野生木黃耆種子休眠解除與萌發研究
2020-11-09 01:44:26李曉梅趙曉英
防護林科技 2020年9期
關鍵詞:物理
李曉梅,趙曉英
(1.山東省濰坊第七中學,山東 濰坊 261021;2.新疆師范大學生命科學學院,新疆特殊環境物種保護與調控生物學實驗室,新疆 烏魯木齊 830054)
種子萌發是植物生活史的一個關鍵環節,是種群更新和植被構建的基礎。種子萌發對溫度、水分等生態因子的響應反映了其適應局地環境的機制[1]。有活力的種子在適宜條件下不萌發的現象叫種子休眠,冷荒漠地區96%的灌木種子具休眠特性[2]。種子休眠是種子植物長期適應原產地生長環境所形成的遺傳特性。從生態學角度來說,休眠是調節種子在最佳時間萌發,避免在不利條件下萌發或之后幼苗面臨不利條件的一種機制。因為一種植物如果只依靠種子進行種群的繁殖更新,那么該植物在某一地域的持久性則取決于種子的萌發和幼苗建成。由于種皮(有時也包括果皮)不透水而造成的休眠為物理休眠,具物理休眠的種子也稱硬實種子[3]。物理休眠在豆科植物中最為普遍。解除物理休眠的方法主要有物理的、化學的和生物的方法。物理方法主要通過機械、低溫、高溫、干濕交錯、電離輻射、超聲波和高壓等方法處理種子;化學方法主要通過酸蝕處理、堿液浸泡,還有其他一些化學物質處理也可以促進硬實種子的吸脹萌發[3]。
種子的物理休眠及解除會隨貯藏時間的變化而變化[4, 5]。隨著貯藏時間的延長,有些植物種子的物理休眠得以解除,但種子也會產生不同程度的劣變, 最終會導致種子活力下降[6,7]。有關膜莢黃耆(Astragalusmembranaceus)及其變種蒙古黃耆(A.membranaceusvar.mongholicus)[8-13]、草木樨狀黃耆(A.melilotoide)[14]種子休眠和解除的研究已有報道。貯藏后種子休眠的解除因種而異,花棒(Hedysarumscorparium)貯藏9個月后硬實率未發生顯著變化[15],砂生槐(Sophoramoorcroftiana)貯藏1年后硬實率減少了86%[16],刺葉錦雞兒(Caraganaacanthophylla)貯藏1年、6年、8年的種子萌發率分別比成熟鮮種子的提高了13%、27%和24%,可見部分種子休眠在貯藏中得以解除,但時間過長萌發率反而下降[17]。
木黃耆為豆科黃耆屬灌木,是黃耆屬為數不多的木本植物之一,生長于石質山坡、常見于小蓬(Nanophytonerinaceum)荒漠草地[18],是良好的飼用植物,也是潛在的植被恢復先鋒植物。本文擬探討影響野生木黃耆種子萌發的溫度和水分條件、種子休眠比率、不同處理對種子休眠的解除效果及短期干貯藏和長期干貯藏對其種子休眠解除的作用。研究有助于深入探討該植物在種子階段的適應機制,為退化荒漠草地的修復提供依據。
1 材料與方法
種子采集:木黃耆果實于2007年6月采自烏魯木齊市鯉魚山,采集后置于室內通風處晾干,選取健康飽滿的種子備用。從成熟健康的種子中隨機選取100粒稱質量,8個重復,稱質量選用萬分之一天平,計算種子的千粒質量。
