干酪乳桿菌細(xì)菌素的分離純化及抑菌特性分析
2020-10-10 07:22:00高兆建曹珊珊宋玉林丁飛鴻趙宜峰焦魏
食品與發(fā)酵工業(yè) 2020年18期
高兆建,曹珊珊,宋玉林,丁飛鴻,趙宜峰,焦魏
1(徐州工程學(xué)院,食品(生物)工程學(xué)院,江蘇 徐州,221018)2(長(zhǎng)江桂柳食品睢寧有限公司,江蘇 徐州,221000)3(邳州市金大地肥料有限公司,江蘇 徐州,221300)
PurificationofbacteriocinfromLactobacilluscaseiandanalysisofantibacterialcharacteristics
GAO Zhaojian1*,CAO Shanshan1,SONG Yulin1,DING Feihong1,ZHAO Yifeng2,JIAO Wei3
1(College of Food (Biological) Engineering, Xuzhou Institute of Technology, Xuzhou 221018, China)2(Yangtze River Guiliu Food Suining Co., Ltd., Xuzhou 221000, China)3(Pizhou Golden Earth Fertilizer Co., Ltd., Xuzhou 221300, China)
ABSTRACTThe aim of this work was to purify and characterize the bacteriocin (named as BaC21)produced byLactobacilluscaseiXbC-36 and laid a theoretical foundation for the further development of BaC21 as a new food preservative. BaC21 was purified by ammonium sulphate precipitation, Sephadex G-15 column chromatography and reverse-phase high-performance liquid chromatography. Molecular weight of BaC21 was determined by tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis (Tricine-SDS-PAGE). Production of bacteriocin BaC21 in MRS medium reached the maximum yield of 1 695 AU/mL at the beginning of the stationary phase (18 h). The molecular weight of BaC21 was 4.4 kDa. Because its molecular mass was not similar to other known bacteriocins, BaC21 could be a novel bacteriocin. Then, the antimicrobial activity of BaC21 was nearly not affected by heating at 37-100 ℃ for up to 30 min and decreased but not fully suppressed when exposed to a temperature of 120 ℃ for 15 min, indicating high thermal