國(guó)內(nèi)橡膠樹尖孢炭疽復(fù)合種種間分子系統(tǒng)發(fā)育及生物學(xué)特性分析
2020-10-29 07:35:39江濤劉先寶李博勛鄭肖蘭蔡吉苗馮艷麗黃貴修
熱帶作物學(xué)報(bào) 2020年9期
江濤 劉先寶 李博勛 鄭肖蘭 蔡吉苗 馮艷麗 黃貴修

摘 ?要:為明確國(guó)內(nèi)橡膠樹尖孢炭疽復(fù)合種內(nèi)不同種的生物學(xué)特性及種間差異情況,評(píng)價(jià)了4個(gè)代表性菌株(Colletotrichum bannanense YNML52,C. laticiphilum YNJH17109,C. australisinense GX1655,YNJP162)對(duì)不同碳源、氮源的利用情況,以及在不同溫度、pH、水活度下的生長(zhǎng)情況,并評(píng)價(jià)了這些菌株對(duì)橡膠不同品種和不同作物的致病性。結(jié)果表明:供試菌株在碳源、氮源利用,適宜生長(zhǎng)溫度、致死溫度、pH和水分活度等方面均存在顯著差異;致病性分析表明,供試菌株的致病譜也存在顯著差異,YNML52的寄主范圍較窄,具有很強(qiáng)的寄主專化性。通過(guò)評(píng)價(jià)生態(tài)因子對(duì)橡膠樹炭疽病病原菌生物學(xué)特性的影響,為深入了解橡膠病害發(fā)生規(guī)律和制定有效防控措施提供指導(dǎo)依據(jù)。
關(guān)鍵詞:橡膠樹;尖孢炭疽復(fù)合種;生物學(xué)特性;致病性
中圖分類號(hào):S763.7;S794.1 ? ? ?文獻(xiàn)標(biāo)識(shí)碼:A
Abstract: In order to clarify the biological characteristics and interspecific differences of Colletotrichum acutatum complex species from rubber trees in China, four representative strains (C. bannanense YNML52, C. laticiphilum YNJH17109, C. australisinense GX1655, YNJP162) were evaluated for the utilization of different carbon, nitrogen sources and colony growth at different temperature, pH and water activities. The pathogenicity of the strains to different rubber varieties and crops was also evaluated. There were significant differences in the utilization of carbon and nitrogen sources, suitable growth temperature, lethal temperature, pH value and water activity of the tested strains. There were significant differences in pathogenicity spectrum of the tested strains. YNML52 had a narrow host range and strong host specificity. Evaluating the effects of ecological factors on the biological characteristics of the pathogen of rubber tree anthracnose is of certain guiding significance for understanding the occurrence regularity of the disease and formulating effective control strategies.
