一株產酸性菊粉酶乳酸菌的篩選及其酶學性質研究
2020-10-22 07:04:50黃澄陳睿朱春燕盧良華王成華
食品與發酵工業 2020年19期
黃澄,陳睿,朱春燕,盧良華,王成華
(廣西大學 輕工與食品工程學院,廣西 南寧,530000)
菊粉是由D-呋喃果糖通過β-2,1-糖苷鍵縮合形成的線性鏈狀多糖,且末端以α-1,2-糖苷鍵連接一個葡萄糖殘基,通常包含2~60個果糖單元[1-2]。作為一種常見的植物貯藏性多糖,菊粉存在于全世界超過36 000種植物組織中[3]。其中,菊芋塊莖含有大量的菊粉,約占其塊莖干重的70%[4],其種植簡單、適應性強[5],成為目前生產菊粉的最常用植物。菊粉作為可再生非糧生物質資源,在生產高果糖漿、生物乙醇、低聚果糖等食品、生物化工領域具有廣闊的市場[6]。
菊粉酶,學名β-2,1-D-果聚糖酶,一種能夠水解β-2,1-D-果聚糖糖苷鍵的水解酶,是開發利用菊粉等果糖基可再生資源的關鍵酶類。根據菊粉酶作用方式的不同,可將菊粉酶分為2類,外切型菊粉酶和內切型菊粉酶。內切菊粉酶可隨機水解菊粉鏈內部的糖苷鍵,生成物主要為低聚果糖;外切菊粉酶則逐一水解菊粉鏈的非還原性末端的糖苷鍵,主要產物為果糖和少量葡萄糖,可被用于菊粉高果糖漿的生產等[7-8]。目前報道的大多數菊粉酶最適pH范圍在4.5~6.5[9-10],而酸性條件(pH<4.5)更利于維持果糖的穩定性、抑制微生物污染以及有效抑制副產物和色素的產生。因此,大力開發高效酸性菊粉酶,對于改進已有酶法工藝、提高生產效率等具有重要的應用前景和商業潛力。國內外已開展了馬克斯克魯維酵母等來源菌株菊粉酶的篩選,同時開展了畢赤酵母等基因工程菌生產重組菊粉酶的研究,但鮮見報道來源于食品級微生物的酸性菊粉酶。