產蛋白酶菌株的鑒定及其對蝦殼的脫蛋白研究
2020-09-28 06:41:34李永成
中國釀造 2020年9期
張 巧,黃 興,李永成*
(1.海南大學 食品科學與工程學院,海南 海口 570228;2.賀州學院食品科學與工程技術研究院,廣西 賀州 542899)
近年來,隨著我國水產養殖業的發展,蝦的產量和消費量不斷增長。工業上,蝦主要加工成蝦仁用于出口,產生的蝦頭、蝦殼、蝦尾等廢棄物約占整蝦質量的40%左右[1]。這些蝦類廢棄物如果不經適當的加工處理,不僅會產生難聞的氣味,給環境帶來嚴重污染,蝦料中豐富的蛋白質、礦物質、蝦青素和甲殼素等營養物質或生物材料也會白白浪費[2]。甲殼素是一種由N-乙酰-D-氨基葡萄糖通過β-1,4-糖苷鍵連接而成的聚合物,在自然界中的含量僅次于纖維素。從甲殼類廢棄物中提取的甲殼素及其脫乙酰化殼聚糖在食品、醫藥、納米材料、水處理、化妝品、環境等領域有廣泛的應用[3-4]。
在蝦殼廢棄物中,甲殼素的含量約占20%~30%,蛋白質的含量約占20%~30%,礦物質的含量約占30%~40%[5]。蝦殼甲殼素的提取,主要有脫蛋白和脫鹽兩個關鍵步驟。甲殼素提取方法分為化學法和生物法。化學法采用強酸將蝦殼中難溶于水的鹽類物質變成易溶于水的無機鹽,從而達到去除蝦殼中礦物質的目的,再使用強堿脫除蝦殼中的蛋白質[6]。化學法提取甲殼素過程中,產生的大量酸堿廢水難以處理,且強酸強堿還可能造成甲殼素發生部分解聚,影響其品質。而生物催化、生物轉化和發酵法制備甲殼素的反應條件溫和,且相同條件下制備的甲殼素的品質較為穩定[7],近年來,生物法制備甲殼素已逐步取代化學法,得到了科研工作者的廣泛探究。利用微生物發酵產蛋白水解酶對蝦殼進行脫蛋白,不僅是生物法制備甲殼素的重要過程,還能回收蝦殼中的蛋白質[8]。發酵脫蛋白的微生物以芽孢桿菌居多,如地衣芽孢桿菌、枯草芽孢桿菌、蠟樣芽孢桿菌、短芽孢桿菌等,對甲殼類廢棄物的脫蛋白率均在75%以上[9-10]。其中蠟樣芽孢桿菌是一種具有較高蛋白酶活性的微生物,郝凱[11]從凡納濱對蝦的腸道中分離出氣單胞菌屬、芽孢桿菌屬、弧菌屬等微生物,其中產蛋白酶活性最高的菌株是蠟樣芽孢桿菌。目前,發酵法脫蛋白仍存在發酵時間長、操作復雜、脫蛋白效率較低等問題。對蝦殼蛋白質脫除能力強的微生物,在甲殼素制備工藝中具有較好的應用價值。本研究從蝦塘沉積物中分離并鑒定一株產蛋白酶的細菌,研究該菌株與枯草芽孢桿菌在不同條件下的脫蛋白作用,為微生物發酵法提取蝦殼甲殼素提供較好的脫蛋白菌株。
1 材料與方法
1.1 材料與試劑
1.1.1 材料和菌株
南美白對蝦蝦殼:海南照豐水產有限公司;枯草芽孢桿菌(Bacillus subtilis)DX-2:本實驗室保存。
1.1.2 化學試劑
酵母提取粉、胰蛋白胨(均為生化試劑):廣東環凱生物科技有限公司;福林酚試劑:北京索萊寶科技有限公司。
1.1.3 培養基
營養瓊脂(nutrient agar,NA):廣東環凱生物科技有限公司。
篩選培養基:1%蝦殼粉、5%葡萄糖、0.1%酵母膏、0.8%NaCl、0.15% CaCl2、0.07%K2HPO3、0.03% KH2PO3、0.05%MgSO4、2%瓊脂,蒸餾水定容至1 000 mL。
蝦殼發酵培養基:5%蝦殼粉、5%葡萄糖,蒸餾水定容至1 000 mL。
Luria-Bertani(LB)肉湯培養基:0.5%酵母粉、1%胰蛋白胨、1%NaCl,pH 7.0。
以上培養基滅菌條件:121 ℃高壓滅菌15 min。
1.2 儀器與設備
ZQLY-180N立式振蕩培養箱:海知楚儀器有限公司;SH220F石墨消解儀:濟南海能儀器股份有限公司;T6新世紀紫外可見分光光度計:北京普析通用儀器有限責任公司。
1.3 實驗方法
1.3.1 蝦殼預處理
將去雜后的蝦殼用水清洗干凈,自然晾干后粉碎,過40目篩網,獲得蝦殼粉(蛋白質含量為30%左右),密封保存備用。
1.3.2 產蛋白酶菌株的分離
從蝦塘沉積物中取樣10 g,加入到裝有90 mL無菌生理鹽水的三角瓶中,攪拌均勻后,超凈工作臺中靜置10 min。取上清液,用無菌水稀釋至10-3、10-4、10-5g/mL。吸入1 mL的上述稀釋液至倒有篩選培養基的無菌培養皿表面,涂布均勻,35 ℃倒置培養1~2 d,觀察菌落生長情況。
