菊科植物環肽研究進展
2020-11-23 03:52:34楊勇勛
亞太傳統醫藥 2020年11期
楊勇勛,顏 瑜
(1.西昌學院 動物科學學院,四川 西昌 615000;2.西昌學院 圖書館,四川 西昌 615000)
植物環肽(Plant cyclopeptides)一般是指高等植物中由非環肽(直鏈肽)通過酰胺鍵、醚鍵、二硫鍵,或通過一個新的C-C、C-N、N-O或C-S鍵形成的一類環狀含氮化合物。目前發現的植物環肽主要是由2~37個編碼或非編碼氨基酸,且主要是L-構型的氨基酸組成。到2005年,已從分屬26個科、65個屬、120個種的高等植物中分離鑒定出約455個植物環肽,其中大部分的植物環肽來自于石竹科Caryophyllaceae和鼠李科 Rhamnaceae植物[1-2]。
菊科為植物界第一大科,廣布全世界,約有1 000屬,25 000~30 000種,熱帶較少。我國約有200余屬,2 000多種,分布于全國各地[3]。此前一直認為菊科植物環肽只涉及紫菀中的氯化環五肽類,但近期又從菊科其他植物中發現了環肽生物堿以及數十個基因編碼的均環肽,即兩大類環肽:①均環肽:紫菀氯化環五環肽[1,4-6],基因編碼的PDP(PawS-derived Peptides)[7]和PLP(PawL-derived Peptides)環肽[8-9];②雜環肽:絨毛戴星草中的環肽生物堿subfractions Ⅰ和Ⅱ[10-12]。
目前,菊科植物環肽的生物活性、作用機制、生源合成途徑等研究方面取得了長足的進展,如最具有代表性的紫菀環肽astins類化合物具有顯著的細胞毒性,而且其中的astin B活性最強[5]。構效關系研究顯示其細胞毒性與肽鏈是否成環、氯代,環的構象以及肽脯酰胺鍵的順/反式(cis/trans)構象等因素有關[13-15]。研究還顯示astin C具有免疫調節活性,紫菀的止咳、祛痰活性與紫菀環肽類化合物有關[16],PDP環肽SFTI-1(27)具有蛋白酶抑制活性等[7]。
鑒于菊科植物廣布于全世界,且種類較多,因此從中發現種類繁多的新穎結構的生物活性環肽還存有巨大的潛力。此外,近期研究還在菊科植物中環肽的抗腫瘤活性、免疫抑制的作用機制與作用靶點、生源合成途徑、生物技術培養生產等方面取得積極進展。因此,本文對當前菊科植物環肽的來源、分類、合成途徑、化學結構、生物活性、作用機制及構效關系等進行全面的綜述,以期為菊科植物環肽的深入研究與開發提供參考。