stability of BaC21. BaC21 remained stable at pH values ranging from 2 to 8. Antimicrobial activity was also not affected by organic solvents and surfactants. And, SDS improved its antibacterial activity. BaC21 was of proteinaceous nature, in which complete inactivation of its antimicrobial activity was observed after being treated with proteases, while lipase and α-amylase exhibited no effect. In addition, BaC21 had broad antimicrobial spectra and especially for Gram-negative food-borne pathogens. It revealed a bacteriostatic mode of action againstStaphylococcusaureusandEscherichiacoliin culture media. In conclusion, bacteriocin BaC21 shows great potential as a food bio-preservative.
KeywordsLactobacilluscasei; bacteriocin; separation and purification; antibacterial characteristics
近年來(lái),全球消費(fèi)者對(duì)安全天然食品重視程度日益增長(zhǎng),人們?cè)絹?lái)越追求健康的生活方式,對(duì)食品加工有了更高的要求,如食品加工過程簡(jiǎn)單、維生素含量豐富、對(duì)健康有益、方便食用、感官特性保持完好、貨架期保存優(yōu)良等。滿足這些期望是當(dāng)前食品工業(yè)面臨的重大挑戰(zhàn)。伴隨著消費(fèi)者對(duì)食品的生物保藏需求不斷增長(zhǎng),各種化學(xué)防腐劑、添加劑、抗生素等安全性問題得到普遍關(guān)注[1]。生物防腐劑除了減少因食物變質(zhì)而造成的經(jīng)濟(jì)損失外,還有助于保存食物的天然味道。因此,研究和開發(fā)安全高效的生物抑菌劑至關(guān)重要。乳酸菌是一群能利用碳水化合物生成乳酸的微生物,在自然界分布廣泛,是公認(rèn)的食品級(jí)安全微生物,并且在醫(yī)藥及食品等領(lǐng)域具有極高的應(yīng)用價(jià)值。
乳酸菌在糖發(fā)酵過程中產(chǎn)生各種各樣的天然代謝物,如乳酸、雙乙酰、過氧化氫和細(xì)菌素,同時(shí)賦予發(fā)酵食品獨(dú)特的風(fēng)味、質(zhì)地和香氣。細(xì)菌素是由某些細(xì)菌在代謝過程中通過核糖體合成的一類具有抗菌生物活性的蛋白質(zhì)或多肽[2],能夠抑制多種病原菌和腐敗菌,而產(chǎn)生菌對(duì)其有自身免疫性[3-4]。細(xì)菌素多數(shù)由革蘭氏陽(yáng)性(Gram-positive,G+)菌產(chǎn)生,并且可以抑制其他親緣關(guān)系較近的菌株,而對(duì)大多數(shù)革蘭氏陰性(Gram-negative,G-)菌、真菌等均沒有抑制作用,如乳酸鏈球菌素。