Keywords: rubber trees; Colletotrichum acutatum complex species; biological characteristics; pathogenicity
DOI: 10.3969/j.issn.1000-2561.2020.09.019
巴西橡膠樹[Hevea brasiliensis (Willd. ex A. Juss) Muell. Arg]屬大戟目大戟科橡膠樹屬的多年生高大落葉喬木,因相較于其他產(chǎn)膠植物具有不可媲美的優(yōu)越特性,得以在中國(guó)和世界其他適宜地區(qū)廣泛種植[1]。一直以來(lái),病害是制約我國(guó)天然橡膠生產(chǎn)的重要因子,由刺盤孢屬(Colleto??tri?chum spp.)侵染引起的橡膠樹炭疽病正是其中最為嚴(yán)重的葉部病害之一[2]。國(guó)內(nèi)該病最早于1962年在海南大豐農(nóng)場(chǎng)開割林段爆發(fā)流行,引起橡膠樹嚴(yán)重落葉、枝條枯死,開割時(shí)間推遲導(dǎo)致干膠產(chǎn)量嚴(yán)重?fù)p失[3]。
該病病原早期一直被鑒定為膠孢炭疽(C. gloeosporioides)。1992年,在印度尼西亞和斯里蘭卡首次報(bào)道了由尖孢炭疽(C. acutatum)引起的炭疽病,在印度和斯里蘭卡等國(guó)家認(rèn)為尖孢炭疽是優(yōu)勢(shì)致病種群;我國(guó)直到2008年才在西雙版納墾區(qū)首次發(fā)現(xiàn)尖孢炭疽為害[4-5]。最新的分類學(xué)研究將刺盤孢屬劃分為13個(gè)復(fù)合種,與橡膠樹相關(guān)的有膠孢炭疽、尖孢炭疽和博寧炭疽3個(gè)復(fù)合種[6-7]。已有研究表明國(guó)內(nèi)橡膠樹炭疽病原主要屬于膠孢炭疽和尖孢炭疽兩大復(fù)合種,其中膠孢炭疽復(fù)合種包括暹羅炭疽(C. siamense)、果生刺盤孢(C. fructicola)和樂(lè)東炭疽(C. ledongense)3個(gè)種,尖孢炭疽復(fù)合種包括版納炭疽(C. ban-nanense)、華南炭疽(C. australisinense)和C. laticiphilum 3個(gè)種[7-10]。據(jù)報(bào)道,復(fù)合種內(nèi)在種群比例、分布范圍、為害癥狀和為害程度等方面存在明顯差異,導(dǎo)致這一現(xiàn)象的原因可能與各種間不同的生物學(xué)特性有關(guān)[4, 7, 11]。然而,先前文獻(xiàn)報(bào)道的關(guān)于橡膠樹炭疽菌生物學(xué)特性的研究還是以傳統(tǒng)方法鑒定的膠孢炭疽和尖孢炭疽為研究對(duì)象,且多見(jiàn)于膠孢炭疽。在當(dāng)前的分類水平下,其相關(guān)結(jié)果顯得滯后且不夠準(zhǔn)確。
本研究旨在進(jìn)一步明確國(guó)內(nèi)橡膠樹尖孢炭疽復(fù)合種內(nèi)各種間的生物學(xué)特性,以期為橡膠樹炭疽病病原檢測(cè)和制定有效防治策略提供參考。
1 ?材料與方法
1.1 ?材料
1.1.1 ?供試菌株與植物品種 ?供試菌株:橡膠樹尖孢炭疽菌株為本研究室已鑒定、發(fā)表的YNML52、GX1655、YNJP162、YNJH17109,于2017年在云南新收集的菌株。供試植物(品種):番木瓜(‘蜜紅)、芒果(‘金煌芒)、番石榴(‘新世紀(jì))和辣椒(‘辣椒309)。
1.1.2 ?供試培養(yǎng)基、試劑和儀器 ?供試培養(yǎng)基:PDA培養(yǎng)基參照《植病研究方法》配制[12],Czapek培養(yǎng)基參照《植物病理學(xué)實(shí)驗(yàn)技術(shù)》配制[13]。試劑:CTAB、10?Biffer、dNTPs、Taq酶、引物對(duì)、DNA Marker等。儀器:PCR儀、電泳儀、凝膠成像系統(tǒng)、真菌培養(yǎng)箱、超凈工作臺(tái)等。