解除休眠處理:2007年11月,將木黃耆種子分2組:放于100 mm的培養皿中濕潤濾紙上,每組50粒,各3次重復。在 5 ℃下層積處理12周[19]后,將種子從5 ℃移至2.2所述的溫度進行萌發試驗;同時將木黃耆種子埋于其原生境10 cm深的土壤中,待翌年3月將種子取回移至2.2所述進行萌發試驗。2007年12月,將木黃耆種子用砂紙擦破種皮后置于4個變溫進行萌發試驗。
貯藏:將晾干1周的成熟鮮種子裝入磨口玻璃瓶中,室溫下貯藏。分別將鮮種子、貯藏4個月、貯藏10年的種子進行萌發。
萌發試驗:將種子置于100 mm的培養皿中,每組50粒,各3個重復,在光照培養箱(MGC-250)中進行萌發。培養箱溫度為15/5 ℃、20/10 ℃、25/15 ℃及30/20 ℃(晝12 h/夜12 h),這4個溫度接近木黃耆原生境生長季的月平均最高/最低溫度,光照強度為8 000 lx。通過每天在培育皿內濾紙上加水,保持濾紙濕潤。萌發過程以長出2 mm胚根視為萌發,每天檢測1次,并將已萌發的種子撿出。試驗結束后,對未萌發的種子用TTC法檢測生活力。萌發率為萌發種子占有生活力種子的百分率[19]。試驗持續30d。
數據處理:利用SPSS22.0統計分析軟件對所測數據進行處理分析,用單因素方差分析法在95%水平上檢驗種子的萌發率,差異顯著性檢驗時數據做反正弦轉換[19]。試驗結果以平均值(Mean)±標準誤(SE)表示。
2 結果與分析
木黃耆種子的千粒質量為3.089 5 g。由圖1可知,成熟鮮種子在15/5 ℃、20/10 ℃、25/15 ℃及30/20 ℃(晝12 h/夜12 h)4個溫度的萌發率分別為26%、27%、34%、56%。
2.1 不同處理方法對種子萌發的影響
在同一變溫下,擦破種皮處理的種子萌發率最高(P<0.05),均超過97%。除25/15 ℃變濕外,5 ℃下低溫層積與原生境土壤中層積的萌發率差異不顯著(見圖1)。經兩種方式層積處理的種子萌發率均比未處理的低(P<0.05)。
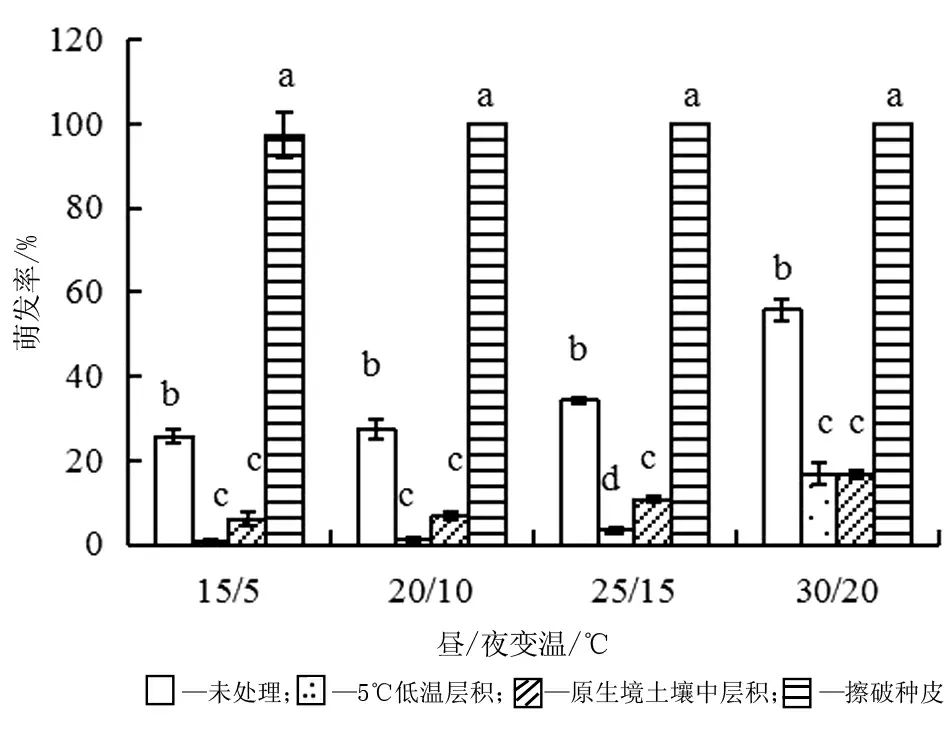
圖1 不同處理方法對種子萌發的影響(平均值±標準誤)
2.2 干貯藏對萌發的影響
由圖2可知,不同貯藏時間的種子在設置的3個變溫條件下的萌發率均沒有顯著性差異。在20/10 ℃和25/15 ℃兩個變溫條件下干貯藏4個月與10年的種子萌發率沒有顯著性差異,鮮種子與貯藏4個月的種子萌發率也沒有顯著性差異。在3個變溫條件下貯藏10年的種子均比鮮種子的萌發率低(P<0.05)。
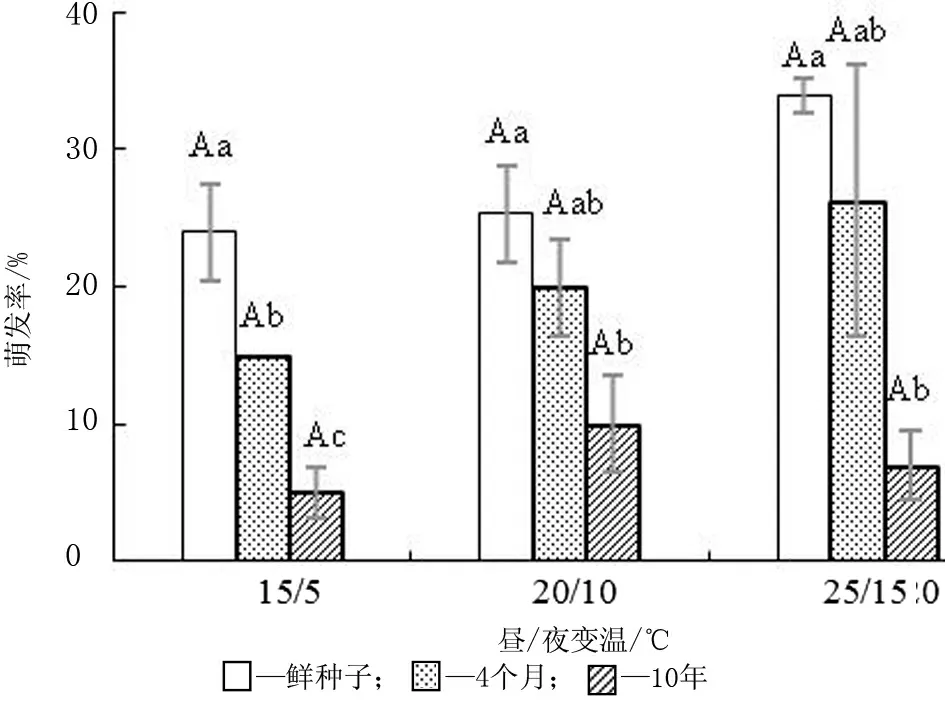
圖2 干貯藏不同時間后種子的萌發率(平均值±標準誤)
3 結論與討論