PALUDAN-MüLLER等[11]報道了1株產弱酸性菊粉酶的Lactobacilluspentosus,這也是目前報道的唯一一個乳酸菌來源的食源酸性菊粉酶,這為篩選新型酸性乳酸菌菊粉酶提供了思路。本研究從課題組前期構建的來源于廣西特色生榨米粉的乳酸菌庫中篩選出產酸性菊粉酶的乳酸菌,對其分類鑒定,并進行及酶學性質的研究和水解產物的檢測,為酸性菊粉酶的開發應用提供了新的選擇。
1 材料與方法
1.1 菌株與試劑
廣西特色生榨米粉來源乳酸菌,本實驗室保藏;菊粉(90%),上海源葉生物公司;酵母提取物,OXOID 公司;蛋白胨、牛肉膏,Solarbio公司;K2HPO4、MgSO4、MnSO4、醋酸鈉、檸檬酸二銨、吐溫-80、3,5-二硝基水楊酸,成都金山化學試劑有限公司;(NH4)2SO4、NaOH、醋酸、正丁醇、磷酸、異丙醇、二苯胺、丙酮、苯胺、結晶酚,成都市科龍化工試劑廠;微量點樣毛細管、硅膠層析板(20 cm×20 cm),青島海洋化工廠;菊糖、果糖、蔗糖、葡萄糖標準品,Sigma公司;瓊脂糖,OMEGA 公司;細菌基因組DNA試劑盒、DNA純化試劑盒,北京康為世紀生物科技有限公司。
DNS試劑:稱取5.3 g 3,5-二硝基水楊酸、9.9 g NaOH于708 mL水中,充分溶解后加入153 g酒石酸鉀鈉,再加入4.15 g Na2SO3和 3.8 mL 45 ℃水浴溶解的結晶酚,不斷攪拌直至完全溶解后,定容至1 000 mL,貯存于棕色瓶中,于室溫避光放置7 d。
培養基:20 g菊粉、10 g牛肉粉、5 g酵母提取物、10 g蛋白胨、2.0 g檸檬酸二銨、5.0 g乙酸鈉、2 g K2HPO4、0.58 g MgSO4、0.25 g MnSO4、1 mL吐溫-80,1 000 mL蒸餾水,pH 6.6~6.8,配制固體培養基時,加入瓊脂20 g,121 ℃高壓滅菌 15 min。
