與RABV感染相關(guān)表觀遺傳修飾酶的篩選
2020-11-13 06:02:12趙銘昕趙維榮徐婧郭藝迪張茂林
山東農(nóng)業(yè)科學(xué) 2020年10期
關(guān)鍵詞:水平
趙銘昕,趙維榮,徐婧,郭藝迪,張茂林
(1.吉林大學(xué)人獸共患病研究所/人獸共患病研究教育部重點(diǎn)實(shí)驗(yàn)室,吉林 長(zhǎng)春 130062;2.吉林大學(xué)動(dòng)物醫(yī)學(xué)學(xué)院,吉林長(zhǎng)春 130062)
狂犬病是由狂犬病病毒(rabies virus,RABV)感染引起的一種嗜神經(jīng)性的人獸共患傳染病,致死率100%[1]。在RABV感染中,不同毒株引發(fā)的臨床表現(xiàn)差異很大,引起了研究者的廣泛關(guān)注[2]。表觀遺傳學(xué)調(diào)節(jié)機(jī)制主要包括DNA甲基化、組蛋白修飾和非編碼RNA作用等。DNA甲基化是目前研究最深入的表觀遺傳修飾方式,涉及的 DNA甲基轉(zhuǎn)移酶主要有 DNMT1、DNMT3a、DNMT3b和DNMT3L。組蛋白表觀遺傳修飾包括甲基化、乙酰化、磷酸化、泛素化等,修飾過(guò)程動(dòng)態(tài)可逆,主要有組蛋白甲基轉(zhuǎn)移酶、組蛋白乙酰轉(zhuǎn)移酶、組蛋白激酶和組蛋白泛素化酶等參與,催化修飾基團(tuán)結(jié)合到組蛋白的氨基殘基。相應(yīng)地,組蛋白去甲基化酶、組蛋白脫乙酰基酶、組蛋白磷酸酶和組蛋白去泛素化酶可去除結(jié)合在組蛋白端氨基殘基上的分子基團(tuán)。非編碼RNA最終使基因沉默從而調(diào)控基因表達(dá)[3-5]。
通常病毒感染會(huì)影響宿主細(xì)胞部分正常生理功能,本質(zhì)上就是宿主基因表達(dá)的修飾,因此病毒自身或引起表觀遺傳學(xué)的變化已成為近年來(lái)的研究熱點(diǎn)[6]。已有研究發(fā)現(xiàn),表觀遺傳修飾酶參與調(diào)控多種病毒感染,例如,位于病毒長(zhǎng)末端重復(fù)序列(LTR)上的組蛋白的去乙酰化和甲基化作用,使?jié)摲腍IV前病毒被沉默[7];乙肝病毒的轉(zhuǎn)錄和復(fù)制過(guò)程依賴于病毒HBx蛋白與 LSD1和Set1A的協(xié)同作用[8];流感病毒NP蛋白在真核細(xì)胞中通過(guò)乙酰化修飾,調(diào)控病毒顆粒的成熟和釋放[9]。