番鴨產蛋前期和產蛋期肝臟組織非靶向代謝組學比較分析
2020-11-26 13:55:44朱文俊楊萬里韋聰聰耿照玉陳興勇
中國農業大學學報 2020年12期
關鍵詞:差異
朱文俊 楊萬里 韋聰聰 劉 樂 耿照玉,2 陳興勇,2*
(1.安徽農業大學 動物科技學院,合肥 230036;2.安徽農業大學 地方畜禽遺傳資源保護與生物育種安徽省重點實驗室,合肥 230036)
番鴨為優良瘦肉型鴨,具有生長快、體重大、耐粗飼和肉質好等特點,但其繁殖性能差,產蛋率低,且產蛋量在個體間存在顯著差異[1]。肝臟為機體物質代謝主要器官,在脂質合成、降解和運輸過程中起重要作用[2]。不同于哺乳動物,禽類脂質約90%于肝臟中合成。激素刺激肝臟合成低密度脂蛋白(LDL)和極低密度脂蛋白(VLDL)等脂類物質,通過血液運輸轉運到機體各部位。肝臟合成的脂類經受體介導沉積于卵泡中,促進等級前和等級卵泡發育[3]。此外,甘油磷脂、膽固醇、甘油三酯、載脂蛋白B等在產蛋期肝臟中大量合成[4-5],可能與卵黃前體物形成和卵泡發育密切相關。推測,產蛋前后肝臟物質合成差異可能在卵黃前體物合成過程中發揮重要作用。
已有研究表明家禽產蛋前后肝臟組織基因差異表達。Li等[2]比較雞產蛋前后(20和30周齡)肝臟脂質代謝相關基因表達譜及相關通路差異,發現差異表達基因在氧化還原、甾醇和膽固醇代謝過程、脂質生物合成過程中顯著富集,表明產蛋時期肝臟轉錄組水平大部分變化與脂肪代謝密切相關;Bourin等[6]采用轉錄組學方法定義肝臟中與卵黃生成有關蛋白酶和抗蛋白酶基因,發現3個蛋白酶和反蛋白酶可能參與蛋黃形成。Li等[7]使用轉錄組學比較IncRNA在產蛋前期和產蛋期蛋雞肝臟表達差異,發現一系列與雞肝臟脂質代謝相關的IncRNA;IncLTR受通過ERβ信號傳導的雌激素調控,與雞屠宰性狀和血液甘油三酯含量有關。盡管前人在家禽產蛋前后對肝臟組織進行轉錄組相關分析,解釋產蛋前后基因轉錄水平表達差異,但受基因表達和翻譯水平調控影響,產蛋前后肝臟轉錄組學研究不能完全解釋其參與肝臟代謝物合成的產物類型。
非靶向代謝組學(Nontargeted metabolomics, NTM)通過檢測一定質量范圍內所有離子,包括結構獨特代謝物離子,為生物機體提供一個無偏倚的整體代謝物譜[8-9]。靶向代謝組學只對有限的幾個或幾類與生物學事件相關的代謝物進行分析和研究,因此無法檢測未知代謝物。NTM可有效捕捉機體生命活動過程中的細微變化,目前已成為畜禽育種、食品安全等領域研究的重要工具,包括畜禽經濟性狀育種價值評估、畜禽產品安全性評價和動物醫學模型使用等。Wen等[10]使用UHPLC-MS非靶向代謝組學測定冷藏雞肉質量,發現隨著冷藏時間的延長,大多數氨基酸豐度降低,而組胺、酪胺、色氨酸和N-乙酰半胱氨酸等豐度增加。Yuan等[11]通過UHPLC-MS非靶向代謝組學分析發現雞十二指腸中幾種糖類如纖維二糖,甘露糖和阿洛糖可能與蒼白桿菌和紅球菌共同降解木質纖維素有關。本研究采用UPLC-MS非靶向代謝組學技術,分析產蛋期與產蛋前期肝臟代謝物譜差異,為肝臟代謝物在卵泡發育中的調控機制提供新的研究視角。
1 材料與方法
1.1 試驗材料
產蛋前期(22周,2 152±73 g,TT 對照組)和產蛋期(40周,2 760±94 g,FT試驗組)的健康番鴨各12羽地面平養于安徽安慶永強農業開發股份有限公司種鴨場。22周種鴨光照11 h/d,采食量103 g/d;40周齡種鴨光照17 h/d,采食量160 g/d,各飼養階段的營養水平均按國家研究委員會(NRC)標準進行[12]。屠宰后,快速收集肝右葉樣本于液氮中保存,后轉移至超低溫冰箱中儲存備用。
1.2 樣品前處理