由于細(xì)菌素?zé)o毒、無(wú)殘留、無(wú)抗藥性,可以作為一種生物防腐劑廣泛應(yīng)用于食品工業(yè)[1],近年來(lái)不同來(lái)源的新型細(xì)菌素受到科研工作者的廣泛關(guān)注,眾多新型細(xì)菌素被陸續(xù)報(bào)道,如HU等[5]從發(fā)酵肉中分離的LactobacillusalimentariusFM-MM4細(xì)菌素對(duì)細(xì)菌及酵母菌有廣譜抗菌活性;周佳[6]所研究的腸球菌素SAU-2有良好的耐熱性與酸堿性,并且對(duì)G+菌有明顯抑菌作用,但對(duì)G-菌和真菌基本無(wú)抑菌活性;還有ZACHAROF等[4]對(duì)乳酸菌素的研究,COTTER等[7]對(duì)細(xì)菌素免疫的研究。乳酸鏈球菌素(nisin)是目前已經(jīng)商業(yè)化生產(chǎn)應(yīng)用的細(xì)菌素,對(duì)G+細(xì)菌有良好的抑菌效果,但它對(duì)G-菌、酵母和霉菌無(wú)抑菌效果,在中性pH環(huán)境下穩(wěn)定較低,因此僅限于酸性食品使用[8]。盡管近年來(lái)不同特性的細(xì)菌素有大量報(bào)道,但在抑菌譜、穩(wěn)定性、產(chǎn)量、抑菌活力及適用范圍等方面還存在眾多缺陷,故繼續(xù)開發(fā)新的細(xì)菌素資源,深入研究其特性仍十分必要。
本研究以實(shí)驗(yàn)室分離的干酪乳桿菌XbC-36為菌種,從其發(fā)酵液中分離純化細(xì)菌素,并對(duì)其生長(zhǎng)動(dòng)力學(xué)、抑菌譜、相對(duì)分子質(zhì)量、抑菌活性以及穩(wěn)定性等方面分析,為該細(xì)菌素在天然食品防腐劑開發(fā)方面奠定理論基礎(chǔ)。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株
本研究所用產(chǎn)細(xì)菌素菌株干酪乳桿菌(Lactobacilluscasei)XbC-36分離自山東臨沂農(nóng)村自然發(fā)酵豆制品,并通過菌體菌落形態(tài)、生理生化特征及16s rDNA序列分析鑒定,保藏于徐州工程學(xué)院食品與生物工程中心。
1.1.2 試劑及培養(yǎng)基
試劑:柱層析填充材料Sephadex G-15,美國(guó)Pharmacia公司;胰化蛋白胨、酵母提取物,英國(guó)Oxoid公司;蛋白酶K、胃蛋白酶、胰蛋白酶、α-胰凝乳蛋白酶、脂肪酶、α-淀粉酶,上海源葉生物科技有限公司;其他試劑均為國(guó)產(chǎn)分析純。
培養(yǎng)基:MRS培養(yǎng)基、營(yíng)養(yǎng)瓊脂(nutrient agar,NA)培養(yǎng)基、馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)培養(yǎng)基均由實(shí)驗(yàn)室配制而成,各菌株所用培養(yǎng)基及培養(yǎng)溫度如表1所示。
1.1.3 儀器與設(shè)備
Sigma3k15冷凍離心機(jī),德國(guó)SIGMA公司;UV-2450紫外可見光分光光度計(jì),日本島津公司;DYY-6B凝膠水平電泳儀,北京市六一儀器廠;AKTAprime蛋白純化系統(tǒng),美國(guó)GE Healthcare公司;Agilent 1260半制備高效液相色譜儀、Zorbax SB-C18高效液相色譜半制備柱,美國(guó)安捷倫公司;CascadaTM AN超純水系統(tǒng),美國(guó)PALL公司。
1.2 實(shí)驗(yàn)方法
1.2.1 菌體生長(zhǎng)動(dòng)力學(xué)及細(xì)菌素合成