已有研究發(fā)現(xiàn),RABV街毒與固定毒株在體內(nèi)感染的細(xì)胞類型不同,而且前者引起病毒基因的轉(zhuǎn)錄水平也遠(yuǎn)遠(yuǎn)低于后者,并將此歸結(jié)為前者逃避機(jī)體免疫的機(jī)制[2]。對(duì)RABV病毒基因組表觀遺傳學(xué)特征進(jìn)行研究,有利于揭示不同病毒株在同類感染介質(zhì)中轉(zhuǎn)錄和復(fù)制水平的變化,有關(guān)RABV的同類研究也在進(jìn)行中[10]。鑒于目前RABV感染的表觀遺傳機(jī)制尚不清楚,本試驗(yàn)通過(guò)對(duì)小鼠神經(jīng)母細(xì)胞瘤(N2a)感染實(shí)驗(yàn)室標(biāo)準(zhǔn)攻擊毒株CVS-11,并在感染后不同時(shí)間點(diǎn)檢測(cè)多種類型表觀遺傳修飾酶基因表達(dá)水平變化情況,篩選參與調(diào)控RABV感染的表觀遺傳修飾酶,以期為不同RABV毒株的可行性研究提供參考。
1 材料與方法
1.1 細(xì)胞和毒株
BHK細(xì)胞和N2a細(xì)胞,均由本實(shí)驗(yàn)室保存;實(shí)驗(yàn)室標(biāo)準(zhǔn)攻擊毒株CVS-11,由長(zhǎng)春獸醫(yī)研究所OIE狂犬病參考實(shí)驗(yàn)室涂長(zhǎng)春研究員惠贈(zèng)。
1.2 主要試劑
細(xì)胞培養(yǎng)基DMEM,購(gòu)自Corning公司;胎牛血清FBS,購(gòu)自Biological Industries公司;Trizol試劑,購(gòu)自TaKaRa公司;反轉(zhuǎn)錄試劑,購(gòu)自Bioteke;熒光定量試劑FastStart Universal SYBR Green Master,購(gòu)自 Roche。
1.3 病毒的培養(yǎng)
將BHK細(xì)胞接種在T25細(xì)胞培養(yǎng)瓶中,待細(xì)胞密度達(dá)到70%時(shí),將CVS-11以MOI=1接種于細(xì)胞瓶中,37℃下感作1 h后,加含1%FBS的DMEM培養(yǎng)基,繼續(xù)培養(yǎng)96 h。-80℃反復(fù)凍融細(xì)胞及培養(yǎng)基3次,收取混合液,將混合液4℃、1 000 r/min離心10 min,收取上清培養(yǎng)基,分裝保存于-80℃。