肝臟組織樣本前處理方法按照He等[13]方法制備,并做適當修改。準確稱取30 mg肝臟組織樣本轉移至1.5 mL EP管,加入內標(L-2-氯苯丙氨酸,0.3 mg/mL;Lyso PC 17∶0,0.01 mg/mL)各20 μL,加入甲醇-水溶液(V∶V=4∶1)400 μL。-20 ℃預冷樣品2 min后,于全自動樣品快速研磨儀中60 Hz研磨2 min。冰水浴中超聲10 min后于-20 ℃環境下靜置20 min。13 000 rpm、4 ℃離心10 min,取300 μL上清液真空干燥。加入400 μL甲醇-水溶液(V∶V=1∶4),渦旋30 s后超聲 2 min 以充分復溶樣品。13 000 rpm、4 ℃離心樣品10 min,吸取上清液150 μL。使用0.22 μm有機相針孔過濾器過濾上清液并轉移至LC進樣小瓶,-80 ℃ 保存,直至LC-MS上機分析。質控樣本(QC)由所有樣本提取液等體積混合制備而成,每個QC體積與樣本相同。所有提取試劑使用前均在 -20 ℃ 預冷。
1.3 代謝物檢測
代謝物檢測方法按照He等[13]方法進行,并做適當修改。ACQUITY UPLC超高效液相系統(Waters公司, USA)串聯QE高分辨質譜儀系統(Thermo Fisher公司, USA)用于分析ESI正負離子模式下的代謝譜。所使用色譜柱為ACQUITY UPLC BEH C18(2.1 mm×100 mm, 1.7 μm)。二元梯度洗脫體系由流動相A:水+0.1%甲酸,B:乙腈+0.1%甲酸組成。梯度洗脫程序如下:0 min, 5% B;1 min, 5% B;12 min, 95% B;14 min, 95% B;14.1 min, 5% B;15 min, 5% B。流速:0.35 mL/min,柱溫:40 ℃。自動進樣器溫度:4 ℃,進樣體積:10 μL。
樣品質譜信號采集分別采用正負離子掃描模式。質譜參數如下:噴霧電壓:3 800V(+)/3 800V(-);毛細管柱溫度:320 ℃(+)/320 ℃(-);探針加熱溫度:350 ℃(+)/350 ℃(-);鞘氣壓:45 Arb(+)/5 Arb(-);輔氣壓:10 Arb(+)/10 Arb(-);S-lens RF level:50(+)/50(-);質量范圍:100-1 200 m/z(+)/100-1 200 m/z(-);一級質譜分辨率:70 000(+)/70 000(-);二級質譜分辨率:17 500(+)/17 500(-);NCE/stepped NCE:10,20,40eV(+)/10,20,40eV(-)。質譜分析過程中,每6個正式樣本中插入一個QC樣本,以用于評價整個試驗過程中系統質譜平臺穩定性。
1.4 數據統計分析
UPLC-MS獲得原始數據經代謝組學處理軟件Progenesis QI軟件(V2.3,UK)作基線過濾、峰識別、積分、保留時間校正、峰對齊和歸一化。化合物鑒定基于精確質量數、二級碎片以及同位素分布,使用 The Human Metabolome Database(HMDB)、Lipidmaps(V2.3)以及METLIN數據庫進行定性。使用包含保留時間、質荷比、峰值強度和樣本信息的數據矩陣作統計分析。每個樣本、每個代謝物對應峰值強度由該樣本總峰值強度校正。將正、負離子數據合并導入R包中作多元統計分析。采用主成分分析(PCA)與正交偏最小二乘方-判別分析(OPLS-DA)對試驗組間代謝物變化作可視化分析。OPLS-DA分析中,變量權重值(VIP)用于衡量各代謝物表達模式對各組樣本分類判別的影響強度和解釋能力。為防止模型過擬合,7次循環交互驗證和200次響應排序檢驗方法用于考察模型質量。篩選VIP>1,P<0.05的差異代謝物。使用KEGG pathway(https:∥www.kegg.jp/)數據庫對肝臟樣品差異代謝物作通路富集分析。獨立樣本T檢驗用于比較對照組和試驗組的差異。
2 結果與分析
2.1 肝臟代謝物定性分析
所有肝臟組織樣本均在UPLC-MS平臺上分析,代謝物在正、負離子模式下得到較好分離(圖1)。經The Human Metabolome Database、Lipidmaps(V2.3)以及METLIN數據庫定性,共鑒定到4 519種代謝物,其中負離子模式鑒定到1 708種代謝物,正離子模式鑒定到2 811種代謝物。
老陸變化太大了。看上去比你還要老(我老嗎?),頭發都掉光了,禿腦門兒。上學那會,也算是個帥哥,現在整個一老頭模樣,簡直不敢認了。你知道他是做什么的嗎,教育局當處長,你知道他開的什么牌子的車,奧迪Q 5,我操,六七十萬吧!