1.2 ?方法
1.2.1 ?橡膠尖孢炭疽復(fù)合種種間分子系統(tǒng)發(fā)育分析 ?DNA提取、PCR擴(kuò)增和測(cè)序:參照Liu等[7]的方法獲得供試菌株基因組DNA并擴(kuò)增內(nèi)轉(zhuǎn)錄間隔區(qū)序列(ITS)、β-微管蛋白2(TUB2)、肌動(dòng)蛋白(Act)、甘油醛-3-磷酸脫氫酶(GAPDH)和幾丁質(zhì)酶(CHS-1)等5個(gè)基因序列。
系統(tǒng)發(fā)育分析:參考最新的尖孢炭疽復(fù)合種系統(tǒng)發(fā)育研究,在NCBI網(wǎng)站下載模式菌株和代表菌株序列[7]。使用Mega V6.0軟件手動(dòng)調(diào)整,所有的空白視為數(shù)據(jù)缺失。使用MrModeltest V3.6生成核苷酸替換模型。使用Mega V6.0對(duì)數(shù)據(jù)集進(jìn)行最大簡(jiǎn)約法(maximum parsimony, MP)系統(tǒng)發(fā)育分析,分析尖孢炭疽復(fù)合種種間的系統(tǒng)進(jìn)化關(guān)系,進(jìn)化樹利用Mega V6.0進(jìn)行觀察和編輯。
1.2.2 ?生物學(xué)特性測(cè)定 ?碳、氮源對(duì)供試菌株菌絲生長(zhǎng)的影響:選擇Czapek培養(yǎng)基為基礎(chǔ)培養(yǎng)基。分別用等質(zhì)量的木糖、阿拉伯糖、葡萄糖、果糖、半乳糖、麥芽糖、乳糖、果膠、纖維素、可溶性淀粉、多聚半乳糖醛酸、堿性木素、檸檬酸鉀和酒石酸銨[14]替代Czapek培養(yǎng)基中的蔗糖,配制成不同碳源的培養(yǎng)基,以不含碳源的Czapek培養(yǎng)基為空白對(duì)照。分別用等質(zhì)量的硝酸銨、硝酸鉀、磷酸二氫銨、硫酸銨、氯化銨、蛋白胨和天冬酰胺替換Czapek培養(yǎng)基中的硝酸鈉,配制成不同氮源的培養(yǎng)基,以不含氮源的Czapek培養(yǎng)基為空白對(duì)照。將4株供試菌株在PDA平板上活化7 d后,用滅菌的內(nèi)徑5 mm的打孔器制取菌餅接種于含不同碳、氮源的平板中央,各3次重復(fù)。28 ℃恒溫培養(yǎng)5 d后,采用十字交叉法測(cè)量菌落直徑。
溫度對(duì)供試菌株菌絲生長(zhǎng)的影響:將4株供試菌株菌餅接種于PDA平板中央,各3次重復(fù)。后分別放置在12、16、20、24、28、32、36、40 ℃條件下恒溫培養(yǎng)。5 d后,采用十字交叉法測(cè)量菌落直徑。
pH對(duì)供試菌株菌絲生長(zhǎng)的影響:PDA培養(yǎng)基冷卻至50 ℃左右,加入1 mol/L HCl溶液或1 mol/L NaOH溶液將培養(yǎng)基pH分別調(diào)至4、5、6、7、8、9,制作平板。后接入4株供試菌株菌餅,各3次重復(fù)。28 ℃恒溫培養(yǎng)5 d后,采用十字交叉法測(cè)量菌落直徑。
水活度對(duì)供試菌株菌絲生長(zhǎng)的影響[15]:水活度(Aw)代表微生物生長(zhǎng)過(guò)程中參與生化反應(yīng)的自由水的有效值。在1 L PDA培養(yǎng)基中分別添加0、108、198、245、313 mL甘油,然后加入dH2O定容至1 L制備成不同水活度(1.00、0.99、0.95、0.93、0.90)的培養(yǎng)基(甘油將附著一部分水,使其無(wú)法被病原菌利用),以水活度為1作對(duì)照。后接種菌餅,各3次重復(fù)。28 ℃恒溫培養(yǎng)8 d后,采用十字交叉法測(cè)量菌落直徑。
