外加碳源和氮源對酸化水稻土溫室氣體釋放的影響
2020-12-07 05:31:18朱春權韋翠珍曹小闖朱練峰孔亞麗金千瑜張均華
中國稻米 2020年6期
朱春權 韋翠珍 曹小闖 朱練峰 孔亞麗 金千瑜 張均華*
(1中國水稻研究所/水稻生物學國家重點實驗室,杭州310006;2 淮河水資源保護科學研究所,安徽蚌埠233001;第一作者:zhuchunquan@caas.cn;*通訊作者: zhangjunhua@caas.cn)
全球約有39.5 億hm2酸性土壤,占潛在可耕地面積的40%~50%[1]。我國酸性土壤廣泛分布在東南部熱帶、亞熱帶地區,面積多達218 萬km2,約占全國陸地面積的22.7%[2]。中國南方水稻田多為酸性土壤。土壤酸化導致土壤板結,質地粘重,土壤結構遭到破壞,通氣性變差,養分失調,肥力變差,從而限制作物的養分吸收,降低作物產量[3]。
溫室氣體已經對環境造成明顯的負面影響[4],其中氮氧化物(Nitrogen Oxide,NOx)是溫室氣體的重要組成成分[5-6],它包含如氧化亞氮(N2O)、一氧化氮(NO)和二氧化氮(NO2)等在內的多種化合物。土壤是生態系統NO 產生的重要來源[7]。其中土壤微生物在土壤NO 等溫室氣體釋放過程中發揮著重要作用,包括硝化反應和反硝化作用在內的微生物活動均可產生溫室氣體NO 和N2O[8],并且進一步酸化土壤,從而影響植物的生長。
南方酸化土壤作為我國亞熱帶地區的主要土壤資源,其對溫室氣體NO 和N2O 的釋放需要重點研究。前人對安徽郎溪的耕地酸化土壤進行了3 種不同氮源(尿素、硫酸銨和碳酸氫銨)的施加試驗后發現,3 種化學氮肥均能顯著刺激土壤的硝化反應,從而進一步降低土壤的pH 值[9]。對湖南祁陽長期定位試驗站的酸化土壤進行分析后發現,與不施加任何肥料的酸化土壤相比,長期施加氮磷鉀肥以及有機肥料的土壤N2O 的釋放顯著增加[10]。由此可見,雖然酸化土壤擁有較低的pH 值,但其仍然存在硝化和反硝化作用,并且施加肥料后,其硝化和反硝化作用進一步增強,從而擴大氮氧化物的釋放以及刺激土壤進一步酸化。