2.2 多元統計分析
主成分分析(PCA)模型中QC樣本緊密聚集,表明試驗穩定性和重復性較好(圖2(a))。R2X為決定PCA模型質量的主要參數,FT/TT組的R2X(cum)=0.507>0.5,表明PCA模型可靠。此外,PCA圖也顯示FT組和TT組完全分離,2組間差異顯著(圖2(b))。建立OPLS-DA模型進一步驗證代謝譜在FT組與TT組之間的分離。OPLS-DA模型參數R2Y(cum)=0.983,Q2(cum)=0.963均>0.5,表明模型建立較好。OPLS-DA圖中,FT組和TT組(圖2(c))完全分離,進一步表明不同產蛋時期肝臟代謝物存在顯著差異。7次循環交互驗證和200次響應排序檢驗的結果顯示,Q2=-0.597<0,表明OPLS-DA模型沒有過擬合(圖2(d))。上述結果表明,FT組和TT組的肝臟樣本在PCA模型和OPLS-DA模型中均差異顯著,所獲得的數據可用于差異代謝物的篩選。
2.3 差異代謝物豐度變化分析
與TT組相比,在FT組中共篩選出324種差異代謝物,其中168種代謝物豐度顯著下調(P<0.05),156種代謝物豐度顯著上調(P<0.05)。為更清楚了解代謝物豐度變化,繪制了324種差異代謝物熱圖(圖3)。熱圖分為2個聚類,每種差異代謝物在FT和TT組之間豐度均存在顯著差異(P<0.05)。根據物質信息進一步分類可將差異代謝物歸為12種類型:苯環型化合物、脂類及類脂分子、核苷,核苷酸及類似物、有機酸及其衍生物、有機含氧化合物、有機雜環化合物、糖類多酮類化合物、生物堿及其衍生物、含氮有機物、有機硫化合物、有機磷化合物和未分類代謝物。分類信息表明脂類及類脂分子和有機酸及其衍生物這2類代謝物數量占比均較高,在上調差異代謝物中分別占19.87%和21.79%,在下調差異代謝物中分別占41.07%和13.1%(圖4)。
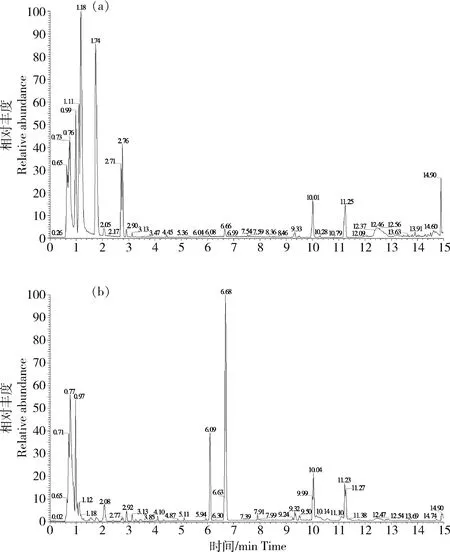
圖1 正離子(a)和負離子(b)模式下肝臟代謝物基峰強度色譜圖Fig.1 Base peak intensity chromatographs of liver metabolites obtained in ES+mode (a) and ES—mode (b)

