況,并對分析數(shù)據(jù)進行整理歸納。
2結(jié)果與分析
2.1黏綠木霉與里氏木霉NRPS蛋白數(shù)量差異
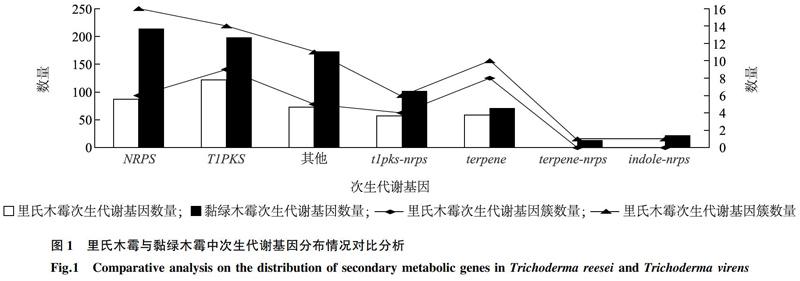
通過SMURF及antiSMASH預(yù)測和基因注釋,發(fā)現(xiàn)里氏木霉QM6a中含有32個次生代謝基因簇,基因簇的大小為6~21個基因,共涉及398個基因;而黏綠木霉GV29-8中則含有59個次生代謝基因簇,基因簇的大小為4~35個基因,共涉及793個基因。里氏木霉含有87個NRPS蛋白,黏綠木霉含有214個NRPS蛋白(圖1)。此外,里氏木霉和黏綠木霉中數(shù)量較多的次生代謝合成基因都為T1PKS和NRPS,但黏綠木霉含有terpene-nrps、indole-nrps。
2.2NRPS蛋白的理論等電點
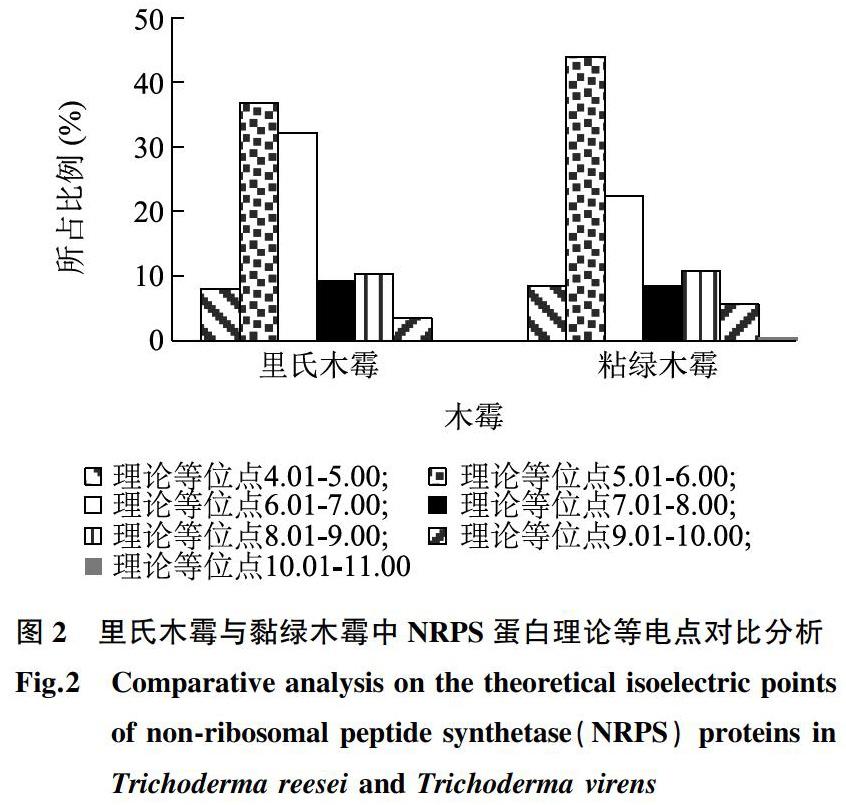
對2種木霉中全部NRPS蛋白的理論等電點進行對比分析,結(jié)果表明,數(shù)值范圍為4.25~10.43,其中,里氏木霉中蛋白ID為XP_006968101.1的NPRS蛋白的理論等電點為4.25,黏綠木霉中蛋白ID為XP_013954358.1的NPRS蛋白的理論等電點為10.43。進一步以理論等電點間隔單位為1.00進行整理分析,發(fā)現(xiàn)里氏木霉與黏綠木霉中NRPS蛋白理論等電點多集中在5.01~7.00,其數(shù)量分別為60、142個,所占比例分別為68.97%、66.36%(圖2)。同時,對屬于酸性蛋白質(zhì)(理論等電點小于6.01)的NRPS蛋白進行分析,里氏木霉與黏綠木霉中NRPS蛋白數(shù)量分別為39、112個,所占比例分別為44.83%、52.34%;對屬于中性蛋白質(zhì)(理論等電點位于6.01~8.00)的NRPS蛋白進行分析,里氏木霉與黏綠木霉中中性NRPS蛋白數(shù)量分別為36、66個,所占比例分別為41.38%、30.84%;對屬于堿性蛋白(理論等電點大于8.00)的NRPS蛋白進行分析,里氏木霉與黏綠木霉中堿性NRPS蛋白數(shù)量分別為12、36個,所占比例分別為13.79%、16.82%(圖2)。由此可見,兩種木霉中NRPS蛋白多屬于酸性蛋白質(zhì)。
2.3NRPS蛋白的不穩(wěn)定性系數(shù)
對2種木霉中全部NRPS蛋白的不穩(wěn)定性系數(shù)進行統(tǒng)計分析,結(jié)果表明,里氏木霉與黏綠木霉中NRPS蛋白的不穩(wěn)定性系數(shù)集中在30.01~50.00,其數(shù)量分別為65、169個,所占比例分別為74.71%、78.97%。同時,對NRPS蛋白中不穩(wěn)定性系數(shù)小于40.01(穩(wěn)定蛋白質(zhì))的分泌蛋白質(zhì)進行分析,里氏木霉與黏綠木霉中穩(wěn)定NRPS蛋白數(shù)量分別為35、93個,所占比例分別為40.23%、43.46%;其次對不穩(wěn)定性系數(shù)大于40.01(不穩(wěn)定蛋白質(zhì))的分泌蛋白質(zhì)進行分析,里氏木霉與黏綠木霉中不穩(wěn)定NRPS蛋白數(shù)量分別為52、121個,所占比例分別為59.77%、56.54%(圖3)。上述結(jié)果表明,兩種木霉中的NRPS蛋白多屬于不穩(wěn)定性蛋白質(zhì)。