將活化后的菌株XbC-36按照接種量1%接種到MRS液體培養(yǎng)基(pH 6.5),培養(yǎng)溫度32 ℃,培養(yǎng)時(shí)間48 h,每隔3 h取樣測(cè)定其吸光度及抑菌活性。
通過瓊脂孔擴(kuò)散法測(cè)定抑菌活性,具體測(cè)定參照文獻(xiàn)[9-11]并稍作修改。制備對(duì)應(yīng)不同指示菌的瓊脂培養(yǎng)基平板,指示菌調(diào)整到2.0×106CFU/mL并在平板上涂布150 μL,晾干后打孔。100 μL發(fā)酵液的上清液注入孔中,最適溫度下培養(yǎng)1~5 d,測(cè)定抑菌圈直徑大小。抑菌活力單位的定義為以S.aureus為指示菌,樣品經(jīng)最大稀釋后,打孔檢測(cè)仍產(chǎn)生清晰抑菌圈的最大稀釋倍數(shù)的倒數(shù)作為1個(gè)抑菌活力單位。
1.2.2 細(xì)菌素的純化
BaC21的發(fā)酵制備:MRS瓊脂平板活化2次的L.caseiXbC-36接種于MRS液體培養(yǎng)基,32 ℃培養(yǎng)16 h后,取5 mL 菌液接種于495 mL MRS液體培養(yǎng)基中。在32 ℃下培養(yǎng)18 h,10 000 r/min、4 ℃離心10 min,取上清液并經(jīng)濾膜過濾后于80 ℃加熱15 min,調(diào)節(jié)pH 7.0,得到去細(xì)胞發(fā)酵上清液(cell-free fermentation supernatant,CFS),于4 ℃冰箱短時(shí)保存?zhèn)溆谩?/p>
分子篩凝膠過濾層析:以0.02 mol/L pH 7.0的磷酸緩沖液充分平衡層析柱(80 cm×1.5 cm)。經(jīng)(NH4)2SO4鹽析、透析并超濾濃縮后的樣品上樣,相同緩沖液洗脫,流速0.8 mL/min。收集各洗脫峰,測(cè)抑菌活性,合并有活性收集管,超濾濃縮后備用。
半制備反向高效液相色譜(reverse-phase high-performance liquid chromatography, RP-HPLC)純化BaC21: RP-HPLC線性梯度洗脫,1 mL/min洗脫速度,溶劑A:體積分?jǐn)?shù)5%乙腈中含有體積分?jǐn)?shù)0.1%三氟乙酸(trifluoroacetic acid,TFA),溶劑B:體積分?jǐn)?shù)100%乙腈中含有體積分?jǐn)?shù)0.1% TFA,52 min內(nèi)溶劑A從體積分?jǐn)?shù)85%到體積分?jǐn)?shù)30%,溶劑B從體積分?jǐn)?shù)15%到體積分?jǐn)?shù)70%。手動(dòng)收集洗脫峰,真空濃縮器中徹底去除乙腈,抑菌活性檢測(cè),活性組分合并、凍干并進(jìn)一步分析。
1.2.3 Tricine-SDS-PAGE測(cè)定BaC21分子質(zhì)量
純化的BaC21 Tricine-SDS-PAGE測(cè)定純化程度及分子質(zhì)量。電泳結(jié)束后凝膠的一部分用考馬斯亮藍(lán)染色,另一半無(wú)菌蒸餾水中洗滌3 h,置于培養(yǎng)皿中,LB(luria-bertani)培養(yǎng)基覆蓋,涂布S.aureus并37 ℃培養(yǎng)16~18 h,與染色脫色后的蛋白凝膠比較,觀察抑菌條帶。
1.2.4 酶、有機(jī)溶劑、化學(xué)試劑、pH和溫度對(duì)細(xì)菌素的影響
用胰蛋白酶、α-胰凝乳蛋白酶、胃蛋白酶、蛋白酶K、脂肪酶和α-淀粉酶做酶解實(shí)驗(yàn)。調(diào)節(jié)部分純化的BaC21至酶最適pH,加入終質(zhì)量濃度2 mg/mL的酶,37 ℃水浴2 h,80 ℃加熱15 min滅酶,檢測(cè)抑菌活性。