(a)所有樣本PCA模型得分圖;(b)FT組和TT組PCA模型得分圖;(c)FT組和TT組OPLS-DA模型得分圖;(d)FT組和TT組排列測試圖。(a) PCA score plot for the data from QC samples and all groups; (b) PCA score plot for the data in FT group and TT group; (c) OPLS-DA score plot in FT group and TT group; (d) Permutation test plot in FT group and TT group.圖2 多元統計分析Fig.2 Multivariate statistical analysis
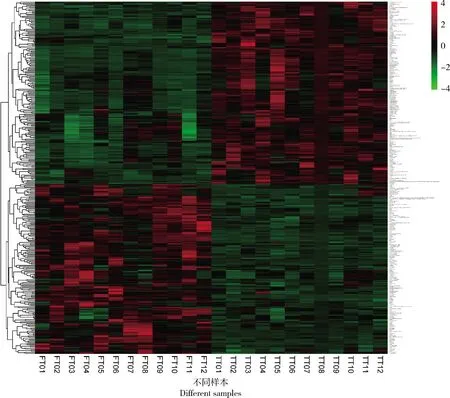
變化程度由不同顏色標記,紅色或藍色分別表示代謝物相對增加或減少的水平。每列代表1個單獨樣品,每行代表1個差異代謝物。TT組為開產前22周齡番鴨,FT組為產蛋期40周齡番鴨。The degree of change is marked by different colors, and red or blue represents the relatively increased or decreased levels of the metabolites, respectively. Each column represents an individual sample, and each row represents a differential metabolite. TT group represents Muscovy duck in pre-laying stage (22 wk), and FT group represents Muscovy duck in laying stage (40 wk).圖3 番鴨產蛋前期和產蛋期肝臟差異代謝物熱圖分析Fig.3 Heat map analysis of hepatic differential metabolites of Muscovy duck in pre-laying and laying stages

圖4 番鴨產蛋前期和產蛋期肝臟差異代謝物分類Fig.4 Classification of hepatic differential metabolites of Muscovy duck in pre-laying and laying stages
2.4 差異代謝物通路富集