1.4 病毒的感染及增殖曲線
在六孔細(xì)胞培養(yǎng)板中接種適宜密度的N2a細(xì)胞,37℃過(guò)夜培養(yǎng)后棄掉上清液,將CVS-11按照病毒量MOI=1接種于細(xì)胞,以接種與病毒等體積的磷酸鹽緩沖液(PBS)為對(duì)照。37℃感作1 h,并在感染后 0、3、12、24、48 h提取細(xì)胞總RNA,對(duì)RABV N基因和相關(guān)表觀遺傳修飾酶基因進(jìn)行定量檢測(cè)。
1.5 實(shí)時(shí)熒光定量PCR
采用FastStart Universal SYBR Green Master染料進(jìn)行分析。試驗(yàn)所用RABV N基因,DNA甲基化轉(zhuǎn)移酶 DNMT1、DNMT3a、DNMT3b,組蛋白去乙酰化酶 HDAC1、HDAC2、HDAC3、HDAC5、HDAC6,組蛋白甲基轉(zhuǎn)移酶 EZH2、MLL1、SETDB1、SUV39H1和組蛋白去甲基化酶 KDM6B、PHF8、LSD1以及內(nèi)參GAPDH基因擴(kuò)增引物見(jiàn)表1。
反應(yīng)體系(25μL):上、下游引物各1μL,模板 cDNA 5μL,SYBR Green 12.5μL,ddH2O 5.5 μL,將配好的體系加入96孔板中。反應(yīng)程序:95℃ 10 min,95℃ 15 s,60℃ 1 min,40個(gè)循環(huán)。得到Ct值進(jìn)行統(tǒng)計(jì)分析。
1.6 數(shù)據(jù)統(tǒng)計(jì)與分析
數(shù)據(jù)分析使用GraphPad Prism 5軟件,統(tǒng)計(jì)方法為t檢驗(yàn)。P<0.05有差異(*),P<0.01有顯著差異(**),P<0.001有極顯著差異(***)。

表1 擴(kuò)增表觀遺傳修飾酶基因、RABV N基因與內(nèi)參GAPDH的引物序列
2 結(jié)果與分析
2.1 N2a細(xì)胞上的RABV增殖曲線