1.2.3 ?致病性測(cè)定 ?供試菌株對(duì)不同橡膠品種的致病性測(cè)定:采用孢子懸浮液接種離體葉片法測(cè)定。摘取PB86、PB235、PB260、IAN873、GT1、PR107、RRIC600、RRIC100、文昌11、云研77-4、熱研7-33-97共11個(gè)當(dāng)前各省主栽或主推橡膠品種的健康古銅期葉片,先用70%的酒精表面消毒1 min,后用無(wú)菌水清洗3次,晾干備用。取活化培養(yǎng)7 d的平板,用無(wú)菌水洗下孢子,配制成濃度為106 CFU/mL的孢子懸浮液。將處理好的葉片正面放置在鋪有吸水紙的培養(yǎng)皿中,在合適位置(以半邊葉片中間位置最佳)接種20 μL孢子液,各3次重復(fù)。以滴加等量無(wú)菌水為空白對(duì)照。噴灑適量無(wú)菌水于吸水紙上,封口后放置在28 ℃下恒溫培養(yǎng)。每12 h定時(shí)觀察葉片表面是否出現(xiàn)侵染癥狀,記錄發(fā)病時(shí)間,4 d后拍照記錄并測(cè)量病斑大小。
供試菌株對(duì)4種作物的致病性測(cè)定:采用孢子懸浮液接種離體葉片法測(cè)定。孢子懸浮液配制同上。選取番木瓜、芒果、番石榴的健康嫩葉和辣椒葉片及果實(shí),消毒晾干后,設(shè)刺傷和健康2個(gè)處理組,按同樣的方法接種孢子懸浮液,各3次重復(fù)。以無(wú)菌水為對(duì)照。封口后28 ℃下保濕培養(yǎng),4~6 d后觀察、測(cè)量,評(píng)價(jià)供試菌株是否能侵染其他植物。
1.3 ?數(shù)據(jù)處理
采用Excel 2007軟件整理數(shù)據(jù),采用SPSS 17.0軟件進(jìn)行差異顯著性分析。
2 ?結(jié)果與分析
2.1 ?橡膠尖孢炭疽復(fù)合種種間分子系統(tǒng)發(fā)育分析
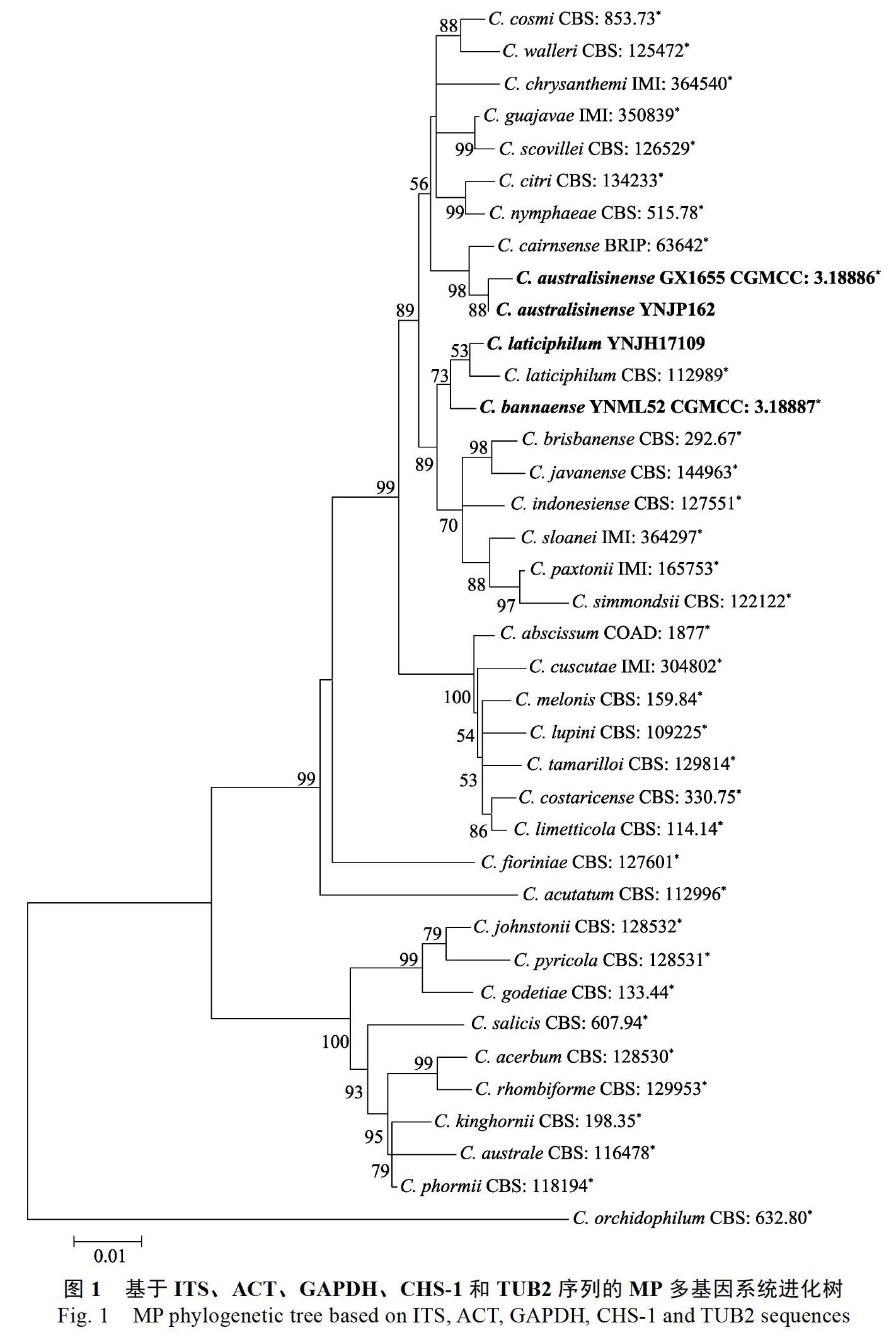
根據(jù)NCBI數(shù)據(jù)庫(kù)中ITS序列的BLAST搜索結(jié)果顯示,YNJH17109與尖孢炭疽復(fù)合種同源性達(dá)99%以上。圖1展示了來(lái)源于橡膠樹的尖孢炭疽(圖中粗體表示)在尖孢炭疽復(fù)合種中的系統(tǒng)發(fā)育關(guān)系。36株菌的基因序列(ITS、TUB2、ACT、GAPDH和CHS-1)串聯(lián)排列,以C. orchidophilum CBS 632.80作為外群。組合序列包含空白共1649個(gè)特征,其中各基因區(qū)間為:ITS:1~478,TUB2:479~961,GAPDH:962~1197,ACT:1198~1417,CHS-1:1418~1649。MRmodeltest v3.6預(yù)測(cè)最大似然樹構(gòu)建最佳模型為GTR+G。利用Mega V6.0構(gòu)建最大似然樹,1000次重復(fù),各分支大于50%的后驗(yàn)概率標(biāo)注于相應(yīng)節(jié)點(diǎn)處。4株來(lái)自橡膠樹的尖孢炭疽分屬于3個(gè)獨(dú)立分支。歸屬于華南炭疽的GX1655和YNJP162與C. cairnsense親緣關(guān)系最近,版納炭疽YNML52與C. laticiphilum聚為姐妹枝,YNJH17109則與C. laticiphilum聚在一個(gè)分支。
2.2 ?不同碳源、氮源對(duì)供試菌株菌絲生長(zhǎng)的影響
2.2.1 ?不同碳源對(duì)菌絲生長(zhǎng)的影響 ?4株尖孢炭疽在供試的5種單糖、3種二糖、5種多糖和2種有機(jī)酸共15種碳源上均有生長(zhǎng),但不同碳源的影響程度有差異(圖2)。半乳糖和乳糖分別是4株尖孢炭疽利用率最低的單糖和二糖;多糖中則果膠的利用率最高,堿性木素最低。其中,YNML52分別在葡萄糖、蔗糖為碳源時(shí)菌落直徑最大,在堿性木素上最小;YNJH17109在半乳糖上最大,分別在葡萄糖、果糖、蔗糖、纖維素、可溶性淀粉和多聚半乳糖醛酸上最小;YNJP162分別在乳糖、果膠、可溶性淀粉、多聚半乳糖醛酸和堿性木素上最大;GX1655分別在木糖、阿拉伯糖上最大;以上差異均達(dá)到顯著水平。此外,YNJP162對(duì)2種有機(jī)酸的利用率極顯著大于另外3個(gè)菌株。