通路富集分析可獲得46種差異代謝物的17個通路存在顯著差異(P<0.05)。通路富集表明,氨基酸代謝通路最為豐富,分別為甘氨酸,絲氨酸與蘇氨酸代謝、氨基酸生物合成、氨酰-tRNA生物合成、組氨酸代謝、β-丙氨酸代謝、賴氨酸降解、精氨酸生物合成、精氨酸與脯氨酸代謝和半胱氨酸與蛋氨酸代謝。其中甘氨酸,絲氨酸與蘇氨酸代謝和氨基酸生物合成富集到的差異代謝物數量最多(圖5)。除葉酸一碳庫、半胱氨酸與蛋氨酸代謝、嘌呤代謝、賴氨酸降解和谷胱甘肽代謝通路下調外,其他12個通路均表現為顯著上調(表1)。
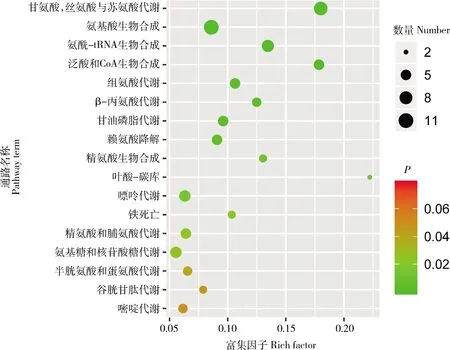
圖5 番鴨產蛋前期和產蛋期差異代謝物KEGG通路富集分析Fig.5 KEGG pathway enrichment analysis of differential metabolites of Muscovy duck in pre-laying and laying stages
分析差異代謝物富集通路,發現谷胱甘肽、半胱氨酸和膽堿等在產蛋前后肝臟組織中代謝差異顯著。谷胱甘肽和半胱氨酸在FT組豐度顯著下調,膽堿在FT組豐度顯著上調,其log2FC分別為-1.48、-1.86和0.64(圖6)。谷胱甘肽參與鐵死亡、半胱氨酸和蛋氨酸代謝和谷胱甘肽代謝;半胱氨酸參與甘氨酸,絲氨酸與蘇氨酸代謝、氨基酸生物合成、氨酰-tRNA生物合成、泛酸與CoA生物合成、鐵死亡、半胱氨酸與蛋氨酸代謝和谷胱甘肽代謝;膽堿參與甘氨酸,絲氨酸與蘇氨酸代謝和甘油磷脂代謝(表1)。
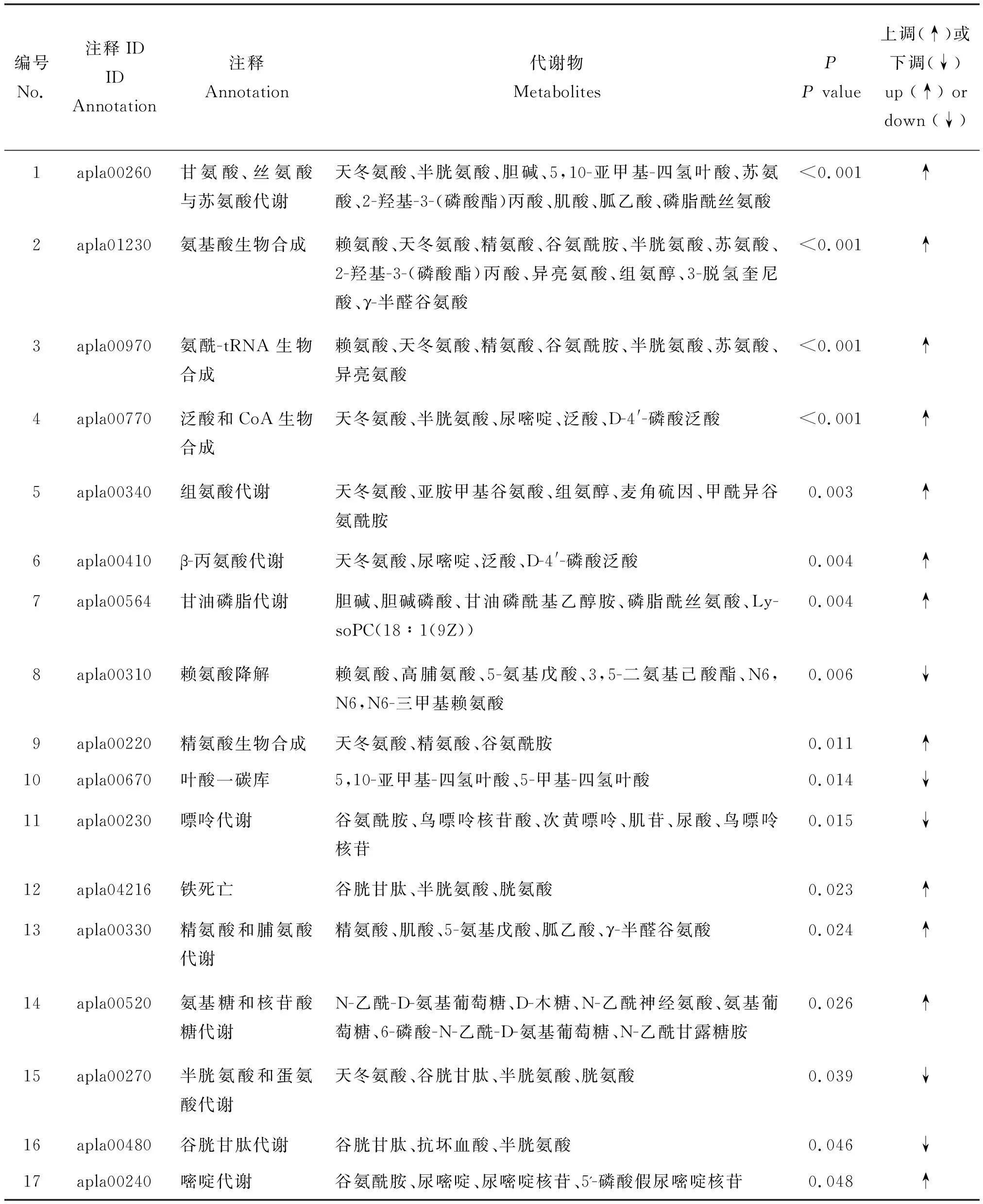
表1 番鴨產蛋前期和產蛋期KEGG通路富集分析Table 1 Enrichment analysis of KEGG pathways of Muscovy duck in pre-laying and laying stages
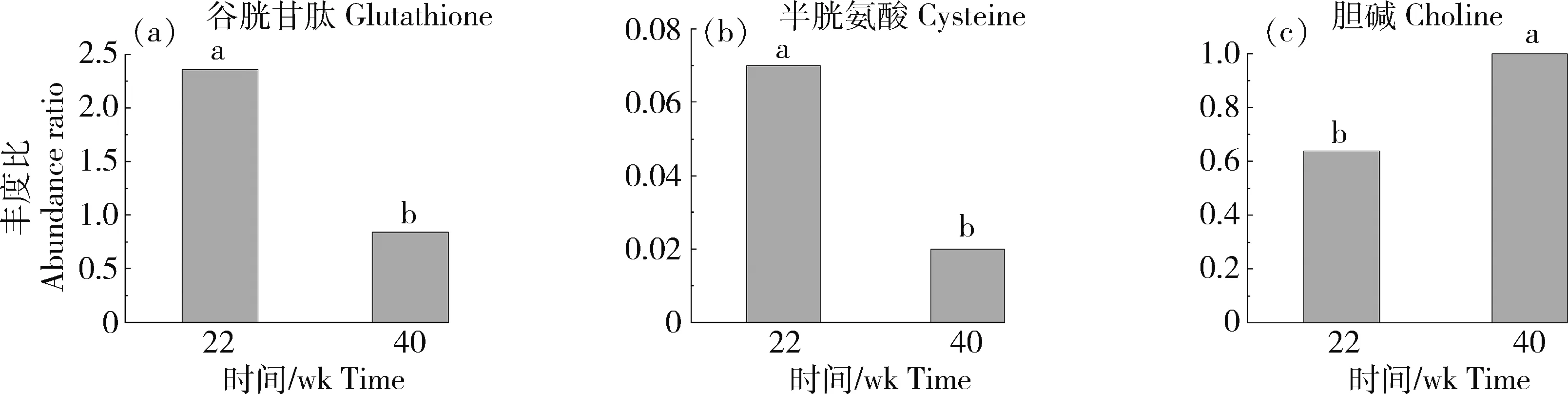
柱上不同字母表示組間差異顯著(P<0.05)Columns marked with different letters represent significant difference (P<0.05) within groups.圖6 番鴨產蛋前期和產蛋期關鍵差異代謝物豐度比Fig.6 Abundance ratio of key differential metabolites of Muscovy duck in pre-laying and laying stages
3 討 論