1 化學結構
1.1 第Ⅰ類:均環肽
1.1.1 亞類Ⅰ:紫菀環肽 目前,從紫菀根和根莖中共分離得三類環肽類化合物:第1類為氯化環五肽[5,17-20]、astins A-I(1-9)、K-P(10-15);第2類為第1類紫菀氯化環五肽的水解開環直鏈肽[21-23]、aster J(16)和asterinins A-F(17-22);第3類為一類結構新穎的具一個Δ2,4脯氨酸側鏈的環四肽化合物tataricins A-B[6](23-24),氨基酸組成與第Ⅰ類類似。結構見圖1。
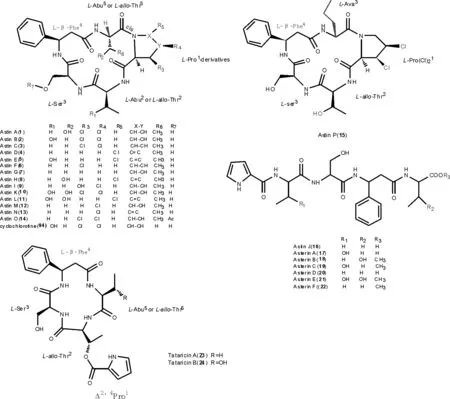
圖1 紫菀環肽astins A-I (1-9),K-P (10-15),astin J,asterins A-F(17-22)和tataricin A-B(23-24)的結構(以脯氨酸為第1氨基酸編號)
1.1.2 亞類Ⅱ:菊科PawS基因編碼的PDPs環肽 此類環肽由12~21個編碼氨基酸頭尾環合而成,含2個半胱氨酸,具有1個二硫鍵。此類環肽首先從向日葵種子中分離得到,如SFTI-1(27)[7,24-26]和SFT-L1 (SFTI-Like1)[27],結構見圖2。隨后,從菊科多種植物中通過LC/MS/MS技術發現多個與SFTI-1(27)、SFTI-L1(28)結構類似的PDPs環肽,部分結構還通過化學合成途徑證實,其具體的名稱、氨基酸序列及來源見表1[28],其中,PDP-10、PDP-11為直鏈肽。
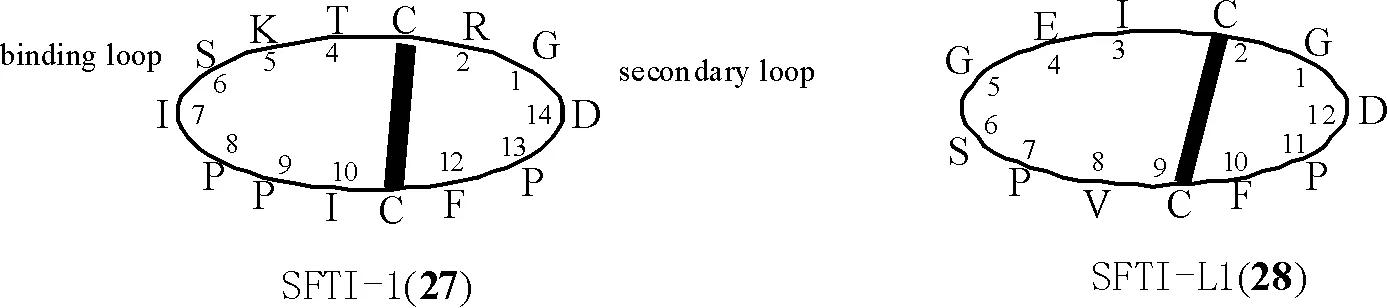
注:文中的編碼氨基酸縮寫表示如下(下同):A=alanine;C =cysteine;D =aspartic acid;E = glutamic acid;F = phenylalanine;G =glycine;H =hystidine;I= isoleucine;K =lysine;L =leucine;M =methionine;N =asparagine;P = proline;Q =glutamine;R =arginine;S =serine;T = threonine;V = valine;W =tryptophan;Y =tyrosine