本研究中,木黃耆種子在15/5 ℃到30/20 ℃的晝夜變溫范圍內均可萌發,最適溫度下的萌發率為54.7%,即成熟鮮種子有45.3%的種子存在休眠,這與蒙古黃耆[11-13]的種子休眠率相近。擦破種皮后,除15/5 ℃變濕外,其他三個較高溫度下的萌發率均達到100%,說明木黃耆種子為物理休眠,即硬實。物理休眠在豆科植物中普遍存在,有利于種子活力的保持。如在室溫下保存的大豆硬實種子保存4年萌發率仍在90%以上,而非硬實種子2年左右的時間即失去生活力[20]; 硬實還能保證種子在成熟、收獲以后的不同時期內,由于環境條件(低溫、變溫、土壤濕度變化和微生物活動等)的作用改變種皮透性,使不同的個體逐漸萌發[21],確保其在適宜條件下有個別種子成功萌發和建苗。低溫層積和土壤中埋放只能解除一小部分種子的休眠。在自然環境中,木黃耆的硬實種子在土壤中可能經歷礫石擦破種皮以及土壤溫度和濕度變化,從而解除休眠。
種子的自然壽命是不同的,即使同種同屬的也不盡相同[22]。刺葉錦雞兒[17]的種子隨著貯藏時間的延長,種子的萌發率越來越高,干貯藏能解除部分種子物理休眠,草原錦雞兒(C.pumila)[17]種子的萌發不受貯藏時間的影響,紫花苜蓿(Medicagosativa)、黃花苜蓿(M.falcata)[4]的種子隨著貯藏時間延長萌發率先升高又降低。木黃耆種子經干貯藏10年后的萌發率顯著低于鮮種子的萌發率,這與砂生槐[16]、串葉松香草(Silphiumperfoliatum)[4]的結果相似,這可能與木黃耆種皮的特殊結構有關,干貯藏提高種子休眠率的機理還有待于進一步的研究。在20/10 ℃和25/15 ℃這兩個溫度條件下,貯藏4個月和10年種子的萌發率差異不顯著,證實了木黃耆種子的壽命很長,使其可以進入持久種子庫,為種群更新提供保障。種子保持長時間休眠是植物對不確定降水和不可預測環境的一種適應[23]。
綜上所述,木黃耆成熟鮮種子在四個溫度下均可萌發,適宜溫度下的硬實率高達44%。擦破種皮解除休眠的效果最好,低溫層積和原生境埋放種子的萌發率比鮮種子顯著降低,不能解除種子的休眠。短期干
猜你喜歡
中學生數理化·八年級物理人教版(2023年11期)2023-12-26 07:50:02
井岡教育(2022年2期)2022-10-14 03:11:44
中學生數理化(高中版.高考理化)(2022年5期)2022-06-01 06:27:46
中學生數理化(高中版.高考理化)(2022年1期)2022-04-26 13:53:42
中學生數理化(高中版.高考理化)(2022年3期)2022-04-26 13:41:26
中學生數理化(高中版.高考理化)(2022年3期)2022-04-26 13:41:24
課堂內外(初中版)(2022年2期)2022-02-28 02:00:30
云南教育·中學教師(2020年9期)2020-11-16 00:27:58
中學生數理化·八年級物理人教版(2019年9期)2019-11-25 07:33:00
中學生數理化·八年級物理人教版(2017年9期)2017-12-20 08:11:28