分別向部分純化的BaC21中添加體積分?jǐn)?shù)1%的乙醇、異丙醇、丙酮、乙腈,1 g/L的Tween-80、Tween-20、Triton X-100和SDS, 37 ℃處理2 h后測(cè)定抑菌活性。
將部分純化的BaC21的pH調(diào)至2~9,37 ℃下處理2 h,測(cè)定其抑菌活性。
將部分純化的BaC21取5 mL分別置于37、60、80、100 ℃保溫30 min,121 ℃處理15 min,待溫度恢復(fù)到室溫測(cè)定抑菌活性。
以上活性檢測(cè)均以S.aureus為指示菌采用瓊脂孔擴(kuò)散法測(cè)定。空白對(duì)照均是以未經(jīng)過任何處理的BaC21為樣品。
1.2.5 細(xì)菌素對(duì)S.aureus和E.coli的抑菌作用
將S.aureus和E.coli在37 ℃下培養(yǎng)至指數(shù)期中期(4 h),按照兩指示菌100 mL培養(yǎng)液加入20 mLL.caseiXbC-36 CFS,并繼續(xù)培養(yǎng),每隔2 h測(cè)定600 nm處的吸光度。對(duì)照組為未加入CFS正常培養(yǎng)的菌體。
1.2.6 數(shù)據(jù)統(tǒng)計(jì)與分析方法
數(shù)據(jù)統(tǒng)計(jì)分析參照陳文瑩等[12]報(bào)道的方法,檢測(cè)實(shí)驗(yàn)均重復(fù)3次,結(jié)果以平均值±標(biāo)準(zhǔn)差(SD)表示。對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行方差分析(ANOVA),采用SNK方法評(píng)價(jià)差異顯著的結(jié)果進(jìn)行兩兩比較,P<0.05認(rèn)為有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果與分析
2.1 干酪乳桿菌XbC-36生長(zhǎng)及細(xì)菌素合成
L.caseiXbC-36生長(zhǎng)曲線及BaC21合成情況如圖1所示,接種6 h后開始合成BaC21(126 AU/mL),在發(fā)酵6~18 h,BaC21的產(chǎn)量與吸光值成正比,抑菌活性顯著增加(P<0.05),在穩(wěn)定期初始階段菌體濃度最大時(shí)(OD600=2.156)合成最大量BaC21(1 695 AU/mL)。而在18 h后可能由于酸性抑制、菌體分泌蛋白酶的降解等因素導(dǎo)致BaC21活性略有下降[13],但在36~48 h,抑菌活性變化不明顯(P>0.05),而菌體吸光值基本保持不變(P>0.05)。不同來(lái)源細(xì)菌素達(dá)到最大產(chǎn)量的時(shí)間差異較大,如YI等[14]研究的Lactobacilluscrustorum細(xì)菌素60 h最大,WORAPRAYOTE等[15]報(bào)道的Weissellahellenica發(fā)酵12 h最大,PERUMAL等[16]報(bào)道的Enterococcusfaecalis細(xì)菌素發(fā)酵20 h產(chǎn)量最高。BaC21在指數(shù)期開始合成,隨著菌體生長(zhǎng)BaC21產(chǎn)量增大并在穩(wěn)定期達(dá)到最高,因此BaC21屬于初級(jí)代謝產(chǎn)物,這與MIRANDA等[17]、MARTINEZL等[18]研究的細(xì)菌素一致,但從細(xì)菌素發(fā)酵產(chǎn)量看L.caseiXbC-36所產(chǎn)細(xì)菌素更占優(yōu)勢(shì)。