為了明確RABV在N2a細(xì)胞上的增殖曲線,將不同時(shí)間點(diǎn)收集并提取的細(xì)胞總RNA反轉(zhuǎn)錄進(jìn)行熒光定量檢測(cè)。結(jié)果表明,RABV感染量隨時(shí)間的推移而增加,并在感染48 h達(dá)到最大值(圖1)。
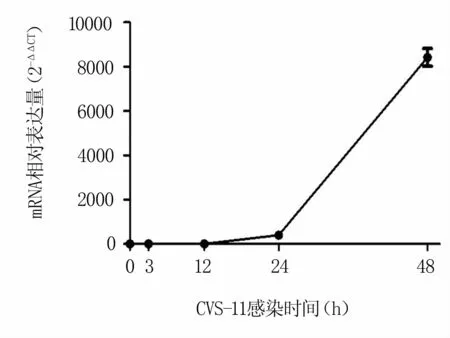
圖1 N2a細(xì)胞上的RABV N基因增殖曲線
2.2 與RABV感染相關(guān)的DNA甲基化轉(zhuǎn)移酶篩選
對(duì)參與甲基化的主要DNA甲基化轉(zhuǎn)移酶進(jìn)行檢測(cè)發(fā)現(xiàn),N2a感染 RABV后,DNMT1、DNMT3a基因表達(dá)水平雖然有變化,但與對(duì)照組(ctrl)相比無(wú)統(tǒng)計(jì)學(xué)差異(圖2 A、B)。DNMT3b基因表達(dá)水平在病毒感染12 h后極顯著下調(diào)(圖2 C)。以上結(jié)果表明,RABV感染抑制DNA甲基化轉(zhuǎn)移酶DNMT3b基因的表達(dá)。
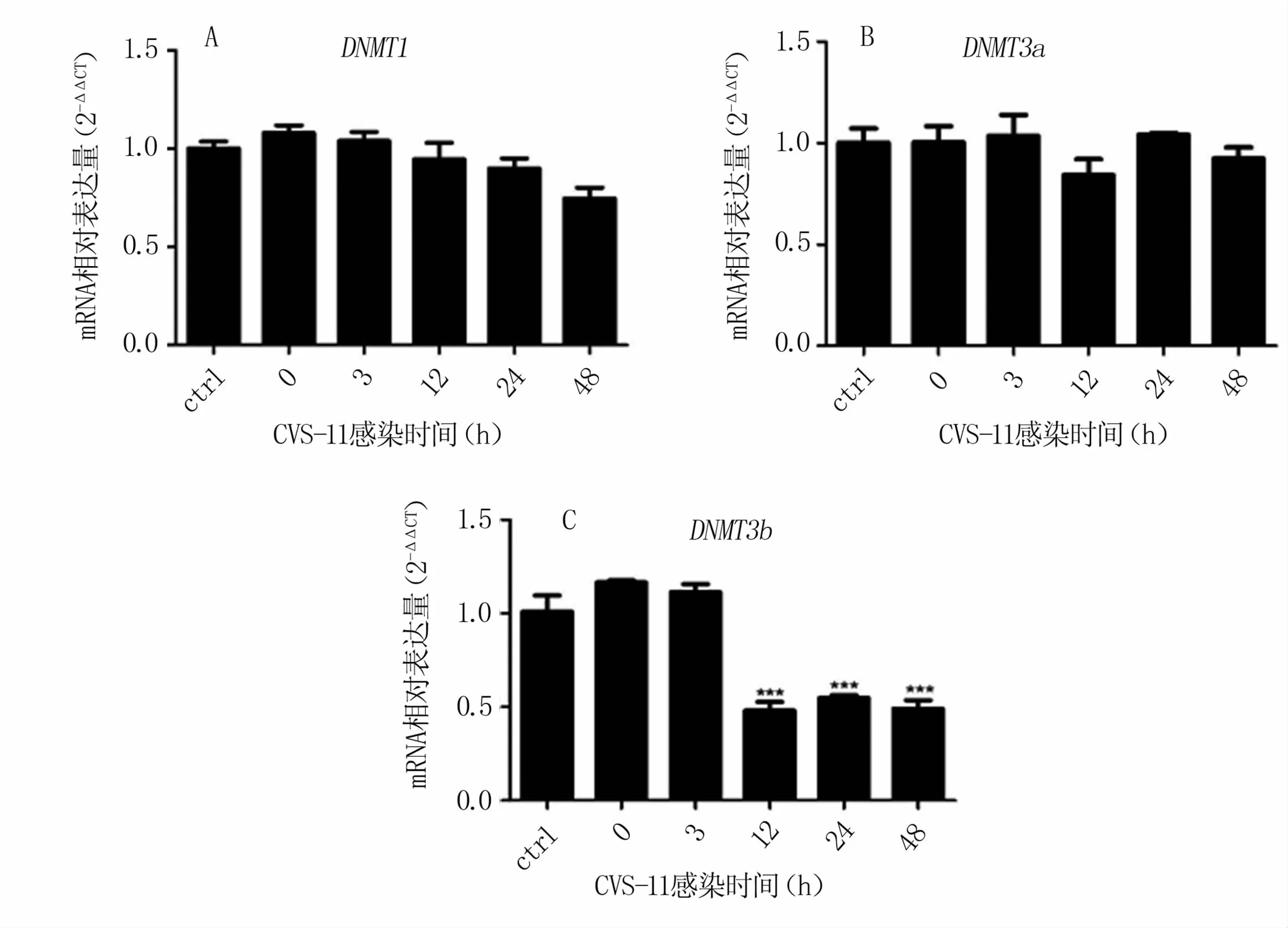
圖2 與RABV感染相關(guān)的DNA甲基化轉(zhuǎn)移酶基因的表達(dá)量
2.3 RABV感染相關(guān)的組蛋白去乙酰化酶篩選
對(duì)參與RABV感染相關(guān)的組蛋白去乙酰化酶進(jìn)行檢測(cè)發(fā)現(xiàn),HDAC2、HDAC5基因表達(dá)水平與對(duì)照組相比無(wú)統(tǒng)計(jì)學(xué)差異(圖3 B、D);HDAC1基因表達(dá)水平24 h開(kāi)始極顯著下降,下降約50%(圖3 A);HDAC3基因表達(dá)水平下調(diào)不明顯(圖3 C);HDAC6基因表達(dá)水平在感染RABV后各時(shí)間點(diǎn)與對(duì)照組相比均極顯著下降,在感染24 h后表達(dá)水平下降50%以上(圖3 E)。以上結(jié)果表明,RABV感染抑制組蛋白去乙酰化酶 HDAC1、HDAC3、HDAC6基因的表達(dá),且對(duì)HDAC6基因表達(dá)水平抑制最為顯著。

圖3 與RABV感染相關(guān)的組蛋白去乙酰化酶基因的表達(dá)量
2.4 與RABV感染相關(guān)的組蛋白甲基轉(zhuǎn)移酶、去甲基化酶篩選