薄層層析(thin-layer chromatography,TLC)試劑:展開劑為正丁醇、異丙醇、醋酸、水按7∶5∶1∶2(體積比)混合,顯色劑為體積分數2%二苯胺-丙酮溶液、體積分數2%苯胺-丙酮溶液、體積分數85%磷酸按5∶5∶1(體積比)混合。
1.2 儀器設備
SW-SJ-2F超凈工作臺,蘇凈安泰公司;5401D006843 低溫高速離心機,Eppendorf公司;TOMY Autoclave 高壓滅菌鍋,TOMY公司;DYY-8C 電泳儀,北京六一生物科技有限公司;MJ-Ⅱ 電熱恒溫培養箱,上海一恒科技有限公司;Infinite M200 PRO 酶標儀,TECAN公司;ChemiDoc MP 全能型凝膠成像分析儀、T100 梯度PCR儀,Bio-Rad公司。
1.3 實驗方法
1.3.1 產菊粉酶菌株篩選
初篩:將本課題組前期從廣西壯族傳統生榨米粉中篩選出的乳酸菌劃線至固體培養基中(菊粉為唯一碳源),于30 ℃靜置培養直至出現單菌落,將正常生長的單菌落接種至液體發酵培養基中,于30 ℃靜置發酵1~2 d。
復篩:取出15 mL初篩所制備的發酵液在4 ℃條件下,10 000 r/min離心2 min,棄上清液,用600 μL醋酸緩沖溶液(0.2 mol/L、pH 5.0)重懸沉淀,后加入溶菌酶進行破胞,破胞結束后,按上述相同條件離心40 min,檢測上清液中的菊粉酶酶活力,選出其中酶活力最高的菌株 1-1進行后續試驗。
1.3.2 酶活力測定
酶活力測定中所用底物質量濃度均為5 g/L,菊粉溶液和蔗糖溶液均用醋酸緩沖液(0.2 mol/L、pH 5.0)配制而成。