2.4NRPS蛋白的親水性
進一步對2種木霉中的全部NRPS蛋白進行親水性分析,結(jié)果表明,NRPS蛋白多屬于親水性蛋白質(zhì)(親水性值小于0),里氏木霉與黏綠木霉中親水性NRPS蛋白數(shù)量分別為69、166個,所占比例分別為79.31%、77.57%;同時,對疏水性蛋白質(zhì)(親水性值大于0)進行統(tǒng)計,里氏木霉與黏綠木霉中疏水性NRPS蛋白數(shù)量分別為18、48個,所占比例分別為20.69%、22.43%。上述結(jié)果表明,里氏木霉和黏綠木霉中NRPS蛋白多屬于親水性蛋白質(zhì)。
此外,進一步對全部NRPS蛋白親水性最強氨基酸殘基、疏水性最強氨基酸殘基種類進行統(tǒng)計分析,結(jié)果顯示里氏木霉中NRPS蛋白親水性最強氨基酸殘基有18種,缺少C(半胱氨酸)、M(甲硫氨酸),而黏綠木霉中NRPS蛋白親水性最強氨基酸殘基有19種,缺少C,存在較多的親水性最強氨基酸殘基都為E(谷氨酸)、D(天冬氨酸)、R(精氨酸)、K(賴氨酸)、N(天冬酰胺);里氏木霉中NRPS蛋白疏水性最強氨基酸殘基有18種,缺少E、Y(酪氨酸),而黏綠木霉中NRPS蛋白疏水性最強氨基酸殘基有20種,存在較多的疏水性最強氨基酸殘基都為A(丙氨酸)、L(亮氨酸)、V(纈氨酸)、G(甘氨酸)、I(異亮氨酸)(圖4)。
2.5NRPS蛋白的二級結(jié)構(gòu)組成特征
對里氏木霉與黏綠木霉中NRPS蛋白進行二級結(jié)構(gòu)預(yù)測,結(jié)果表明,2種木霉中NRPS蛋白均含有無規(guī)則卷曲、α螺旋、β折疊、跨膜螺旋(TM)等二級結(jié)構(gòu),其中,里氏木霉NRPS蛋白中上述二級結(jié)構(gòu)所占比例分別是22.59%、44.36%、12.55%、9.34%,而黏綠木霉NRPS蛋白中所占比例分別21.74%、44.16%、13.22%、9.45%(圖5)。
2.6里氏木霉和黏綠木霉NRPS蛋白的轉(zhuǎn)運肽預(yù)測
對里氏木霉和黏綠木霉中NRPS蛋白的轉(zhuǎn)運肽進行預(yù)測分析,以明確2種木霉中NRPS蛋白的定位情況和預(yù)測可靠性。在定位情況中,里氏木霉NRPS蛋白定位在分泌通路、線粒體及其他位置的數(shù)量分別為11、8、67個,尚有1個NPRS蛋白未能實現(xiàn)預(yù)測;而黏綠木霉NRPS蛋白定位在分泌通路、線粒體及其他位置的數(shù)量分別為35、19、152個,尚有8個NPRS蛋白未能實現(xiàn)預(yù)測(圖6)。
2.7NRPS蛋白中的保守結(jié)構(gòu)域
里氏木霉NRPS蛋白中有79個具有保守結(jié)構(gòu)域,占比為90.80%;黏綠木霉NRPS蛋白中有184個具有保守結(jié)構(gòu)域,占比為85.98%(圖7)。
2.8NRPS蛋白的亞細胞定位
里氏木霉和黏綠木霉中NRPS蛋白亞細胞定位主要集中在線粒體、細胞質(zhì)膜和細胞質(zhì)等,里氏木霉中在上述細胞器中所占比例分別是33.33%、19.54%、16.09%,黏綠木霉中在上述細胞器中所占比例分別是26.64%、21.96%、18.69%(圖8)。
3討論