N2a感染 RABV后,組蛋白去甲基化酶KDM6B、LSD1基因表達(dá)水平與對(duì)照組相比無(wú)統(tǒng)計(jì)學(xué)差異(圖4 A、C);PHF8在3、12、24、48 h的表達(dá)水平約下調(diào)至對(duì)照組的60%以下,差異極顯著(圖4 B)。
組蛋白甲基轉(zhuǎn)移酶MLL1基因表達(dá)水平在感染12 h后呈上升趨勢(shì),但與對(duì)照組相比無(wú)統(tǒng)計(jì)學(xué)差異(圖4 E);EZH2基因表達(dá)水平在感染后出現(xiàn)降低趨勢(shì),48 h后約降低至對(duì)照組的60%(圖4 D);SETDB1基因表達(dá)水平在病毒感染12 h后顯著下調(diào),并在感染24 h后下降極顯著,只占對(duì)照組的14%左右(圖4 F);SUV39H1基因表達(dá)水平在病毒感染3 h后極顯著降低,48 h后約降低至對(duì)照組的30%(圖4 G)。
以上結(jié)果表明,RABV感染顯著下調(diào)組蛋白去甲基化酶PHF8和組蛋白甲基轉(zhuǎn)移酶SUV39H1基因的表達(dá),而組蛋白甲基轉(zhuǎn)移酶SETDB1基因表達(dá)在RABV感染晚期被顯著抑制。
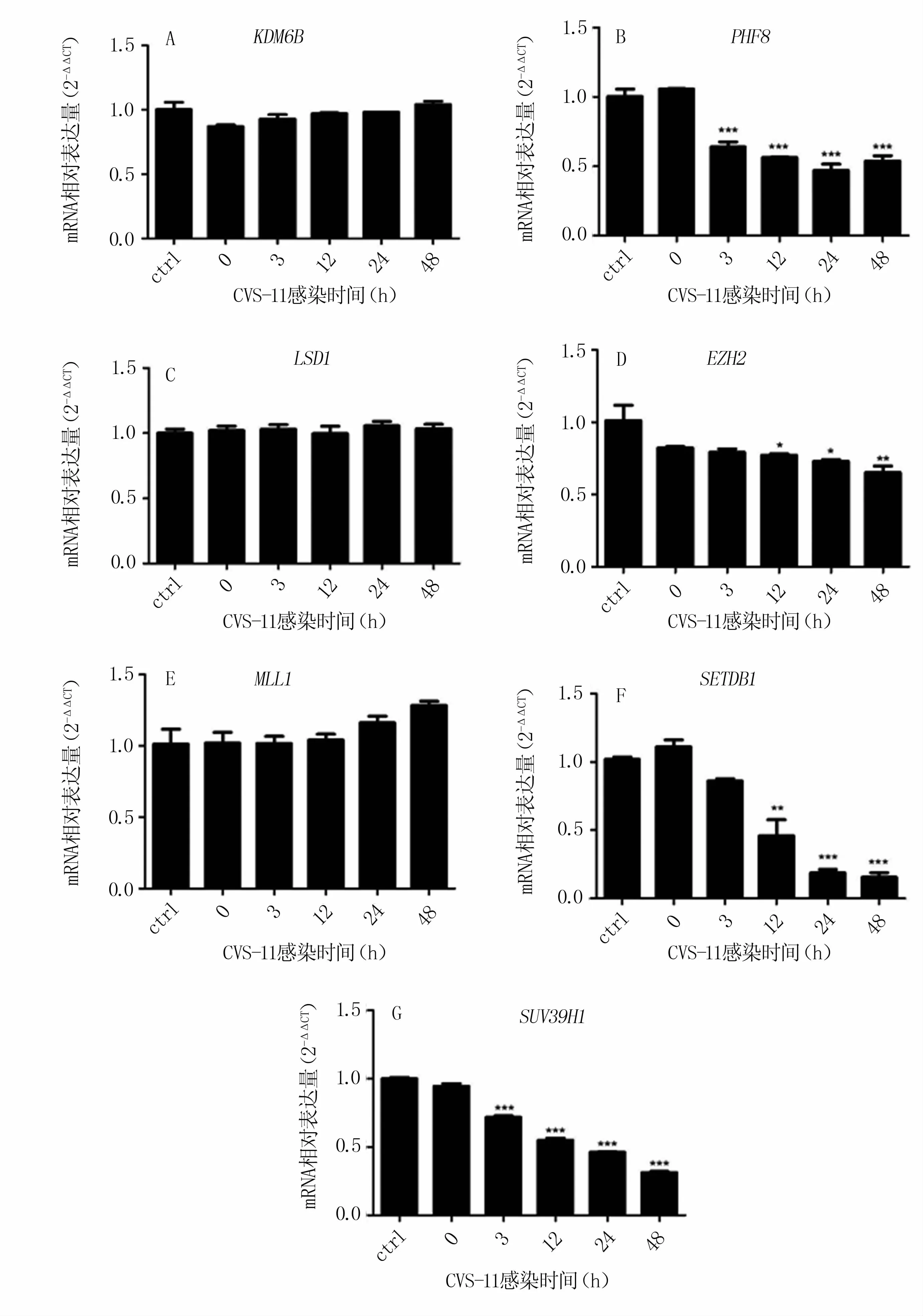
圖4 與RABV感染相關(guān)的組蛋白甲基轉(zhuǎn)移酶、去甲基化酶基因的表達(dá)量
3 討論與結(jié)論
本研究通過(guò)對(duì)小鼠神經(jīng)母細(xì)胞瘤(N2a)接種CVS-11,探討表觀遺傳修飾系統(tǒng)在該毒株感染神經(jīng)細(xì)胞中的作用,從中篩選出與RABV感染相關(guān)且變化顯著的表觀遺傳修飾酶,以期從表觀遺傳修飾水平解析RABV引起臨床表現(xiàn)的致病機(jī)制。
已有研究表明,HIV直接感染調(diào)節(jié)性T細(xì)胞(regulatory T cells,Tregs),誘導(dǎo) DNMT3b的表達(dá)增加,造成轉(zhuǎn)錄因子Foxp3高度甲基化,破壞宿主Tregs的免疫調(diào)節(jié)功能[11]。