參照PESSONI等[12]的方法對菊粉酶進行酶活力測定。取100 μL酸性菊粉酶分別加入200 μL 5 g/L菊粉溶液和200 μL 5 g/L蔗糖溶液,在30 ℃水浴中放置30 min后立即100 ℃滅活5 min,冷卻后加入300 μL DNS,置于100 ℃水浴中7 min進行顯色,迅速冷卻后在540 nm波長處測定吸光值,參照葡萄糖標準曲線來計算還原糖含量,以滅活的酶液作為空白對照。設3次重復試驗,取平均值。
酶活力定義[13]:在30 ℃、pH 5.0的條件下,每1 min產生1 μmol還原糖所需的酶量,即為1個酶活力單位(U)。其中1個菊粉酶活力(I)單位定義為每1 min催化菊粉轉化為1 μmol果糖所需的酶量,1個轉化酶活力(S)單位定義為每1 min催化蔗糖轉化為1 μmol果糖所需的酶量。
(1)
式中:E,樣品酶活力,U/mL;c,標準曲線上對應的葡萄糖濃度,μmol/mL;V1,反應液總體積,mL;n,粗酶液稀釋倍數;T,反應時間,min;V2,反應液中粗酶體積,mL。
1.3.3 菌株鑒定
通過菌落觀察和基因序列分析對菌株進行鑒定。使用細菌基因組DNA試劑盒,按照說明書提取菌株基因組DNA,使用超微量全波長分光光度計對提取的基因DNA樣液進行定量,將其稀釋至1 ng/μL,并以此作為PCR擴增的模板。使用細菌通用引物27F:5′-AGAGTTTGATCCTGGCTCAG-3′和1429R:5′-GGTTACCTTGTTACGACTT-3′進行PCR擴增,擴增條件為94 ℃預變性5 min,94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1 min,進行30次循環,72 ℃終延伸2 min。取5 μL PCR產物進行0.8%瓊脂糖凝膠電泳驗證,將驗證后的PCR產物通過DNA純化試劑盒進行純化,再由六合華大基因科技有限公司進行測序,測序結果輸入基因數據庫(GenBank)進行同源性分析。