3.1 番鴨產蛋前后肝臟差異代謝譜分析
甘油磷脂和甘油三酯在分泌早期與載脂蛋白結合,形成富含甘油三酯的脂質蛋白顆粒,在VLDL成熟分泌前進一步引入甘油磷脂[14],組裝成VLDL,成為肝臟分泌的主要脂質成分。產蛋期間,在雌激素作用下每克肝臟每小時可轉錄出1.3×1011mRNA分子,相當于每個肝細胞每分鐘轉錄出11個VTG mRNA分子,并經翻譯后生成大量VTG進入血液[15]。VLDL和VTG為2種主要卵黃前體物,在卵黃形成過程中發揮重要作用,分別構成60%和24%卵黃干物質以及共同構成95%以上的卵黃脂質[16-17]。本研究中,脂類及類脂分子和有機酸及其衍生物數量在FT/TT組占比均較高,可能與VLDL和VTG等卵黃前體合成有關。產蛋為一種復雜的生理過程,從差異代謝物類型可知產蛋涉及到糖代謝、核苷酸代謝、脂代謝、維生素代謝和氨基酸代謝等。
3.2 番鴨產蛋前后肝臟差異代謝通路分析
3.2.1氨基酸代謝通路
3.2.2甘油磷脂代謝
甘油磷脂為機體含量最多的一類磷脂,除構成生物膜外,還是膽汁和膜表面活性物質等成分之一,并參與細胞膜對蛋白質識別和信號傳導[19]。甘油磷脂合成于細胞質滑面內質網,通過高爾基體加工,在VLDL形成初期以及形成前參與其組裝[14,20]。在本研究中,甘油磷脂代謝通路(apla00564)上調,富集到的5種甘油磷脂,除磷脂酰絲氨酸外,膽堿、膽堿磷酸、甘油磷酰基乙醇胺和LysoPC(18∶1(9Z))在FT組豐度顯著增加,可能與肝臟代謝生成大量甘油磷脂以滿足VLDL組裝有關。
3.2.3核苷酸代謝
核苷酸是DNA和RNA的基本組成單位,是體內合成核酸的前體物質。在產蛋期肝臟核苷酸合成增加,GMPS和CMPK1基因顯著上調促進嘧啶合成[21],故嘧啶代謝(apla00240)代謝通路顯著上調。在本研究中,嘌呤代謝(apla00230)通路顯著下調,可能與增加ATP和rRNA合成速率有關[22]。
3.2.4維生素代謝
葉酸為嘌呤和胸腺嘧啶核苷酸合成以及同型半胱氨酸合成蛋氨酸提供一碳單位[23-24]。葉酸不能由動物組織合成,由外源性食物供給并由腸道吸收[25]。本試驗番鴨屠宰前未進食,且肝臟代謝需大量四氫葉酸提供一碳單位,導致葉酸一碳庫代謝通路(apla00670)富集到的2個差異代謝物5,10-亞甲基-四氫葉酸和5-甲基-四氫葉酸在FT組豐度顯著下調。