圖1 L. casei XbC-36生長(zhǎng)動(dòng)力學(xué)及BaC21合成Fig.1 Growth kinetics and bacteriocin biosynthesis of L. casei XbC-36注:未標(biāo)注小寫字母表示差異顯著(P<0.05),相同小寫字母表示差異不顯著(P>0.05)
2.2 細(xì)菌素的純化
分子篩凝膠柱層析結(jié)果如圖2所示,經(jīng)層析得到5個(gè)明顯分離的洗脫峰。抑菌活性檢測(cè)顯示P3蛋白質(zhì)峰出現(xiàn)抑菌圈,收集P3峰樣品進(jìn)一步純化。
RP-HPLC如圖3所示,0~52 min很多不同種類的物質(zhì)被洗脫出,表明經(jīng)分子篩凝膠柱的樣品含有較多雜質(zhì)且經(jīng)RP-HPLC分離明顯,36 min后無(wú)洗脫峰出現(xiàn),說明已經(jīng)洗脫充分。手動(dòng)收集11個(gè)峰的樣品檢測(cè)抑菌活性,保留時(shí)間19 min的洗脫產(chǎn)物即4號(hào)峰有抑菌活性。RP-HPLC是純化細(xì)菌素的高效方法,據(jù)報(bào)道陳一然[19]、PIARD等[20]、方佳琪[21]也采用該方法純化到了細(xì)菌素。
2.3 細(xì)菌素的分子質(zhì)量及原位活性檢測(cè)
Tricine-SDS-PAGE如圖4所示,RP-HPLC得到的活性樣品1.2~4.6 kDa顯示1條清晰條帶,說明凝膠過濾層析和RP-HPLC可以有效純化BaC21,根據(jù)標(biāo)準(zhǔn)蛋白marker遷移率計(jì)算得到樣品分子質(zhì)量4.4 kDa。凝膠原位抑菌實(shí)驗(yàn)顯示,條帶對(duì)應(yīng)位置產(chǎn)生明顯的抑菌條帶。由此可確定,考馬斯亮藍(lán)染色顯示的電泳條帶為有抑菌活性的目的產(chǎn)物,其分子質(zhì)量在報(bào)道的絕大多數(shù)細(xì)菌素分子質(zhì)量范圍內(nèi)。據(jù)報(bào)道,不同微生物所產(chǎn)細(xì)菌素分子質(zhì)量通常存在差異,BaC21分子質(zhì)量與KAKTCHAM等[22]研究的乳鏈球菌素2MT分子質(zhì)量相近(4.1 kDa),但比來(lái)源于XI等[23]研究的糞腸球菌TG2(6.3 kDa)、AYED等[24]研究的芽孢桿菌(11 kDa)、GOH等[25]研究的腸球菌C1(10 kDa)的細(xì)菌素要小;比趙圣明等[26]報(bào)道的植物乳桿菌細(xì)菌素(0.9 kDa)分子質(zhì)量要大。由此推測(cè),BaC21可能是一種新型細(xì)菌素。

圖2 Sephadex G-15凝膠層析及BaC21抑菌檢測(cè)Fig.2 Sephadex G-15 gel chromatography and bacteriostatic experiments of BaC21
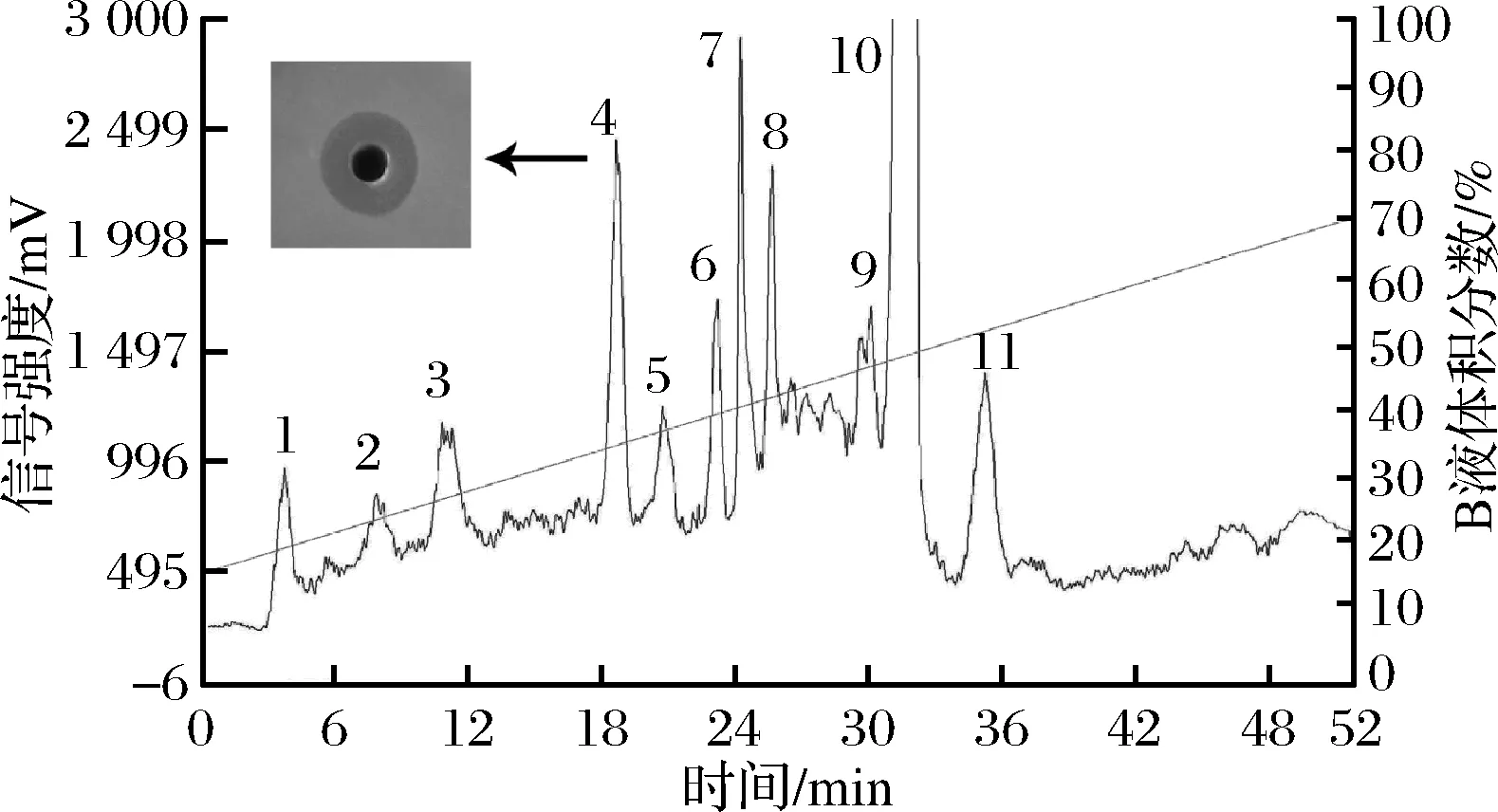
圖3 RP-HPLC純化L. casei XbC-36 BaC21Fig.3 Purification of BaC21 from L. casei XbC-36 by RP-HPLC

M-標(biāo)準(zhǔn)分子質(zhì)量蛋白;1-純化后樣品;2-凝膠原位抑菌檢測(cè)圖4 Tricine-SDS-PAGE分析及抑菌活性檢測(cè)Fig.4 Tricine-SDS-PAGE analysis and bacteriostatic activity detection
2.4 細(xì)菌素的抗菌譜