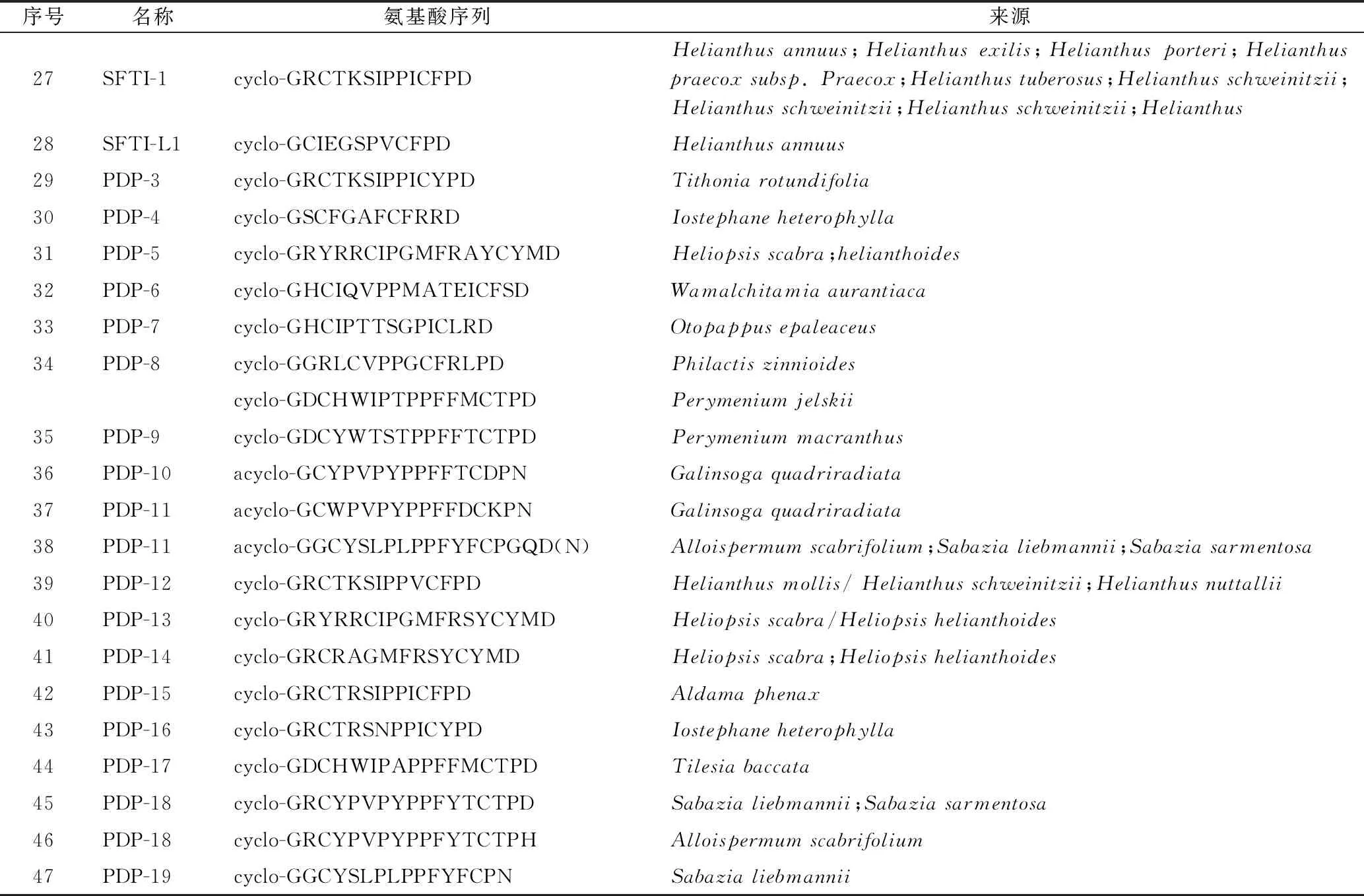
圖2 環肽化合物SFTI-1(27)和SFTI-L1(28)的化學結構(粗實線代表二硫鍵;以甘氨酸為第1氨基酸)
1.1.3 亞類Ⅲ:菊科PawL基因編碼的PLPs環肽 此類環肽由5~12個編碼氨基酸頭尾環合而成,很少含有半胱氨酸(cys),無二硫鍵,如小百日菊(Zinniahaageana)植物中的 PLP-1 (cyclo-AIIPGLID)(48)和千里光屬(Seneciopinnatifoliusvar.maritimus)植物中的PLP-2 (cyclo-DLFVPPID)(49)[8-9]。
到目前為止,共從17種菊科植物中鑒定出46個PLPs型環肽[9]。其具體名稱、氨基酸序列、來源見表2,部分結構見圖3。
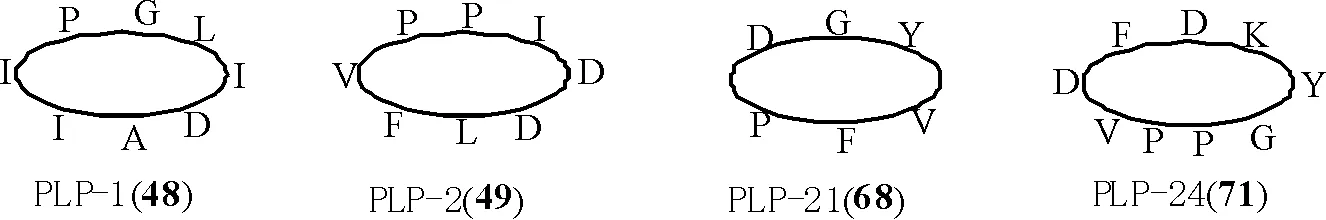
圖3 菊科植物中的部分PLPs型環肽結構
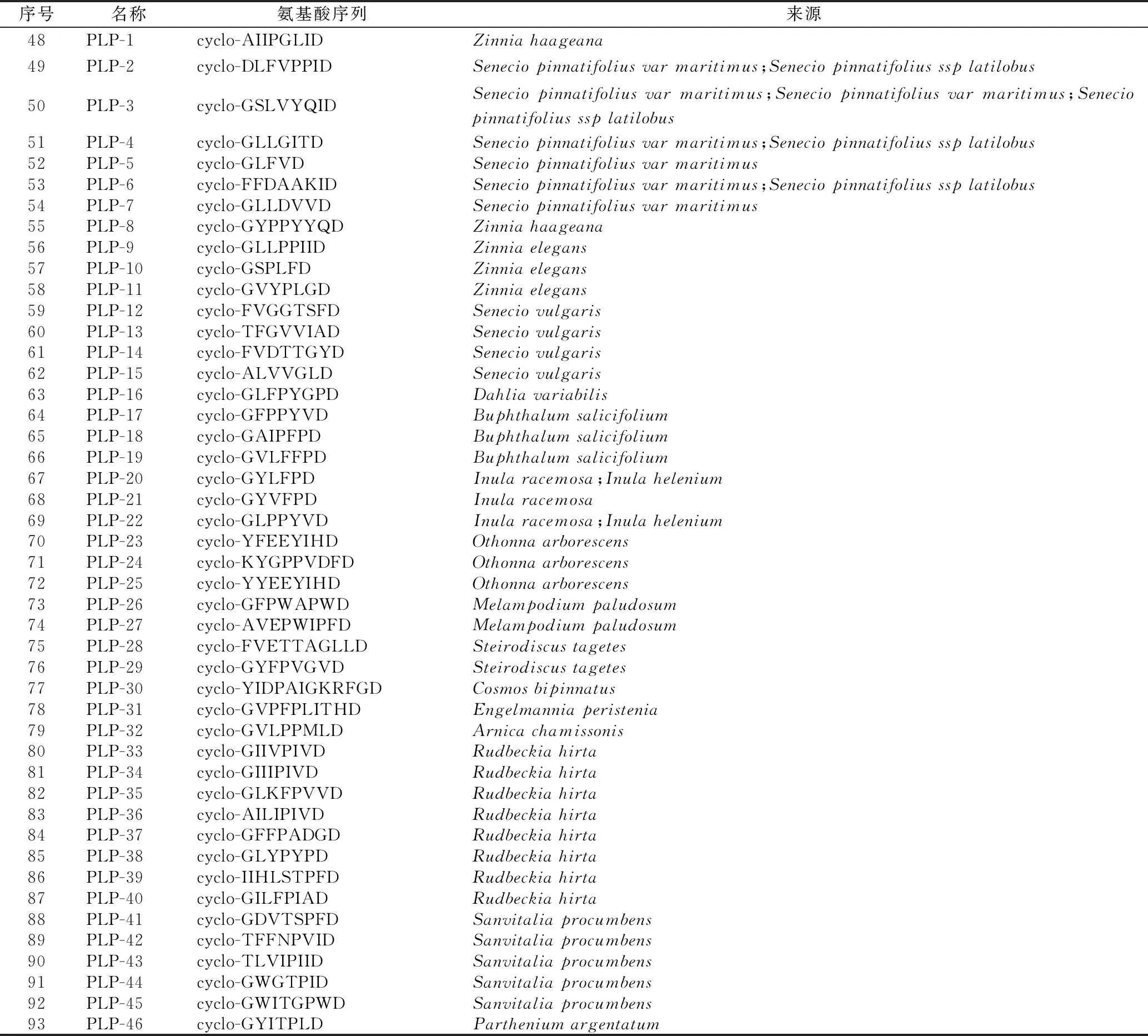
表2 菊科植物中的PLPs環肽
1.2 第Ⅱ類:雜環肽、環肽生物堿、絨毛戴星草環肽
從絨毛戴星草(S.indicus)花[12]中分離得兩個生物堿subfractions Ⅰ和Ⅱ(25-26)[29-30],結構見圖4。
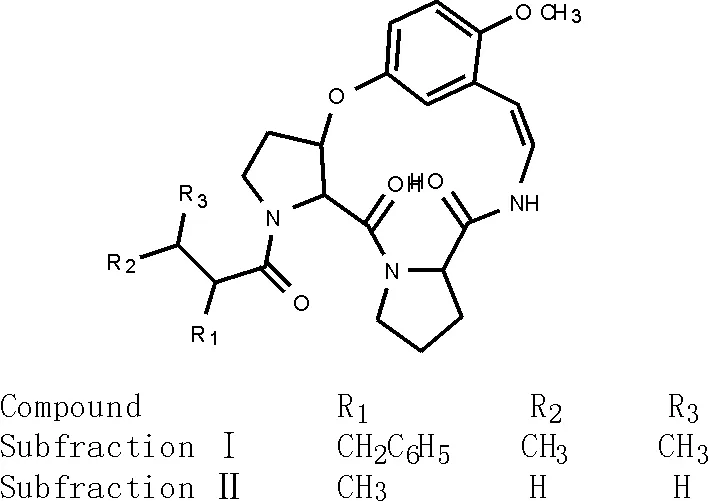
圖4 絨毛戴星草環肽subfractions Ⅰ 和Ⅱ(25-26)的化學結構
2 生物活性
2.1 抗腫瘤活性、構效關系及作用機制
Morita H等[5]通過細胞壓積法(total packed cell volume method),以小鼠腹水肉瘤180(sarcoma 180 ascites)為模型,以腫瘤細胞生長比GR(%)為評價指標,對紫菀環肽astins A-C(1-3)的抗腫瘤活性進行了研究。研究揭示了astins A和B具有顯著的抗腫瘤活性,而且其活性是astinC的10倍。
隨后,更多的研究將目光集中于其構效關系上。研究結果顯示,環肽的細胞毒活性與肽鏈是否成環、環的構象,尤其是1,2-肽脯酰胺鍵的順、反式結構、氯代、硫酰化、羥基數目多少等有關[20,22,31-36]。