本研究發(fā)現(xiàn),甲基化轉(zhuǎn)移酶DNMT3b在RABV感染12 h后表達(dá)水平降低,說(shuō)明RABV可能通過(guò)調(diào)節(jié)DNMT3b表達(dá)水平影響靶基因DNA甲基化以利于RABV感染細(xì)胞,具體機(jī)制仍有待研究。甲型流感病毒NP蛋白的超乙酰化和乙酰化缺失均影響甲型流感病毒的復(fù)制[9]。過(guò)表達(dá)HDAC6可導(dǎo)致乙酰化微管蛋白減少及病毒 -細(xì)胞融合減少[12]。本研究中,HDAC6表達(dá)水平在RABV感染0 h與對(duì)照組相比呈現(xiàn)極顯著降低的趨勢(shì),HDAC6可能通過(guò)乙酰化微管蛋白調(diào)節(jié)微管穩(wěn)定性進(jìn)而影響病毒侵入、胞內(nèi)運(yùn)輸、復(fù)制、出芽等感染過(guò)程。組蛋白甲基化修飾是動(dòng)態(tài)可逆的,組蛋白甲基轉(zhuǎn)移酶和組蛋白去甲基化酶在病毒感染過(guò)程中可以協(xié)同發(fā)揮調(diào)節(jié)作用,例如HPV的E6蛋白可以和組蛋白甲基轉(zhuǎn)移酶相互作用并通過(guò)降低酶活性使抑癌基因P53轉(zhuǎn)錄水平降低,E7蛋白能夠催化組蛋白去甲基酶聚集到TLR9啟動(dòng)子區(qū)域最終降低TLR9的功能[13,14]。本研究發(fā)現(xiàn),RABV感染影響了組蛋白去甲基化酶PHF8和組蛋白甲基轉(zhuǎn)移酶EZH2、SETDB1、SUV39H1基因表達(dá)水平,說(shuō)明在RABV感染過(guò)程中,組蛋白甲基化酶和組蛋白去甲基化酶可能協(xié)同調(diào)控下游靶基因的表達(dá)。
在病毒感染過(guò)程中,可能存在多種機(jī)制利用表觀遺傳系統(tǒng)調(diào)節(jié)生命周期中的生物活性。本研究?jī)H針對(duì)RABV感染中的表觀遺傳修飾酶進(jìn)行初步篩選,所調(diào)控的下游靶基因及具體調(diào)控機(jī)制還有待進(jìn)一步研究。
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問(wèn)題研究(2012年1期)2012-03-25 09:54:45
中國(guó)火炬(2010年12期)2010-07-25 13:26:22
中國(guó)火炬(2010年8期)2010-07-25 11:34:30