BaC21抑菌情況如表1所示。BaC21對(duì)G+菌有較弱的抑菌活性,而對(duì)S.aureus抑制作用非常強(qiáng),但對(duì)乳酸乳球菌無(wú)抑制作用。對(duì)大部分G-菌有強(qiáng)的抑制活性,但對(duì)于霍亂弧菌的抑制活性相對(duì)較弱,對(duì)真菌則無(wú)抑菌活性。整體看,BaC21對(duì)G-菌具有很強(qiáng)的抑菌作用,對(duì)G+菌稍弱。通常,因G-菌細(xì)胞壁中缺乏脂壁酸故細(xì)菌素不能抑制G-菌[13],例如已經(jīng)商業(yè)化的食品防腐劑nisin僅對(duì)G+菌有抗菌活性[27],董雨馨等[28]研究的瑞士乳桿菌M14-1所產(chǎn)細(xì)菌素只對(duì)單核細(xì)胞增生李斯特氏菌有良好的抑菌效果,BENBRAEK等[29]研究的乳糖腸球菌4CP3只對(duì)真菌和G+菌有較強(qiáng)的抑制,楊亞晉等[30]研究的乳酸菌素BSN4和BCN4對(duì)G-菌和真菌都無(wú)明顯抑制效果。食品防腐中通常幾種不同的防腐劑聯(lián)合使用從而達(dá)到滿意的抑菌效果。BaC21可以作為一種天然的生物防腐劑和nisin等細(xì)菌素聯(lián)合使用,發(fā)揮互補(bǔ)抗菌效果。

表1 BaC21對(duì)各指示菌抑制作用Table 1 The antibacterial spectrum of bacteriocin against various indicator strains
2.5 酶及物理化學(xué)處理對(duì)細(xì)菌素抗菌活性影響
酶及各物理化學(xué)因素處理結(jié)果如表2所示。在不同蛋白酶處理后,BaC21抗菌活性完全喪失,同對(duì)照相比差異顯著(P<0.05),脂肪酶和α-淀粉酶處理后抑菌活性無(wú)顯著變化(P>0.05)。表明菌株XbC-36所產(chǎn)抗菌物質(zhì)具有蛋白質(zhì)特性。另一類重要的天然抗菌物質(zhì)是主要來(lái)源于芽孢桿菌的脂肽,因結(jié)構(gòu)中存在非正常氨基酸構(gòu)成的環(huán)狀肽鏈而對(duì)各種蛋白酶不敏感。BaC21經(jīng)所試酶處理后的活性表現(xiàn)說明其分子中不存在脂肪、淀粉鏈,也不存在非正常的氨基酸鏈,這與所報(bào)道的其他細(xì)菌素一致,故可判斷菌株XbC-36所產(chǎn)抗菌物質(zhì)同為細(xì)菌素。BaC21蛋白質(zhì)性質(zhì)意味著它在食品中可以作為天然防腐劑抑制食源性致病菌、腐敗菌的生長(zhǎng),當(dāng)其同食品一起被食用后可在人胃腸道中降解并作為營(yíng)養(yǎng)素被人吸收。
細(xì)菌素從發(fā)酵液中沉淀分離、高分子吸附介質(zhì)層析、疏水層析及HPLC純化時(shí)經(jīng)常用到不同有機(jī)溶劑沉淀或洗脫,有必要分析檢測(cè)其有機(jī)溶劑穩(wěn)定性。用丙酮、乙腈、乙醇和異丙醇等有機(jī)溶劑對(duì)BaC21處理,其抗菌活性無(wú)明顯下降(P>0.05)。證實(shí)BaC21分子中不存在與抗菌活性有顯著影響的易溶于有機(jī)溶劑的脂質(zhì)分子[31],因此,BaC21可以應(yīng)用于高脂肪食品的保鮮。
表面活性劑吐溫20、吐溫80、Triton 100對(duì)BaC21抗菌活性無(wú)顯著影響(P>0.05);而SDS能夠增強(qiáng)抑菌活性,推測(cè)可能是SDS改變了細(xì)胞膜的通透性,與此相似的報(bào)道還有Lysinibacillussp. JX402121細(xì)菌素[32]、Lactococcuslactissubsp. lactis 2MT細(xì)菌素nisin[22]。