雖然前人的研究關注到了微生物在氮氧化物釋放中的作用,但是僅限在施肥后微生物豐度和群落的變化,并未直接關注微生物變化后對土壤溫室氣體釋放的影響。因此,本試驗選擇了浙江富陽酸化水稻土,單獨施加硫酸銨和同時施加硫酸銨以及葡萄糖,進行短期和長期培養后觀察氮氧化物的釋放以及微生物豐度的變化,以進一步確認微生物本身對酸化水稻土硝化和反硝化的作用。
1 材料與方法
1.1 供試土壤
供試土壤樣品采集于浙江富陽常耕水稻土(0~20 cm)深度。土樣經風干、磨細過2 mm 孔徑篩后備用,具體土壤性質見表1。
1.2 試驗設計
設置不施氮(CK)與施N 250 mg/kg 2 個氮施用水平,250 mg/kg 氮施用水平又分為單獨施加硫酸銨(N)和同時添加硫酸銨及100 mg/kg 葡萄糖(N+C)2 個處理組,各處理均設3 個重復。處理過程如下:稱取50 g風干土壤置于250 mL 的廣口瓶中,通過滴加超純水使其含水量達到田間持水量的40%,用無菌透氣封口膜封閉瓶口。在25℃黑暗條件下預培養1 周。然后配置相應的硫酸銨和葡萄糖溶液,滴加到預培養的土壤中,施用的量分別為250 mg/kg N 和100 mg/kg C,并且土壤的最終含水量維持在田間持水量的60%。用無菌透氣封口膜封閉瓶口,25℃黑暗條件下培養。每隔1 周補充水分,使其保持恒定的含水量。在培養的第2 d 和120 d 測定NO 和N2O 釋放后進行破壞性取樣,用鮮土提取土壤DNA 以及測定土壤無機氮(NH4+和NO3-)含量,然后將土壤樣品風干、過篩待用。

表1 土壤基本理化性質

表2 本試驗中使用的引物序列
1.3 測定項目及方法
1.3.1 NO 及N2O
土壤NO 和N2O 釋放速率的測定采用靜態箱法[4],其基本過程如下:裝有土壤樣品的廣口瓶用橡膠塞密閉1 h,橡膠塞同時帶有上進氣管和出氣管,管口用氣墊進行封閉。1 h 后用10 mL 注射器抽取密閉氣體,用氣相色譜儀(Agilent 7890,California,USA)測定 N2O 含量。與此同時,打開瓶蓋放氣后再次封閉瓶口,1 h 后用氮氧化物測定儀直接測定NO 含量,測定時同時打開出氣管與進氣管,出氣管與氮氧化物分析儀相連,記錄NO 的數值,測定時間一般為NO 的數值上升后下降至測定前數值為止。