2.2.2 ?不同氮源對(duì)菌絲生長(zhǎng)的影響 ?4株尖孢炭疽在供試的3種硝態(tài)氮、3種銨態(tài)氮和2種有機(jī)氮共8種氮源上均有生長(zhǎng),但存在差異,同類氮源間利用率也存在差異(圖3)。硝態(tài)氮中,YNML52偏好利用硝酸鈉,另外3株則偏好利用硝酸鉀;YNML52在硝酸鉀上生長(zhǎng)最慢,但在硝酸銨上則最快;YNJH17109在硝酸鈉上生長(zhǎng)最慢。銨態(tài)氮中,YNML52在磷酸二氫銨上具有最大菌落直徑;YNJH17109對(duì)磷酸二氫銨利用率最差;YNJP162在硫酸銨和氯化銨上生長(zhǎng)最慢。蛋白胨是4株炭疽在包括有機(jī)氮在內(nèi)的所有氮源中利用率最高的,且相互間生長(zhǎng)量無(wú)明顯差異;YNML52對(duì)天冬酰胺的利用率最強(qiáng),YNJH17109和GX1655最弱。
2.3 ?不同溫度對(duì)供試菌株菌絲生長(zhǎng)的影響
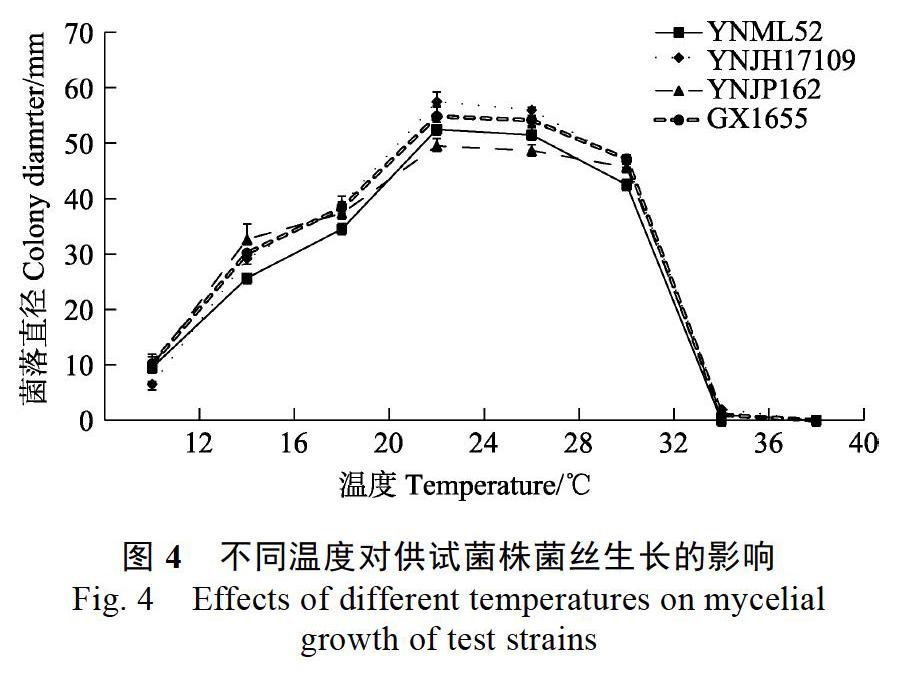
4株尖孢炭疽在供試的不同溫度環(huán)境下生長(zhǎng)情況不同(圖4)。低溫環(huán)境下(12~20 ℃),隨著溫度升高,4株炭疽的菌落直徑均呈不同幅度的增大,其中YNJH17109的菌落直徑增幅最大。適溫環(huán)境內(nèi)(24~28 ℃),溫度升高各菌株間的菌落直徑無(wú)明顯差異,其中28 ℃下菌落直徑最大的是YNJH17109,其后依次是GX1655、YNML52和YNJP162。高溫環(huán)境中(32~40 ℃),32 ℃下4株尖孢炭疽的菌落直徑均顯著減小,YNML52的菌落直徑明顯小于另外3株,其中YNJP162的菌落直徑減幅最小;升高溫度至36 ℃,YNJH17109、YNJP162和GX1655僅呈現(xiàn)極其微弱的生長(zhǎng),而YNML52不能生長(zhǎng);40 ℃下,所有菌株均不能生長(zhǎng)。總體而言,4株尖孢炭疽的最適生長(zhǎng)溫度均為24~28 ℃,YNML52的致死溫度為36 ℃,其余3株的致死溫度為40 ℃。
2.4 ?不同pH對(duì)供試菌株菌絲生長(zhǎng)的影響
4株尖孢炭疽在供試的6個(gè)pH條件下均可生長(zhǎng),梯度間菌落變幅較小(圖5)。YNML52在pH=7時(shí)菌落直徑最大,pH<7時(shí)菌落直徑隨pH上升而增大,pH>7時(shí)則逐漸減小,最適pH為7;YNJH17109在pH為6~8時(shí)菌落直徑均較大,pH為4~6時(shí)菌落直徑逐漸增大,pH>8時(shí)則逐漸減小,最適pH為6~8;YNJP162在pH為7~8時(shí)菌落直徑均較大,pH<7時(shí)菌落直徑隨pH上升而增大,pH>8時(shí)則逐漸減小,最適pH為7~8;GX1655則在pH為5~9時(shí)菌落直徑均較大,pH為4時(shí)菌落直徑最小,最適pH為5~9。
2.5 ?不同水活度對(duì)供試菌株菌絲生長(zhǎng)的影響
不同水活度對(duì)4株尖孢炭疽生長(zhǎng)的影響存在一定差異(圖6)。對(duì)照生長(zhǎng)8 d后,YNJP17109和GX1655的生長(zhǎng)量明顯大于YNML52和YNJP162。添加甘油替代一部分水后,各菌株生長(zhǎng)速率均呈顯著下降趨勢(shì)。水活度為0.99時(shí),YNJP162菌落直徑最大,與對(duì)照相比菌落直徑減幅最大的是YNJH17109和GX1655,減幅分別達(dá)到35.00、48.80 mm。水活度降至0.95時(shí),YNJH17109和GX1655的生長(zhǎng)量極顯著小于YNJP162和YNML52,與對(duì)照相比菌落直徑減幅最大的仍是YNJH17109和GX1655,分別減小了72.50、74.47 mm。當(dāng)水活度降至0.93及以下時(shí),所有菌株均不能生長(zhǎng)。