紫菀環肽astins的抗腫瘤活性是通過caspase介導的凋亡機制實現,即astins連續地激活caspases 8和 9信號通路,進而相應地激活caspase 3效應器[37]。
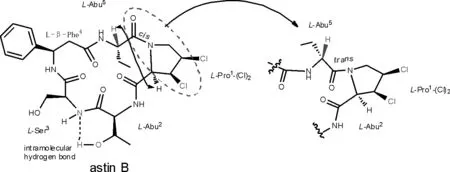
圖5 astin B的結構以及關鍵的NOE相關和分子內氫鍵
2.2 免疫抑制活性、構效關系及作用機制
1990年,Smirnov等[38]首先報道了astin C (原名asterin 3)具有免疫調節作用,其作用機制與T-細胞免疫調節和T-依賴免疫反應相關。
2011年,Shen等[39]報道astin C在體內、體外的免疫活性研究成果:在來自于Fas-mutated MRLlpr/lpr小鼠的激活T細胞中觀察到astin C通過線粒體依賴途徑介導T細胞凋亡;在激活的T細胞中,astin C導致caspase 9和caspase 3裂解,下調Bad蛋白的表達以及細胞色素c的釋放。此外,給小鼠飼喂astin C對抗TNBS-介導的結腸炎體外實驗證實astin C能有效降低血清中TNF-α、IL-4、IL-7水平,誘導活化的T細胞凋亡。實驗證明了astin C能有效抑制激活T細胞的增殖,提示其具有治療結腸炎的潛力。
構效關系研究結果顯示:含D-型氨基酸、親脂長鏈烷基取代、芳基取代的衍生物比具有親水氨基酸和短鏈烷基取代的衍生物活性強。此外,研究結果[40]也顯示,3,4-二氯脯氨酸殘基對astin C發揮免疫抑制作用起到了關鍵作用。
進一步的astin C免疫抑制活性的作用機制研究[41]則顯示,astin C的作用機制是抑制CGAS-STING信號通路,因此astin C可作為調控STING信號通路介導的臨床疾病,如腫瘤、自身免疫性及自身炎癥疾病。
2.3 祛痰、止咳與抗炎活性
YU P等[16]將紫菀根的70%乙醇提取物分為3個流分:Fr-0、Fr-50和Fr-95,采用氣管酚紅分泌法對以上3個流分進行祛痰活性測試,活性最強的部位再采用小鼠酚紅分泌、氨水誘發的咳嗽、二甲苯致耳腫脹實驗評價其祛痰、止咳及抗炎活性。其還采用HPLC-Q-TOF/MS法對活性部位的化學成分進行分析,研究結果顯示紫菀具有止咳、祛痰及抗炎活性,同時也證實了這些活性與紫菀環肽相關。
2.4 蛋白酶抑制活性