N2O 和NO 的測定結果均換算為nmol/kg/h。
1.3.2 土壤理化性質
土壤 pH 值 采用復合電極法測定(V 土∶m 水=1∶2.5)。土壤 NH4+和 NO3-用 2 mol/L 的 KCl 以 1∶10 浸提后,采用連續流動分析儀(San++System,Netherlands)測定。
1.3.3 土壤微生物豐度
土壤總DNA 的提取使用提取試劑盒 FastDNA?SPIN Ki(tMP Biomedicals,Santa Ana,CA),提取步驟按其說明書進行。提取后的DNA 用nanodrop 測定濃度,然后分別稀釋成10 ng/μL 待用。
本試驗選取了細菌的16S rRNA 基因[11]、氨氧化細菌和氨氧化古菌的氨單加氧酶(amoA)基因[12]、反硝化細菌的亞硝酸還原酶基因(nirK 和nirS)[13-14]以及一氧化二氮還原酶基因(nosZ)[14]進行定量分析。試驗體系為5 μL SYBR Green(Takara Bio, Inc., 日本),0.2 μL 正向引物,0.2 μL 反向引物,1 μL DNA 模板和 3.6 μL 滅菌水,總計10 μL。配制好的體系,在熒光定量PCR 儀(Light Cycler 480 II, Roche, Switzerland)上進行擴增。同時用連接相應基因片段的PMD-18T 質粒進行梯度稀釋后做標準曲線[15]。相關基因的引物以及反應條件見表2。
1.4 統計學分析
每個試驗重復3 次,用SPSS 軟件進行單因素方差分析,平均值用Tukey 法進行多重比較。相關性分析采用pearson 雙尾檢驗分析法。
2 結果與分析
2.1 土壤 pH 值、NH4+-N 和 NO3-N 含量變化
添加硫酸銨后水稻土pH 值顯著下降(0 d),其原因在于硫酸銨屬于酸性肥料。短期培養(2 d)后pH 值進一步下降,并且施加硫酸銨和同時施加硫酸銨與葡萄糖的2 個處理組間pH 值變化無顯著差異(圖1)。長期培養(120 d)進一步降低了土壤pH 值,并且同時施加硫酸銨與葡萄糖處理組的pH 值顯著小于單獨施加硫酸銨的處理(圖1)。