前期對里氏木霉QM6a和黏綠木霉GV29-8中的分泌蛋白質(zhì)及CAZymes蛋白開展了預(yù)測分析,研究結(jié)果表明,前者含有分泌蛋白的數(shù)量為356個,占全部蛋白質(zhì)數(shù)量的3.89%,而CAZymes蛋白數(shù)量為201個,占全部蛋白質(zhì)數(shù)量的2.20%[4, 25];黏綠木霉GV29-8中分泌蛋白質(zhì)數(shù)量為377個,占全部蛋白質(zhì)數(shù)量的3.03%,而CAZymes蛋白數(shù)量為185個,占全部蛋白質(zhì)數(shù)量的1.49%[2,26]。由此可見,里氏木霉QM6a在分泌蛋白質(zhì)及CAZymes蛋白占全部蛋白質(zhì)的比例方面均高于黏綠木霉GV29-8,然而,前者中的NRPS數(shù)量為87個,占全部蛋白質(zhì)數(shù)量的0.95%,而黏綠木霉GV29-8中NRPS數(shù)量高達214個,占全部蛋白質(zhì)數(shù)量的1.72%,前者NRPS蛋白所占比例明顯低于后者。基于上述研究結(jié)果,根據(jù)里氏木霉QM6a作為重要的纖維素降解工業(yè)菌株以及黏綠木霉GV29-8作為重要的生防菌株的判斷,我們推斷,分泌蛋白質(zhì)及CAZymes蛋白在兩種木霉中均對實現(xiàn)其破壞植物細胞壁組織方面發(fā)揮著重要作用,而NPRS則在實現(xiàn)其破壞植物纖維組織方面發(fā)揮著更加重要的作用,terpene-nrps和indole-nrps在破壞植物病原真菌的細胞壁組織方面發(fā)揮著重要作用。當(dāng)然,是否存在著時間、空間方面的差異性,有待于進一步通過生物學(xué)試驗進行驗證。
同時,本研究通過次生代謝基因預(yù)測和注釋還明確了里氏木霉QM6a中含有32個次生代謝基因簇,而黏綠木霉GV29-8中則有59個次生代謝基因簇。在這些次生代謝基因簇中不僅編碼NPRS蛋白,還涉及T1PKS、terpene-nrps、indole-nrps等蛋白質(zhì)。上述蛋白質(zhì)在實現(xiàn)利用植物纖維方面的作用有待進一步明確,具有不同功能的木霉與次生代謝基因簇之間的關(guān)系也有待進一步明確。然而,盡管2種木霉在NPRS蛋白數(shù)量及次生代謝基因簇數(shù)量方面均存在著明顯不同,但是其NPRS在理化性質(zhì)、亞細胞定位、二級結(jié)構(gòu)等方面存在著較多相同特征。基于此,我們推斷,2種木霉中NPRS蛋白在功能方面具有同質(zhì)性,而木霉功能的差異性發(fā)揮則表現(xiàn)在其數(shù)量上。
本研究通過SMURF及antiSMASH預(yù)測網(wǎng)站對里氏木霉QM6a和黏綠木霉GV29-8進行了次生代謝產(chǎn)物基因找尋及NPRS的生物信息學(xué)分析,結(jié)果表明,前者含有87個NRPS蛋白,而后者含有214個NRPS蛋白,后者所含有的NPRS蛋白數(shù)量明顯高于前者;同時,明確2種木霉中NRPS蛋白具有以下共同特征:理論等電點多集中在5.01~7.00,不穩(wěn)定性系數(shù)多集中在30.01~50.00,高達75%以上的蛋白質(zhì)屬于親水性蛋白質(zhì),而二級結(jié)構(gòu)組成特征則以α螺旋為主,轉(zhuǎn)運肽情況尚未明確,保守結(jié)構(gòu)域較多,亞細胞定位多集中在線粒體等。就理化性質(zhì)而言,上述木霉中NRPS蛋白的理化性質(zhì)以酸性、不穩(wěn)定性和親水性為主,親水性最強氨基酸主要有E、D、R、K、N等,疏水性最強氨基酸主要有A、L、V、G、I等。同時,明確里氏木霉NRPS蛋白相比于黏綠木霉親水性最強氨基酸中缺少M,而疏水性最強氨基酸中則缺少E、Y。研究結(jié)果為進一步開展里氏木霉QM6a和黏綠木霉GV29-8中次生代謝產(chǎn)物基因利用和改造,以及進一步開展不同木霉之間NPRS功能差異化研究提供了基礎(chǔ)。