1.3.4 酶學性質研究
1.3.4.1 溫度對粗酶液酶活力和穩定性的影響
參照1.3.2的酶活力測定,將菊粉酶分別在20~80 ℃進行反應,每5 ℃為1個溫度梯度,測定酶活力,以測定結果最高酶活力為100%,計算各測定組的相對酶活力;將菊粉酶分別在20~80 ℃保溫30 min,冷卻后測定酶活力,以未經處理酶的酶活力為100%,計算各組的相對酶活力。
1.3.4.2 pH對粗酶液活力和穩定性的影響
參照1.3.2的酶活力測定,配制不同pH的緩沖液,pH 3.5~5.0為醋酸緩沖液,pH 5.5~7.5為磷酸緩沖液,pH 8.0~9為Tris-HCl緩沖液,pH 9.5~11.0為碳酸緩沖液。將菊粉酶與上述不同pH緩沖液配制的菊粉溶液進行反應,測定酶活力,以最高結果為100%,計算各測定組的相對酶活力;用不同pH的緩沖液稀釋菊粉酶液,于4 ℃放置1 h,測定酶活力,以未經處理酶的酶活力為100%,計算各組的相對酶活力。
1.3.5 TLC分析水解產物
以菊粉為底物,將200 μL菊粉酶與100 μL底物在最適溫度、最適pH條件下水解10 h,得到水解產物。TLC方法參照文獻報道[14]。用去離子水將各標準品配制成質量濃度為2.0 mg/mL的標準品試劑,用微量點樣毛細管依次對標準品試劑和水解產物進行點樣,點樣量5 μL。展開完成后噴霧顯色劑,待自然風干后置于105 ℃加熱5 min,即可進行觀察。
2 結果與分析
2.1 產菊粉酶菌株篩選
在本課題組的廣西特色生榨米粉來源乳酸菌庫中,篩選出1株可以正常生長在以菊粉為唯一碳源的培養基上,且具有相對較高酶活力的菌株1-1,后續試驗均選用此菌株。