添加硫酸銨、硫酸銨與葡萄糖后,短期培養和長期培養下,土壤NH4+-N 含量和NO3--N 含量均顯著增加;隨著培養時間的延長,土壤NH4+-N 含量顯著降低,土壤NO3--N 含量顯著增加,并且同時施加硫酸銨和葡萄糖處理組的NH4+-N 降低程度和NO3--N 含量增加程度均顯著大于單數施加硫酸銨處理組(圖2)。說明外加碳源可以顯著促進土壤中氮肥的轉化。
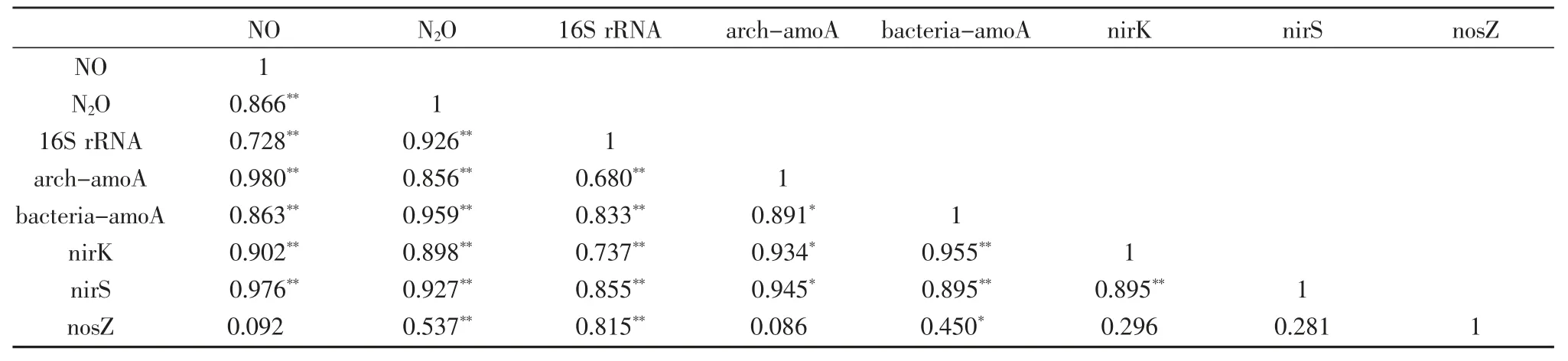
表3 相關性分析

圖1 處理2 d 和120 d 后土壤pH 值變化
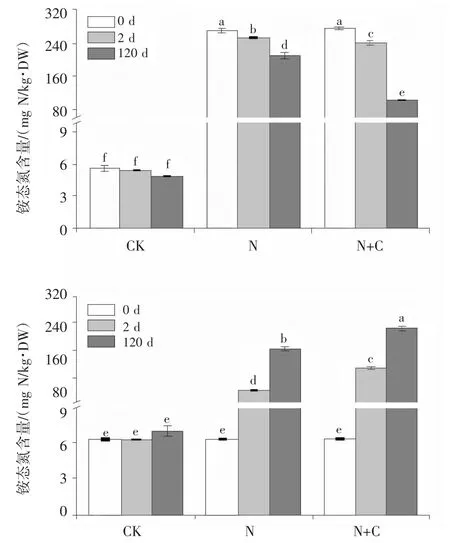
圖 2 處理 2 d 和 120 d 后土壤(A)NH4+和(B)NO3-含量的變化
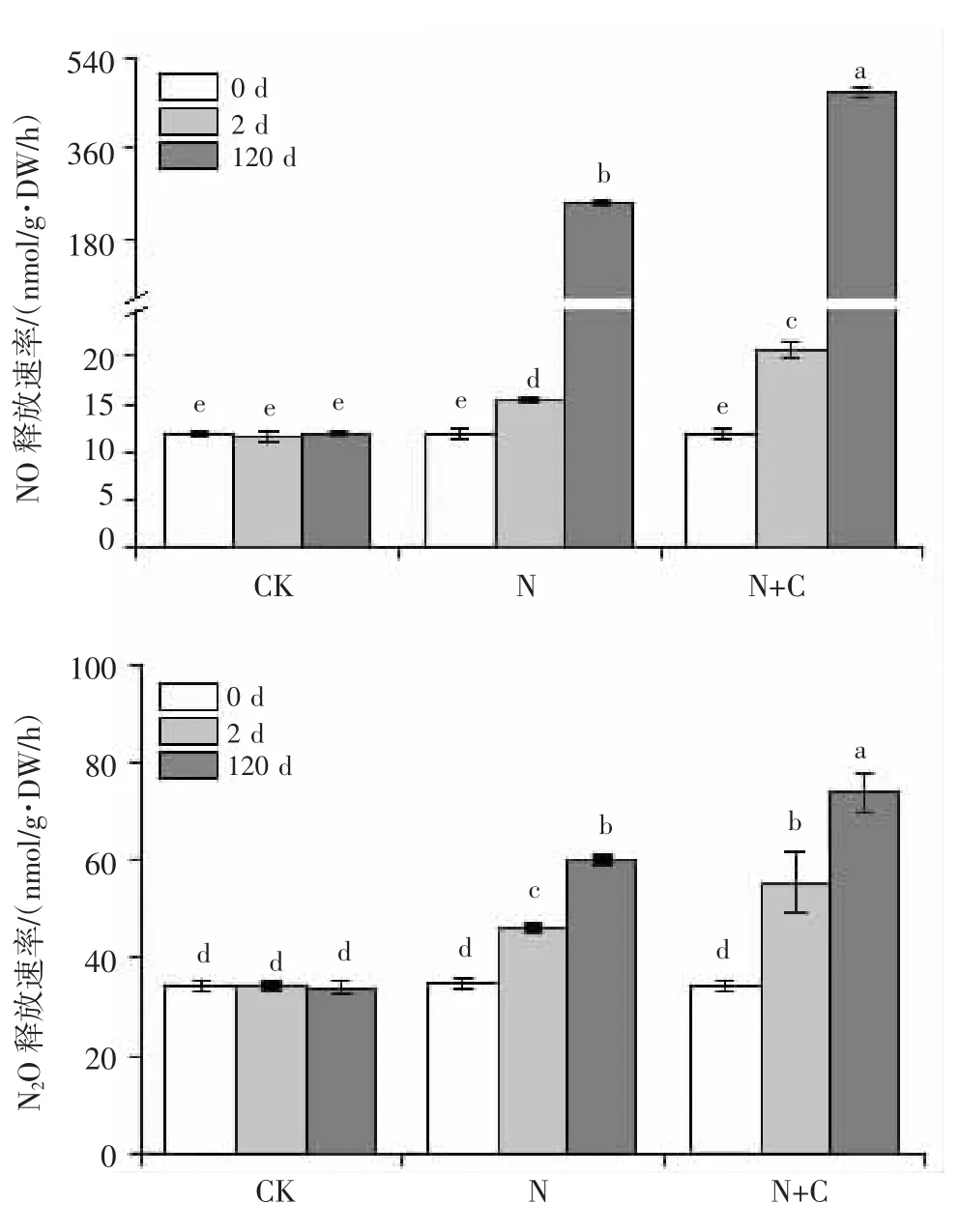
圖 3 處理 2 d 和 120 d 后土壤(A)NO 和(B)N2O 釋放
2.2 土壤NO 和N2O 釋放速率
短期培養和長期培養下,外源施加硫酸銨后均顯著促進土壤NO 和N2O 的釋放,并且外源添加葡萄糖后進一步提高了NO 和N2O 的釋放速率(圖3)。說明外加碳源顯著促進施氮土壤中溫室氣體的產生。
2.3 土壤微生物豐度變化
從圖4A 可見,外源添加硫酸銨短時間內(2 d)就顯著增加土壤中細菌(16s rRNA)的豐度,并且隨著培養時間的延長(120 d),土壤細菌的豐度進一步增加;外源添加葡萄糖后,進一步增加了土壤中的細菌豐度。
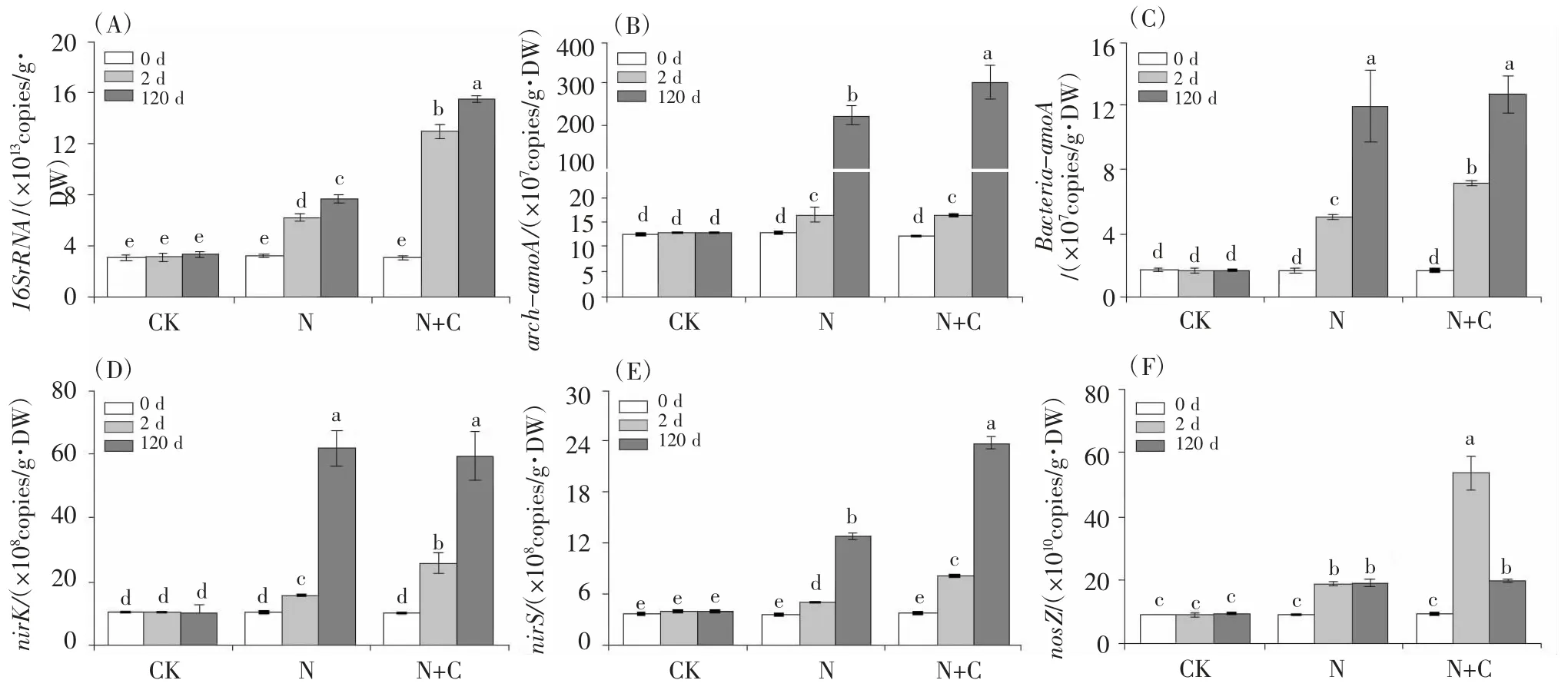
圖 4 處理 2 d 和 120 d 后土壤(A)總菌、(B)厭氧氨氧化細菌、(C)氨氧化細菌和反硝化細菌、(D)nirK 基因、(E)nirS 基因和(F)nosZ基因豐度變化豐度的變化