2.6 ?供試菌株對(duì)不同橡膠品種的致病性評(píng)價(jià)
接種孢子懸浮液4 d后,4株炭疽對(duì)絕大部分橡膠品種均表現(xiàn)致病性,但致病程度存在差異(圖7)。YNML52在PB235、PR107和熱研7-33-97上未表現(xiàn)任何癥狀,在其余品種上潛伏期為2~ 3 d,發(fā)病后在IAN873和RRIC600上病斑擴(kuò)展較快,分別在PB86、IAN873、GT1、文昌11和云研77-4上病斑直徑最小。YNJH17109對(duì)所有供試品種均致病,潛伏期為1.5~2 d,熱研7-33-97發(fā)病后病斑擴(kuò)展較快,在RRIC100上的病斑直徑最大。YNJP162不能侵染PR107,潛伏期為1~2.5 d,RRIC600、熱研7-33-97接種1 d后即發(fā)病,侵染PB86后病斑擴(kuò)展較快,在熱研7-33-97上的病斑直徑最大。GX1655侵染所有供試品種,潛伏期為1.5~2 d,RRIC600發(fā)病后病斑擴(kuò)展較慢,在IAN873上的病斑直徑最大。
2.7 ?供試菌株對(duì)4種作物的致病性評(píng)價(jià)
番木瓜葉片接種4 d后,健康葉片中YNJH-17109和GX1655可使其發(fā)病,形成褐色、萎蔫狀病斑,前者病斑較大;刺傷葉片上除YNML52外均致病,YNJH17109病斑最大,GX1655最小。芒果葉片接種4 d后,健康葉片中僅YNML52不致病,其余均形成黑色、腐爛狀病斑;刺傷葉片上均致病,YNJH17109和YNJP162病斑最大。番石榴葉片接種6 d后,健康葉片中僅YNJP162具有致病力,形成極小黑色點(diǎn)狀病斑;刺傷葉片上除YNML52外均致病,YNJH17109病斑最大,GX1655最小。辣椒葉片接種6 d后,所有處理中僅YNJP162在健康葉片上形成直徑2 mm的極小黑色點(diǎn)狀斑;辣椒果實(shí)接種6 d后,僅見(jiàn)GX1655在刺傷條件下形成褐色、圓形、壞死狀病斑,病斑達(dá)19.50 mm(圖8)。
3 ?討論
增加基因數(shù)目以提高構(gòu)建復(fù)雜程度和準(zhǔn)確性的多位點(diǎn)系統(tǒng)發(fā)育分析方法已成為當(dāng)前刺盤孢屬分類的一大趨勢(shì)。本研究采用的5基因聚類分析結(jié)果支持將編號(hào)YNJH17109的菌株歸入尖孢炭疽復(fù)合種中的C. laticiphilum,2012年[16]報(bào)道該種主要分布于哥倫比亞、斯里蘭卡和印度。國(guó)內(nèi)于2019年在西雙版納植膠區(qū)首次發(fā)現(xiàn)該種[10],與版納炭疽同為省內(nèi)特有種,兩者相對(duì)于華南炭疽分布較為局限。就目前而言,云南是國(guó)內(nèi)最早發(fā)現(xiàn)尖孢炭疽為害的區(qū)域,且物種多樣性較高,一定程度上可能與其靠近東南亞、南亞等尖孢炭疽為害嚴(yán)重的植膠國(guó)有關(guān)。
病原真菌生物學(xué)特性的研究是病害防治中重要的理論來(lái)源,有些研究也將其作為物種劃分的參考依據(jù)。已有文獻(xiàn)[17-20]報(bào)道尖孢炭疽對(duì)果膠和蛋白胨的利用率最高,與本研究中的4株橡膠尖孢炭疽表現(xiàn)一致;張春霞等[21]發(fā)現(xiàn)尖孢炭疽更偏好利用乳糖,這與本研究結(jié)果明顯相反;李菲菲[22]提出銨態(tài)氮會(huì)抑制尖孢炭疽生長(zhǎng),本研究中也觀察到類似現(xiàn)象。真菌菌落生長(zhǎng)速率是鑒別炭疽種群的重要特征[7]。YNML52分別在葡萄糖、蔗糖為碳源和硝酸鈉、硝酸銨、磷酸二氫銨為氮源的基質(zhì)上菌落直徑明顯最大;YNJH17109在半乳糖上最大,分別在磷酸二氫銨、天冬酰胺上最小;YNJP162分別在乳糖、果膠、可溶性淀粉、多聚半乳糖醛酸、堿性木素、檸檬酸鉀和酒石酸鈉上最大,分別在硫酸銨、氯化銨上最小;GX1655則分別在木糖和阿拉伯糖上最大。