SFTI-1(27)具有多種酶抑制活性,其中抑制小牛β-胰蛋白酶的活性最強,為亞納摩爾級別的抑制活性,即其化學計量比為1∶1,解離常數Ki值為 0.1 nmol/L (100 pmol/L)。SFTI-1(27)還具有抑制組織蛋白酶G(cathepsin G) (Ki~<0.15 nmol/L)、彈性蛋白酶(elastase) [Ki~(105±12)μmol/L]、胰凝乳蛋白酶(chymotrypsin)[Ki~(7.4±1.5)μmol/L],以及凝血酶(thrombin) [Ki~(136±21)μmol/L]的活性,但未觀察到對凝血因子Xa (Ki>1 mmol/L)的抑制活性[7]。
2.5 肝毒性
紫菀環肽astins為二氯代脯氨酸環五肽化合物,其與源自于島青霉菌penicilliumislandicum的代謝產物環氯霉毒素cyclochlorotine(94)的結構非常相似,但目前僅報道asterin (astin C,3)具有肝毒性[4]。而研究認為astin B(2)不具有肝毒性的原因是其結構中的肽脯酰胺鍵是順式(cis)構象,而環氯霉毒素(94)中的肽脯酰胺鍵是反式(trans)構象[13](圖5)。但是近年研究證實紫菀環肽,尤其是astin B也具有肝毒性[16-17],因此,紫菀環肽astins的肝毒性應引起重視。
3 紫菀環肽astins的生物合成途徑與生物技術生產
紫菀astins環肽的生物合成途徑尚未闡明,一般認為其是由紫菀植物代謝產生,通過非核糖體肽合成酶(NRPs)合成,因為astins結構中含有非編碼的氨基酸,且NRPSs能夠利用大量非蛋白源(非基因編碼氨基酸)氨基酸底物合成非核糖體肽[42]。然而,源自島青霉菌penicillium islandicum的代謝產物環氯霉毒素cyclochlorotine(94)的結構與astins的結構相似性[43]卻提示astins可能是源自于真菌的代謝產物。
近期有一項研究[44]顯示astins可能源自于真菌:從一株紫菀A.tataricus花序中分離的植物內生真菌Cyanodermellaasteris代謝產生了多個astins類化合物,如asin C(3),但不產生astin A、astin D、astin H等。在未感染植物內生真菌C.asteris的紫菀植物中不會產生astins環肽,而如果紫菀植物再次被感染植物內生真菌C.asteris時又會產生astins環肽,其中包括astin A(1)等,即植物內生真菌C.asteris單獨發酵不產生代謝產物。因此,研究結果提示astin A等astins類環肽既不是單獨由紫菀植物也不是單獨由植物內生真菌產生,而是由植物-真菌二者復合共生交叉代謝產生[45-46]。