氨氧化古菌(bacteria - amoA)和氨氧化細菌(bacteria - amoA)均催化銨態氮向硝態氮的轉化,并在此過程中產生NO 和N2O。研究結果顯示,添加硫酸銨后短時間培養(2 d)就顯著提高土壤中氨氧化古菌和氨氧化細菌的豐度,并且在長期培養(120 d)后進一步提高了兩種菌的豐度(圖4 B 和圖4 C)。短期培養(2 d)時,同時施加硫酸銨和葡萄糖的處理氨氧化細菌豐度大于單獨施加硫酸銨處理組;長期培養(120 d)時,同時施加硫酸銨和葡萄糖處理組的氨氧化古菌豐度大于單獨施加硫酸銨處理組,說明添加葡萄糖后,氨氧化細菌和氨氧化古菌分別在短期培養和長期培養時發揮作用,從而導致不同量的NO 和N2O 釋放。
土壤中反硝化細菌促進NO3-向NO 的轉變(nirK,nirS)并最終轉化為N(nosZ)。研究結果顯示,添加硫酸銨后短時間培養(2 d)就顯著提高土壤中nirK 和nirS基因的豐度,并且在長期培養(120 d)后進一步提高了nirK 和nirS 基因的豐度(圖4 D 和圖4 E)。短期培養(2 d)時,同時施加硫酸銨和葡萄糖處理組的nirK 和nirS 基因豐度大于單獨施加硫酸銨處理組,長期培養(120 d)時,同時施加硫酸銨和葡萄糖處理組的nirK 基因豐度大于單獨施加硫酸銨處理組。添加硫酸銨和同時添加硫酸銨和葡萄糖雖然在短期培養時增加了nosZ基因的豐度,但是在第120 d 時nosZ 基因豐度沒有進一步增加,甚至出現顯著下降(圖4F),從而阻止了NO向N 的轉化,最終導致了NO 的積累。
2.4 相關性分析
相關性分析發現,溫室氣體NO 和N2O 的釋放與氨氧化細菌(bacteria-amoA)、氨氧化古菌(arch-amoA)和反硝化細菌中的nirK 和nirS 基因豐度呈極顯著正相關。總菌16S rRNA 基因豐度的變化與氨氧化細菌、氨氧化古菌和反硝化細菌豐度變化呈極顯著正相關。
3 討論
3.1 酸化水稻土NO 及N2O 氣體釋放特征
前人研究表明,土壤中NO 的釋放與土壤基質中的無機氮(NH4+和NO3-)含量和凈硝化速率密切相關,如VELDKAMP 等在對23 種農田生態系統的研究中發現,NO 釋放的量和土壤無機氮含量之間存在顯著的正相關。STARK 等[16]在對9 種溫帶森林和牧場的土壤進行研究后也得出類似的結論。與前人的研究一致,本試驗在添加外源硫酸銨后,試驗土壤的溫室氣體NO 和N2O 釋放速率均顯著上升,遠高于對照組(圖3)。