這可能成為橡膠樹尖孢炭疽復(fù)合種內(nèi)劃分的一種有效方法。溫度、pH和水活度是影響尖孢炭疽生長(zhǎng)的重要因子。已有文獻(xiàn)[18, 22]報(bào)道的尖孢炭疽適生溫度與本研究總體一致,而YNJP162的適生溫度相對(duì)較寬,YNML52對(duì)高溫的耐受性最弱。胡美嬌等[18]認(rèn)為尖孢炭疽在弱酸至弱堿條件下均能生長(zhǎng),但最適pH范圍差異較大,本研究中供試菌株對(duì)環(huán)境酸堿度的要求也不一致。Es-Soufi等[15]證實(shí)尖孢炭疽對(duì)環(huán)境中水含量變化響應(yīng)強(qiáng)烈,本研究與其類似,正常水活度下生長(zhǎng)較緩慢的菌株在少水條件下的生長(zhǎng)量高于正常條件下生長(zhǎng)較快的菌株,推測(cè)較高生長(zhǎng)速率的菌株對(duì)自由水的需求更大。當(dāng)前,隨著復(fù)合種內(nèi)分類地位逐步清晰,準(zhǔn)確的種間差異情況將為開展病原識(shí)別、病害防控等研究提供必要指導(dǎo)。
明確橡膠樹尖孢炭疽種間致病性和寄主范圍的差異情況,結(jié)合各區(qū)域栽培品種和病原種群的分布信息,對(duì)炭疽病害的流行、監(jiān)測(cè)和防治具有一定的參考價(jià)值。總體而言,4株尖孢炭疽對(duì)當(dāng)前多數(shù)主栽品種均具有致病性。但YNML52對(duì)供試中的3個(gè)品種無(wú)致病性,在致病品種中發(fā)病病斑最小且潛伏期最長(zhǎng),因而版納炭疽危害程度相對(duì)較小。已有研究認(rèn)為,離體條件下的菌株致病程度受到孢子懸浮液濃度、潛伏期、菌斑生長(zhǎng)速率、葉片面積大小等因素的共同影響[23]。李文楊等[24]報(bào)道橡膠品種PB260、IAN873、GTI、RRIC600、云研77-4對(duì)橡膠尖孢炭疽的抗性水平較低,可能會(huì)造成較為嚴(yán)重的病害流行,本研究與其相似;但PR107未被YNML52和YNJP162侵染,且另外2個(gè)菌株侵染后病斑擴(kuò)展速率和平均直徑也最小,推測(cè)其對(duì)尖孢炭疽具有一定程度的抗性。尖孢炭疽復(fù)合種的為害多見(jiàn)于熱帶果蔬作物[18, 20, 25],部分種寄主多樣、致病譜廣[16-17]。測(cè)試發(fā)現(xiàn)YNML52僅在刺傷的芒果葉片上表現(xiàn)致病,推測(cè)版納炭疽的寄主專化性程度較高。YNJH17109和GX1655侵染健康的番木瓜和芒果葉片,而后者病斑遠(yuǎn)小于前者;但僅GX1655在刺傷條件下?lián)p毀辣椒果實(shí),推測(cè)YNJH17109在果樹類作物上致病力更強(qiáng),GX1655則在果蔬類作物上危害更大。此外,YNJP162在多數(shù)處理下的致病程度均大于GX1655,且可能是番石榴和辣椒葉片的潛在病原;結(jié)合基礎(chǔ)生物學(xué)特性及橡膠品種致病性方面表現(xiàn)的差異,推測(cè)YNJP162與GX1655并非同一物種,或是一新種,相關(guān)結(jié)論還需分子實(shí)驗(yàn)進(jìn)一步驗(yàn)證。
參考文獻(xiàn)
[1] 陳明文. 我國(guó)天然橡膠產(chǎn)業(yè)發(fā)展形勢(shì)與因應(yīng)策略[J]. 農(nóng)業(yè)經(jīng)濟(jì)問(wèn)題, 2016, 37(10): 91-94, 112.
[2] 黃貴修, 許燦光, 李博勛. 中國(guó)天然橡膠病蟲草害識(shí)別與防治[M]. 2版. 北京: 中國(guó)農(nóng)業(yè)科學(xué)技術(shù)出版社, 2018.
[3] 劉秀娟, 楊業(yè)銅, 冷懷瓊. 我國(guó)植膠墾區(qū)橡膠樹炭疽病菌的種型鑒定[J]. 熱帶作物學(xué)報(bào), 1987(8): 93-101.