以菊粉為底物,在30 ℃,pH 5.0條件下進行3組平行試驗,取平均值,測得菌株1-1的菊粉酶活力I=(0.23±0.03) U/mL;以蔗糖為底物,在30 ℃,pH 5.0條件下進行3組平行試驗,取平均值,測得菌株1-1的轉化酶活力S=(0.92±0.06) U/mL,兩者的比值(I/S)為0.25,I/S<1表明該酶為外切菊粉酶[15],其菊粉酶活力略低于王翠[7]報道放線菌F01的菊粉酶活力(0.37 U/mL)。
2.2 菌株鑒定
菌株1-1菌落形態如圖1所示,圓形隆起,表面光滑或稍粗糙,呈乳白色或暗黃色。以基因組DNA為模板,PCR擴增16S rDNA序列。PCR產物凝膠電泳結果如圖2所示,擴增條帶在1 500 bp左右,與預期相符。
M-marker; 1-菌株圖1 菌株1-1平板培養菌落形態Fig.1 Colony morphology of 1-1
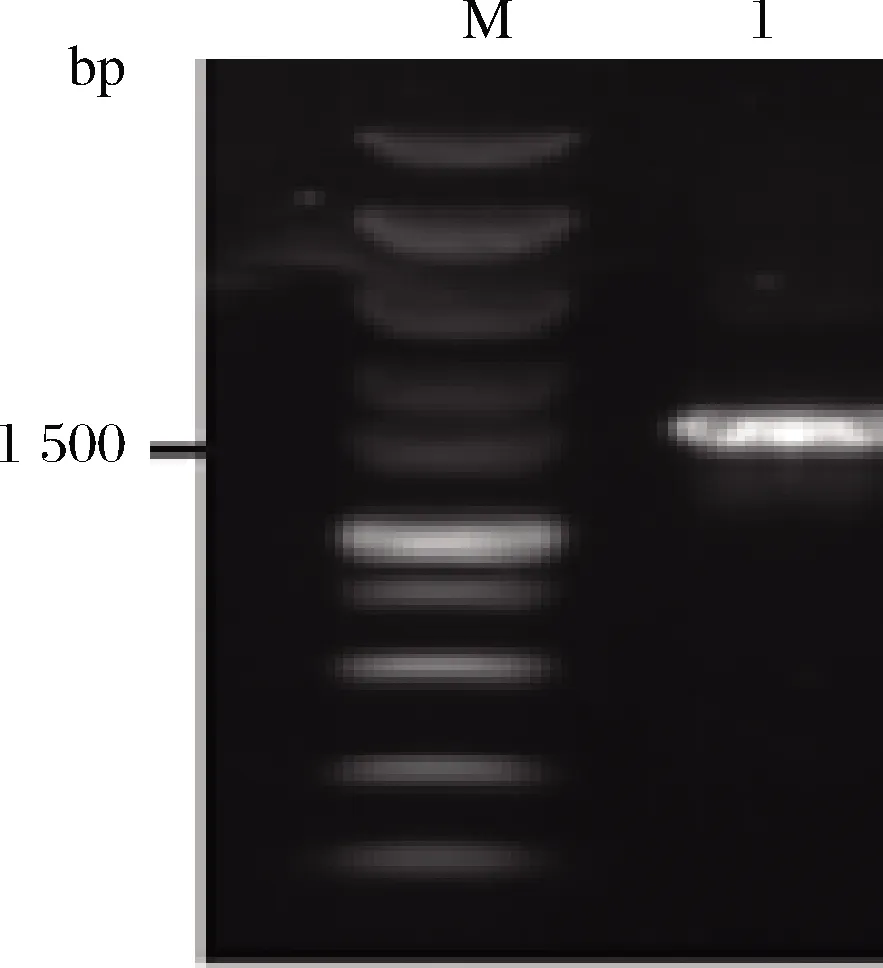
M-marker;1-菌株圖2 PCR產物瓊脂糖凝膠電泳圖Fig.2 Agarose gel electrophoresis of PCR
16S rDNA基因序列與GenBank中進行序列比對,發現其與檸檬明串珠菌Leuconostoccitreumstrain CBA3623相似率達99.59%,可初步鑒定該菌株為檸檬明串珠菌Leuconostoccitreum,將其命名為Leuconostoccitreum1-1,并將該菌株保藏于中國典型培養物保藏中心,保藏號為CCTCC NO:M 2019971。
2.3 酶學性質研究
2.3.1 溫度對菊粉酶酶活力和穩定性的影響
以菊粉為底物,對菌株1-1所產菊粉酶的最適反應溫度和溫度穩定性進行測定,結果如圖3所示。
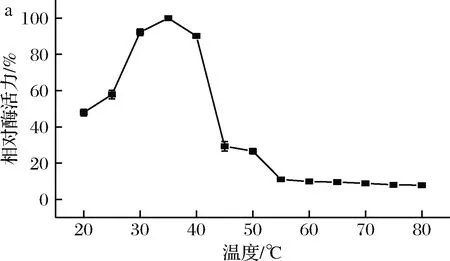