土壤的無機氮是土壤中硝化細菌以及反硝化細菌代謝活動的底物,而土壤生物源NO 的產生正是這兩類細菌活動的結果[17]。因此,外源施加氮可顯著增加溫室氣體NO 或N2O 的產生。本試驗結果顯示,單獨施加硫酸銨和同時施加硫酸銨和葡萄糖處理組的NH4+離子含量,均隨培養時間的延長而減少,同時,NO3-離子含量隨培養時間的延長均相應上升(圖2),說明其中微生物的硝化或反硝化作用對土壤的氮源進行了轉化,并在轉化過程中產生了溫室氣體。
在土壤微生物對土壤銨態氮進行利用的過程中,土壤的pH 值會發生顯著變化。本研究結果顯示,2 個處理的pH 值均隨著培養時間的延長而顯著下降(圖1)。單獨施加硫酸銨以及同時施加硫酸銨和葡萄糖的處理pH 值在第2 d 即顯著下降。原因在于硫酸銨是一種酸性肥料,由于其本身的酸性作用,導致土壤的pH值下降,但是后期土壤pH 值進一步下降的原因則是其中硝化或反硝化作用導致NH4+離子氧化,在土壤中留下 H+。
土壤的pH 值是影響土壤硝化和反硝化作用的決定因素之一[18-20]。微生物的硝化反應主要發生在通氣良好并且pH 值大于4.0 的土壤中。REMDE 等研究發現,在堿性土壤(pH 值7.8)中,NO 主要是由硝化反應產生的,在酸性土壤(pH 值 4.7)中,NO 主要是由反硝化作用產生的。過高或者過低的土壤pH 值都會影響土壤的硝化反應,從而減少NO 等溫室氣體的釋放[12]。JIANG 等[19]研究也發現,pH 值還是決定水稻土硝化過程中起主導作用的菌類的決定因素之一,即在酸性水稻土中,土壤的硝化反應主要由氨氧化古菌主導,而在堿性水稻土中,土壤的硝化反應主要由氨氧化細菌主導。本次試驗采用的土壤來自于酸化水稻土,暗示氨氧化古菌可能在原始酸化水稻土通過硝化反應產生溫室氣體過程中發揮重要作用。
3.2 細菌豐度與NO 和N2O 釋放