[4] Hunupolagama D M, Chandrasekharan N V, Wijesundera W S S, et al. Unveiling members of Colletotrichum acutatum species complex causing Colletotrichum leaf disease of Hevea brasiliensis in Sri Lanka[J]. Current Microbiology, 2017, 74(6): 747–756.
[5] 張春霞, 何明霞, 李加智, 等. 云南西雙版納地區(qū)橡膠炭疽病病原鑒定[J]. 植物保護(hù), 2008, 34(1): 103-107.
[6] Marin-Felix Y, Groenewald J Z, Cai L, et al. Genera of phytopathogenic fungi: GOPHY 1[J]. Studies in Mycology, 2017(86): 99-216.
[7] Liu X B, Huang G X, Cai J M, et al. Diversity of Colletotrichum species causing anthracnose of rubber tree in China[J]. Scientific Reports, 2018(8): 10435.
[8] 林春花, 孫董董, 韓 ?丹, 等. 中國(guó)橡膠樹苗圃2種炭疽病菌分子鑒定及分布分析[J]. 熱帶作物學(xué)報(bào), 2014, 35(9):?1802-1808.
[9] 林春花, 楊 ?歡, 趙曉宇, 等. 海南橡膠樹炭疽菌Colletotrichum siamense和C. fructicola的鑒定及系統(tǒng)發(fā)育分析[J]. 熱帶作物學(xué)報(bào), 2018, 39(1): 129-136.
[10] Shi Y P, Liu Y X, Li L L, et al. First report of Colletotrichum laticiphilum causing anthracnose of rubber tree in China[J]. Plant Disease, 2019, 103(3): 579-580.
[11] Cao X R, Xu X M, Che H Y, et al. Distribution and fungicide sensitivity of Colletotrichum species complexes from rubber tree in Hainan, China[J]. Plant Disease, 2017, 101(10): 1774-1780.
[12] 方中達(dá). 植病研究方法[M]. 北京: 中國(guó)農(nóng)業(yè)出版社, 1998.
[13] 孫廣宇, 宗兆鋒. 植物病理學(xué)實(shí)驗(yàn)技術(shù)[M]. 北京: 中國(guó)農(nóng)業(yè)出版社, 2002.
[14] Prihastuti H, Cai L, Chen H, et al. Characterization of Colletotrichum species associated with coffee berries in northern Thailand[J]. Fungal Diversity, 2009(39): 89-109.
[15] Es-Soufi R, Kbiach M L E, Errabii T, et al. Biology and physiology of Colletotrichum acutatum strains causing strawberrys anthracnose[J]. Agricultural Sciences, 2018, 9(8): 974-990.
[16] Damm U, Cannon P F, Woudenberg J H C, et al. The Colletotrichum acutatum species complex[J]. Studies in Mycology, 2012, 73(1): 37-113.
[17] 夏 ?花. 辣椒上一種新病害病原鑒定及尖孢炭疽菌株比較研究[D]. 長(zhǎng)沙: 湖南農(nóng)業(yè)大學(xué), 2012.
[18] 胡美姣, 李 ?敏, 楊鳳珍, 等. 兩種杧果炭疽病菌生物學(xué)特性的比較[J]. 西南農(nóng)業(yè)學(xué)報(bào), 2005, 18(3): 306-310.
[19] 楊佳文, 趙尊練, 張管曲, 等. 陜西線辣椒炭疽病原菌的鑒定及生物學(xué)特性研究[J]. 西北農(nóng)業(yè)學(xué)報(bào), 2017, 26(11): 1695-1705.
[20] 吳 ?松. 涇陽(yáng)線辣椒炭疽病和葉斑病生物學(xué)特性研究及防控藥劑篩選[D]. 楊凌: 西北農(nóng)林科技大學(xué), 2018.
[21] 張春霞, 李加智, 何明霞, 等. 兩種橡膠炭疽病菌生物學(xué)特性的比較[J]. 西南農(nóng)業(yè)學(xué)報(bào), 2008(3): 667-670.
[22] 李菲菲. 柑橘炭疽病的生物學(xué)特性及其拮抗酵母菌的研究[D]. 武漢: 華中農(nóng)業(yè)大學(xué), 2009.
[23] Cabrera L, Rojas P, Rojas S, et al. Most Colletotrichum species associated with tree tomato (Solanum betaceum) and mango (Mangifera indica) crops are not host-specific[J]. Plant Pathology, 2018, 67(5): 1022-1030.
[24] 李文楊, 鄭春耀, 李超萍, 等. 中國(guó)橡膠樹主栽品系和部分種質(zhì)對(duì)尖孢炭疽病的室內(nèi)抗性評(píng)價(jià)[J]. 熱帶農(nóng)業(yè)工程, 2009, 33(5): 35-40.
[25] 羅 ?霓, 何 ?凡, 范鴻雁. 海南省番木瓜主要真菌病害調(diào)查[J]. 中國(guó)熱帶農(nóng)業(yè), 2008(4): 48-50.