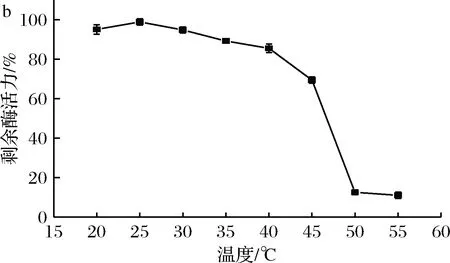
a-溫度對酶活力的影響; b-溫度對穩定性的影響圖3 溫度對酶活力和穩定性的影響Fig.3 Effect of temperature on activity and stability of the inulinase
由圖3-a可知,該酶的最適反應溫度為35 ℃,比PALUDAN-MULLER人[11]報道的Lactobacilluspentosus所產菊粉酶最適溫度高10 ℃。由圖3-b可知,該酶在熱處理30 min條件下,20~45 ℃保持60%以上的最大酶活力。以上結果表明篩選的菊粉酶具有相對較高的最適反應溫度和較寬的溫度耐受范圍,在工業生產中具有一定的開發潛力。
2.3.2 pH對菊粉酶活力和穩定性的影響
菌株1-1菊粉酶的最適反應pH和pH穩定性測定結果如圖4所示。由圖4-a可知,該酶的最適反應pH為4.0,低于目前已報道的菊粉酶,如于春等[16]報道的灰平鏈霉菌StreptomycesgriseoplanusS501的最適pH(5.0)和高威[17]報道的BacillussmithiiT4的最適pH(4.5)。如圖4-b所示,該酶在pH 3.5~10.0較寬泛范圍內,保持65%以上的酶活力。該酸性菊粉酶有著較強的耐酸特性和較寬的pH耐受范圍,有利于菊粉的水解以及抑制副產物的產生。
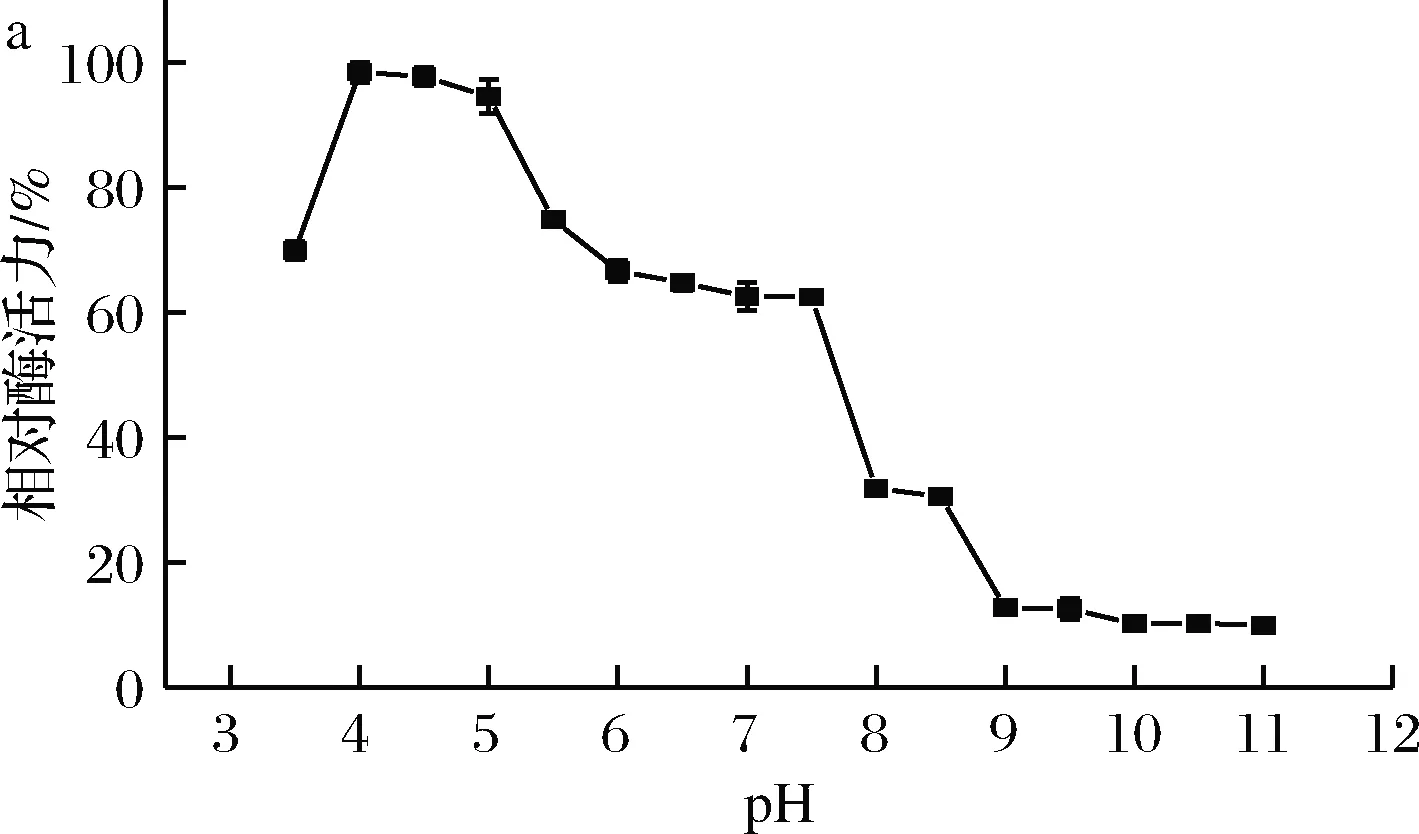
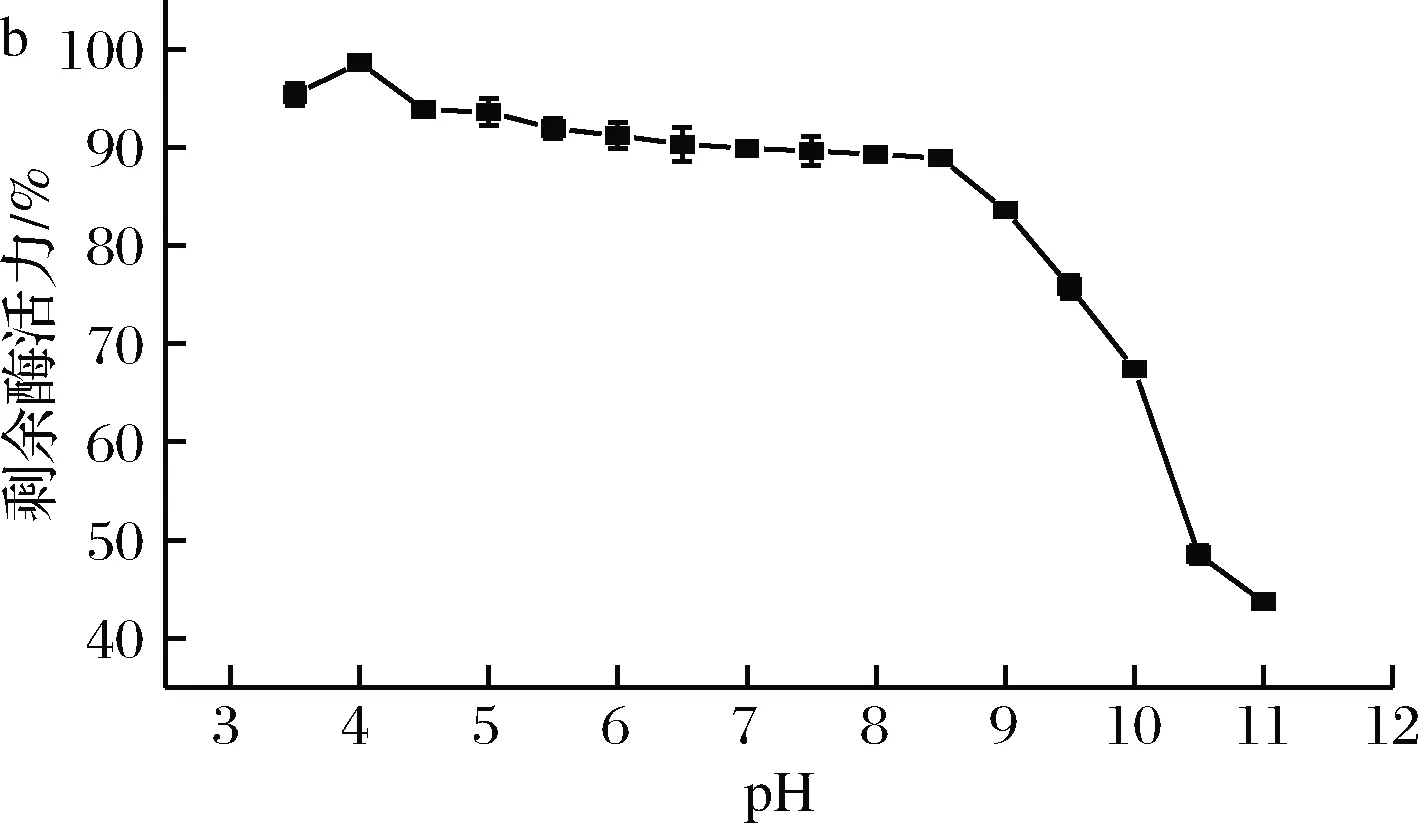
a-pH對酶活力的影響; b-pH對穩定性的影響圖4 pH對酶活力和穩定性的影響Fig.4 Effect of pH on activity and stability of the inulinase