土壤NO 和N2O 的產生主要通過土壤微生物對其中氮素的轉化產生,其中硝化和反硝化細菌是NO 和N2O 的主要產生者[21]。土壤中硝化細菌將NH4+氧化成NO3-離子,反硝化細菌把NO3-最終還原成N2,在硝化和反硝化過程中產生NO 和N2O。土壤微生物的硝化作用包括自養硝化作用和異養硝化作用[22]。在通氣良好的土壤中,自養硝化作用是NO 氣體釋放的主要來源[7,23];而在低pH 即酸性環境中,異養硝化作用在NO 產生中發揮重要作用。同時,在通氣較差的土壤中,反硝化作用將發揮主要作用,將NO3-或NO2-還原為N2或中間產物NO 和N2O[24]。本試驗的相關性分析表明,NO 和N2O的釋放均與氨氧化細菌、氨氧化古菌以及反硝化細菌的中的nirK 和nirS 基因豐度呈顯著正相關,說明本試驗外源添加氮源后的溫室氣體釋放主要是由于硝化及反硝化作用產生的。
前人的研究表明,加入葡萄糖后能增加土壤N2O排放,這可能在于葡萄糖的加入為土壤微生物還原NO3-提供了碳源[25]。本試驗研究結果與前人一致,即同時施加硫酸銨和葡萄糖處理的溫室氣體釋放在短期培養和長期培養后均顯著大于單獨施加硫酸銨處理組(圖3),說明外加碳源顯著增加酸化水稻土溫室氣體的釋放。在短期培養時,溫室氣體的釋放和氨氧化古菌、反硝化細菌中的nirK 和nirS 基因的豐度變化一致,說明短期培養時外加碳源提高土壤溫室氣體釋放主要通過氨氧化古菌和反硝化作用進行。在長期培養后,隨著微生物豐度的變化,溫室氣體釋放的趨勢主要與氨氧化細菌和反硝化細菌中的nirS 基因的豐度一致,同時nosZ 基因的豐度顯著下降,抑制了NO 轉化成N2,說明長期培養時添加碳源提高的溫室氣體主要是通過提高氨氧化細菌和反硝化作用進行的。
相關性分析表明,本試驗中氨氧化細菌和氨氧化古菌的基因豐度變化與反硝化細菌中nirK 和nirS 基因的豐度存在極顯著相關,說明外源施加氮源后,隨著硝化作用的進行,進一步帶動反硝化作用,從而最終產生溫室氣體。
4 結論
單獨施加硫酸銨和同時施加硫酸銨和葡萄糖均可通過提高硝化和反硝化細菌的豐度來增加溫室氣體NO 和N2O 的排放,并且在短期培養和長期培養過程中,外加碳源主要通過調控不同的細菌種類產生溫室氣體。本研究結果為解釋酸化水稻土溫室氣體排放機制提供了理論支持,為進一步研究全球溫室化過程提供了理論基礎,比如解釋秸稈(可作為外源碳源)還田過程中稻田溫室氣體釋放的機制等。