鑒于astins環肽在紫菀植物中的含量很低,即astins A-C在植物中的含量分別僅有5.02、2.96和1.03 mg/g[47]。如果采用全合成的方法來合成astins步驟多,合成前體價格昂貴,尤其是二氯脯氨酸不常見,從而阻礙了astins環肽的全合成工業發展。然而,植物內生真菌C.asteris的分離及其代謝產物的分離鑒定使利用生物技術方法大規模生產astins展現出光明的前景。VASSAUX A等[48]采用細胞固定培養技術,在優化的培養條件下,astin C的產量可達植物提取量的5倍(真菌:0.89 mg/g;植物:0.16 mg/g),因此,該生物技術方法生產astin C的效率遠遠超過工業生產的要求。而且由于采用的是固定化培養技術,下游的分離與純化工藝更加便利,成本更低。
4 結語
在2006年以前,紫菀中的astins環肽被歸類為雜環肽類下的菊科型環肽(Compositae-type cyclopeptides),這是由于當時僅從紫菀中發現了氯化環五肽。而現在,其他植物中也發現了環肽生物堿以及幾十個基因編碼的PDPs與PLPs環蛋白(均環肽),因此,繼續將紫菀植物中的astins類氯化環五肽繼續歸類為菊科環肽已似有不妥。
紫菀環肽中的astin B具有顯著的抗腫瘤活性,astin C具有顯著的免疫抑制活性,但不能因為其肽脯酰胺鍵的cis構象與結構類似的真菌代謝產物環氯霉毒素cyclochlorotine(94)的肽脯酰胺鍵的trans構象不同而認為它們不具有肝毒性。具有顯著活性的astin B和C的構效關系、構毒關系以及作用機制研究應得到重視。
此外,基因編碼的PDPs與PLPs環肽在菊科植物中的分布還顯示出分類學上的特征[9,28],因此,它們可能在解決菊科分類的困難上發揮作用。
菊科植物眾多,且大多數為常用中藥,具有廣泛的臨床應用。因此,繼續從中發掘結構新穎的生物活性環肽,并闡明其生物活性、協同增效關系、生物合成途徑等,將會是未來研究的熱點。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