BaC21在pH 2~7時(shí),穩(wěn)定性最佳,抑菌活性基本無(wú)明顯變化(P>0.05);在pH 9.0時(shí),穩(wěn)定性有所下降,但其抑菌活性仍保持60%以上,同對(duì)照差異顯著(P<0.05)。BaC21在酸性條件下表現(xiàn)出更佳的穩(wěn)定性,這與報(bào)道的大部分細(xì)菌素相似,如HU等[5]從LactobacillusalimentariusFM-MM4分離的細(xì)菌素MM4在pH 7.0以上穩(wěn)定性大幅降低;羅怡等[33]研究的乳酸菌素在堿性條件下完全失活;WAYAH等[1]研究的LactobacillussalivariusSPW1細(xì)菌素在pH 8.0以上穩(wěn)定性變差。在強(qiáng)堿性條件下的不穩(wěn)定性可能是因?yàn)闃O端的堿性pH條件下,分子間強(qiáng)靜電作用導(dǎo)致多肽氨基和羧基解離,最終導(dǎo)致蛋白質(zhì)變性抑菌活性喪失[27]。
BaC21在100 ℃以下30 min內(nèi)活性基本無(wú)變化(P>0.05),但當(dāng)超過100 ℃之后穩(wěn)定性下降,特別是在121 ℃、15 min穩(wěn)定性下降明顯(P<0.05),其抑菌活性保留55.2%。整體說明BaC21具有優(yōu)良的熱穩(wěn)定性。在食品中使用時(shí)可以耐受食品加工工藝的熱處理過程。

表2 細(xì)菌素BaC21穩(wěn)定性分析Table 2 Stability analysis of bacteriocin BaC21
2.6 細(xì)菌素對(duì)S. aureus和E. coli的抑菌模式
BaC21對(duì)E.coli和S.aureus的抑菌情況如圖5所示,在不添加BaC21情況下,兩指示菌株呈現(xiàn)典型的細(xì)菌生長(zhǎng)特征,2 h前為延遲期,2~8 h呈指數(shù)生長(zhǎng),8~24 h逐漸從指數(shù)期過渡到穩(wěn)定期。而當(dāng)在培養(yǎng)4 h加入BaC21后,兩指示菌生長(zhǎng)顯示立即受到抑制,吸光度逐漸降低,培養(yǎng)24 h兩者OD值分別為0.301及0.101。由此可知BaC21對(duì)S.aureus和E.coli不僅具有明顯的抑制作用同時(shí)還可使細(xì)胞裂解。
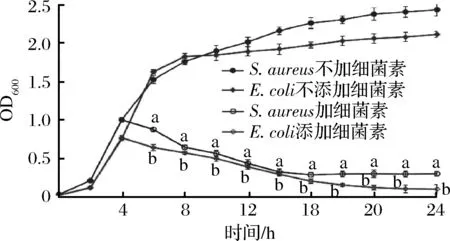
圖5 BaC21對(duì)S. aureus和E. coli生長(zhǎng)的影響Fig.5 Effect of bacteriocin on the growth of S.aureus and E.coli注:小寫字母表示同對(duì)照組間顯著差異(P<0.05)
3 結(jié)論
從L.caseiXbC-36發(fā)酵液中通過兩步純化即分子篩凝膠過濾層析和RP-HPLC純化到單一組分的BaC21,其分子質(zhì)量與報(bào)道的其他細(xì)菌素不同。BaC21在酸性、中性及弱堿性環(huán)境下有良好的穩(wěn)定性,并可耐受100 ℃的熱處理,對(duì)有機(jī)溶劑、表面活性劑耐受性好。對(duì)G+菌和G-菌都有廣譜抑菌活性,特別是對(duì)G-菌如銅綠假單胞菌、大腸桿菌、腸沙門氏菌、副溶血性弧菌等表現(xiàn)出更為顯著的抑菌活性。經(jīng)多種蛋白酶處理后完全失活,但對(duì)脂肪酶和淀粉酶不敏感。S.aureus和E.coli在培養(yǎng)中添加BaC21,菌體生長(zhǎng)受到完全抑制。綜上結(jié)果表明,BaC21有潛力用做天然食品防腐劑,可以和nisin等抗菌劑聯(lián)合使用提高食品的質(zhì)量和安全。