[18]原曉龍,趙能,陳劍,等. 硬枝樹花中非核糖體多肽合成酶NRPS基因的克隆與鑒定[J]. 基因組學(xué)與應(yīng)用生物學(xué),2016, 35 (10):2834-2841.
[19]KHALDI N,SEIFUDDIN F T,TURNER G, et al. SMURF: Genomic mapping of fungal secondary metabolite clusters[J]. Fungal Genet Biol, 2010, 47 (9):736-741.
[20]BLIN K, WOLF T, CHEVRETTE M G, et al. antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification[J]. Nucleic Acids Res,2017, 45 (W1):36-41.
[21]GASTEIGER E,GATTIKER A, HOOGLAND C, et al. ExPASy: The proteomics server for in-depth protein knowledge and analysis[J]. Nucleic Acids Research,2003, 31 (13):3784-3788.
[22]LAWRENCE A K, STEFANS M, CHRISTOPHER M Y, et al. The Phyre2 web portal for protein modeling, prediction and analysis[J]. Nat Protocols,2015, 10 (6): 845-858.
[23]OLOF E, SREN B, GUNNAR V H, et al. Locating proteins in the cell using TargetP, SignalP and related tools[J]. Nature Protocols,2007, 2 (4):953-971.
[24]IVICA L, TOBIAS D, PEER B. SMART 7: recent updates to the protein domain annotation resource[J]. Nucleic Acids Research,2012, 40 (D1):302-305.
[25]韓長志,王娟. 基于全基因組序列預(yù)測里氏木霉QM6a的分泌蛋白[J]. 華中農(nóng)業(yè)大學(xué)學(xué)報,2017, 36 (2):28-32.
[26]韓長志. 基于全基因組測序的粘綠木霉Gv29-8中分泌蛋白預(yù)測[J]. 河南師范大學(xué)學(xué)報(自然科學(xué)版), 2016, 44 (6):145-148.
(責(zé)任編輯:張震林)