泛酸(維生素B5)為輔酶A(CoA)前體物,在調節碳水化合物、脂質、蛋白質和核酸代謝等過程中發揮關鍵作用[26]。Youssef等[27]發現飼喂缺乏泛酸飼料的大鼠肝臟中,過氧化物酶體中脂肪酸β-氧化顯著降低。在本研究中泛酸和CoA生物合成通路(apla00770)上調,可能與維持產蛋期間肝臟正常代謝有關。
3.2.5其他代謝
谷胱甘肽代謝通路(apla00480)中,富集到的3種差異代謝物谷胱甘肽、抗壞血酸和半胱氨酸豐度均在FT組顯著下降。谷胱甘肽和抗壞血酸具有抗氧化性,可消除細胞內過氧化物和自由基[28]。40周齡為產蛋中期,番鴨可能適應產蛋應激,故谷胱甘肽和抗壞血酸豐度下降。
鐵死亡為依賴鐵離子及活性氧誘導脂質過氧化導致的調節性細胞壞死,其在形態學、生物學及基因水平上均明顯不同于凋亡、壞死、自噬等其他形式細胞壞死[29]。研究表明鐵死亡生物學特征為鐵和活性氧聚集,激活絲裂原活化蛋白激酶系統,降低胱氨酸攝取和增加谷胱甘肽消耗[30]。在本研究中,鐵死亡代謝通路(apla04216)上調,富集到的差異代謝物谷胱甘肽和半胱氨酸在產蛋期豐度顯著下降,而胱氨酸豐度顯著上調,符合鐵死亡生物學特征,可能與肝臟正常的調節性細胞壞死有關,但相關研究仍需繼續探究。
產蛋期氨基糖和核苷酸糖代謝通路(apla00520)顯著上調,富集到的6種差異代謝物除氨基葡萄糖外,N-乙酰-D-氨基葡萄糖、D-木糖、N-乙酰神經氨酸、6-磷酸-N-乙酰-D-氨基葡萄糖和N-乙酰甘露糖胺豐度在FT組顯著增加。有關核苷酸糖代謝通路中的代謝物在卵黃前體物合成中的作用機理尚無研究報道,有待進一步研究。
3.3 番鴨產蛋前后肝臟關鍵差異代謝物分析
膽堿為蛋雞日糧必需營養物質,是肝臟中磷脂酰乙醇胺(PE)合成磷脂酰膽堿(PC)反應底物[31]。研究表明飼料中添加膽堿能提高母雞抗氧化能力、卵黃總脂和PC以及血清VLDL含量,增強肝臟GSH-Px活性和T-AOC,同時降低肝臟總脂、甘油三酯以及PC含量[32-33]。膽堿在產蛋期豐度顯著上升,可能與肝臟抗氧化應激能力有關,并增加PC含量。PC為VLDL主要成分,VLDL分泌和脂質從肝臟向血液和周圍組織轉運均需膽堿和PC。谷胱甘肽為細胞重要抗氧化劑,由谷氨酸、半胱氨酸和甘氨酸經ATP依賴的谷氨酸-半胱氨酸連接酶和谷胱甘肽合成酶兩步合成,其合成速率受半胱氨酸限制。半胱氨酸參與多個氨基酸代謝通路、鐵死亡、谷胱甘肽代謝和泛酸與CoA生物合成等。有關半胱氨酸在禽類產蛋期作用機理研究報道較少,其在產蛋期豐度顯著下調,可能與產蛋應激適應有關。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