2.3.3 TLC分析水解產物
采用TLC對L.citreum1-1菊粉酶水解菊粉的產物進行分析,結果如圖5所示。
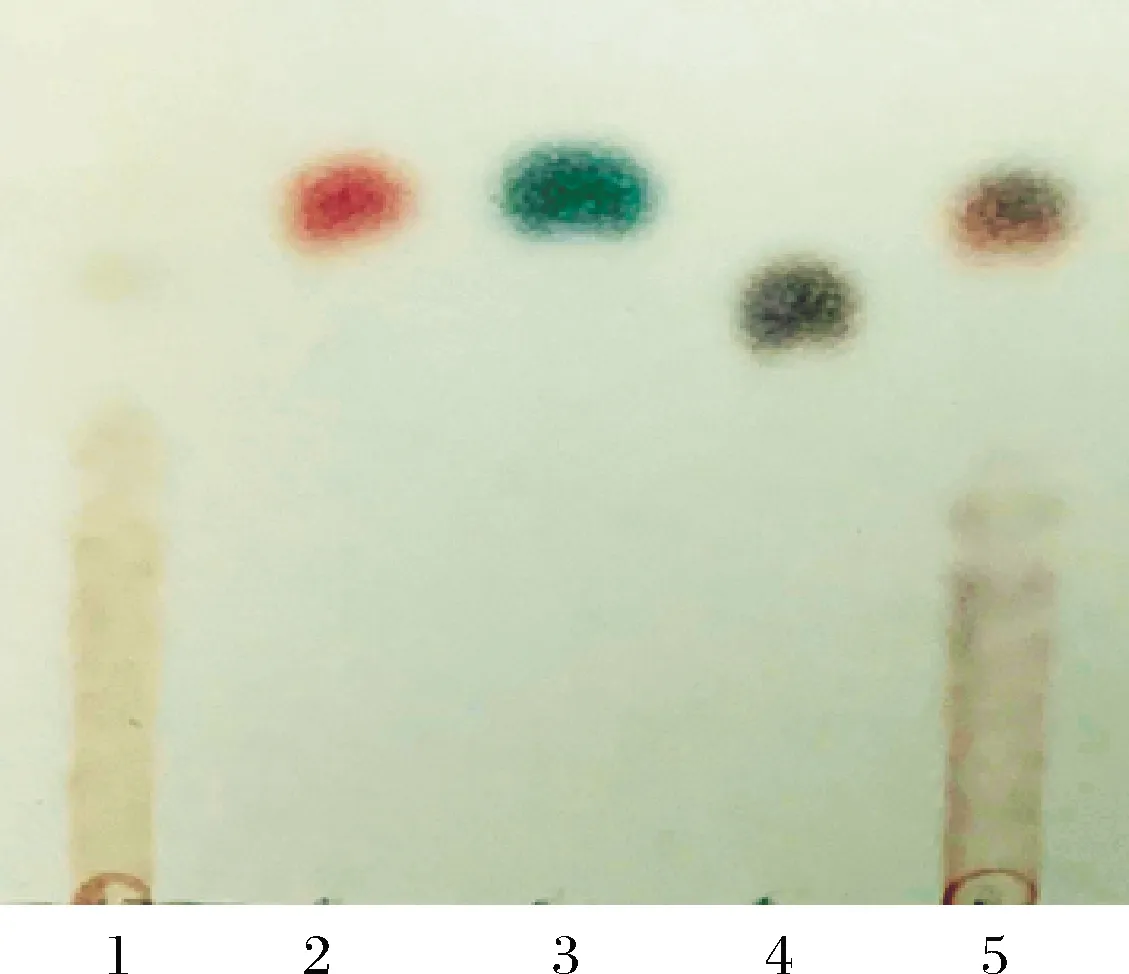
1-菊糖;2-果糖;3-葡萄糖;4-蔗糖;5-水解產物圖5 菊粉酶水解產物TLC分析Fig.5 TLC for characterization of inulin hydrolysis products by inulinase
從圖5可知,在菊粉酶與菊粉反應后,菊粉得到了明顯的降解,降解產物中幾乎100%為單糖,產生大量果糖以及少量葡萄糖,無蔗糖成分,這與L.citreum1-1菊粉酶為外切菊粉酶從非還原末端逐一降解糖苷鍵生成果糖和少量葡萄糖的作用方式相一致,同時與根據I/S值的鑒定結果一致。因此,L.citreum1-1菊粉酶具有應用于高果糖漿生產的潛力。
3 結論
研究首次從廣西壯族傳統生榨米粉中篩選出1株產酸性外切菊粉酶的檸檬明串珠菌L.citreum1-1。該菌株所產菊粉酶的酶活力為0.23 U/mL,轉化酶活為0.92 U/mL,最適反應溫度為35 ℃,最適反應pH為4.0,在20~45 ℃和pH 3.5~10.0的較寬范圍內保持穩定,水解產物為大量果糖和少量葡萄糖。
研究目前對L.citreum1-1只進行了初步研究,未對發酵條件進行優化,可能是其酶活力較低的原因,但是該酶具有更低的最適反應pH和較強的耐酸性,更有利于大規模生產,且該酶來源于傳統食品來源的檸檬明串珠菌,作為一種食源性乳酸菌酸性菊粉酶,其安全性在食品行業具有廣闊的發展前景和商業價值。今后的研究將致力于菌株產酶發酵條件的優化,以及基因重組的手段實現菊粉酶的高效表達和酶學性質表征,為工業發酵生產菊粉酶制劑以及高果糖漿的開發利用打下基礎。