挑取生長良好的單一菌落,接種至LB肉湯培養基中,35 ℃、180 r/min培養1 d。培養好的菌液在倒有NA培養基的培養皿中劃線分離,獲得純菌株。
1.3.3 菌株的鑒定
(1)菌落特征和菌體形態
將分離到的純菌株劃線于NA培養基表面,35 ℃培養1 d,觀察菌落形態,并進行革蘭氏染色[12]。
(2)生理生化特性
參照《常見細菌系統鑒定手冊》[13],對分離到的具有較高蛋白酶活性的細菌進行生理生化特征的測定。
(3)分子生物學鑒定
通過測定菌株的16S rDNA基因序列進行分子生物學鑒定,具體參照潘曉倩等[14]的方法并稍作修改。按照細菌脫氧核糖核酸(deoxyribonucleic acid,DNA)提取試劑盒上的具體方法從分離到的純菌株菌液中提取DNA,以提取的DNA作為模板,采用通用引物27F(5'-AGAGTTTGATCMTGGCTCAG-3')和1492R(5'-TACGGYTACCTTGTTACGACTT-3')進行聚合酶鏈式反應(polymerase chain reaction,PCR)。擴增產物委托上海派諾生物技術股份有限公司進行測序。將測得的序列結果與NCBI GenBank中序列進行BLAST比對,通過MEGA6.06軟件構建系統發育樹。
1.3.4 不同培養基組成對蝦殼脫蛋白率的影響
(1)葡萄糖添加量對蝦殼脫蛋白率的影響
將篩選菌株和枯草芽孢桿菌分別培養至OD600nm=1.0時,按照3%的接種量接種至蝦殼發酵培養基中(裝液量20 mL/100 mL),葡萄糖添加量分別為10 g/L、30 g/L、50 g/L、70 g/L、90 g/L,35 ℃、180 r/min發酵5 d,測定發酵液中蝦殼的脫蛋白率。
(2)蝦殼粉添加量對蝦殼脫蛋白率的影響
在葡萄糖添加量優化的基礎上,蝦殼粉添加量分別為10 g/L、20 g/L、30 g/L、40 g/L、50 g/L,35 ℃、180 r/min發酵5 d,測定發酵液中蝦殼的脫蛋白率。
(3)酵母膏添加量對蝦殼脫蛋白率的影響
在葡萄糖和蝦殼粉添加量優化的基礎上,酵母膏添加量分別為0、1 g/L、2 g/L、3 g/L、4 g/L、5 g/L,35 ℃、180 r/min發酵5 d,測定發酵液中蝦殼的脫蛋白率。
1.3.5 發酵時間對蝦殼脫蛋白的影響
在發酵培養基優化的基礎上,按照3%的接種量,35 ℃、180 r/min分別發酵1 d、2 d、3 d、4 d、5 d、6 d、7 d,測定發酵液中蛋白酶活力和脫蛋白率的變化。
1.3.6 蛋白酶活力的測定
吸取一定量的發酵液,參照GB/T 23527—2009《蛋白酶制劑》測定蛋白酶的活性。蛋白酶酶活定義:在一定反應溫度和時間條件下,1 mL發酵液在1 min內水解酪蛋白產生1 μg酪氨酸,為1個蛋白酶活力單位(U)。
1.3.7 蝦殼粉脫蛋白率的測定
發酵結束后,取出三角瓶,抽濾,將沉淀水洗至上清液的OD600nm為0,獲得的蝦殼粉在60 ℃干燥箱內干燥24 h,參照GB 5009.5—2016《食品中蛋白質的測定》中的分光光度法測定總氮和蛋白質含量。按照如下公式計算蝦殼發酵后的脫蛋白率:
其中:m0為三角瓶中原蝦殼粉的質量,g;N0為原蝦殼粉的含氮量,%;m為發酵后三角瓶中蝦殼粉的質量,g;N為發酵后蝦殼粉的含氮量,%;p為蝦殼粉的蛋白質含量,%。
2 結果與分析
2.1 產蛋白酶菌株的鑒定
2.1.1 菌株形態特征
從蝦塘中分離到的產蛋白酶菌株C1,通過平板劃線得到的單菌落呈圓形,奶油色,不透明,且邊緣粗糙。進一步通過革蘭氏染色,顯微鏡下觀察的形態見圖1,結果表明,該菌株呈桿狀,判斷菌株C1是一株革蘭氏陽性桿菌。
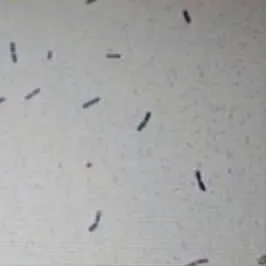
圖1 菌株C1的細胞形態Fig.1 Cell morphology of strain C1
2.1.2 生理生化鑒定
菌株C1的生理生化特性及對碳源的利用能力見表1所示。該菌株具有使明膠液化、牛奶胨化的能力,具有淀粉酶活性,VP試驗、甲基紅(methyl red,MR)試驗呈陽性,但不能利用檸檬酸鈉,最適溫度為37 ℃,最適pH為7.5。菌株C1對葡萄糖、果糖、乳糖等大部分單糖、雙糖或糖醇均能較好地利用,但不能以纖維素作為碳源。
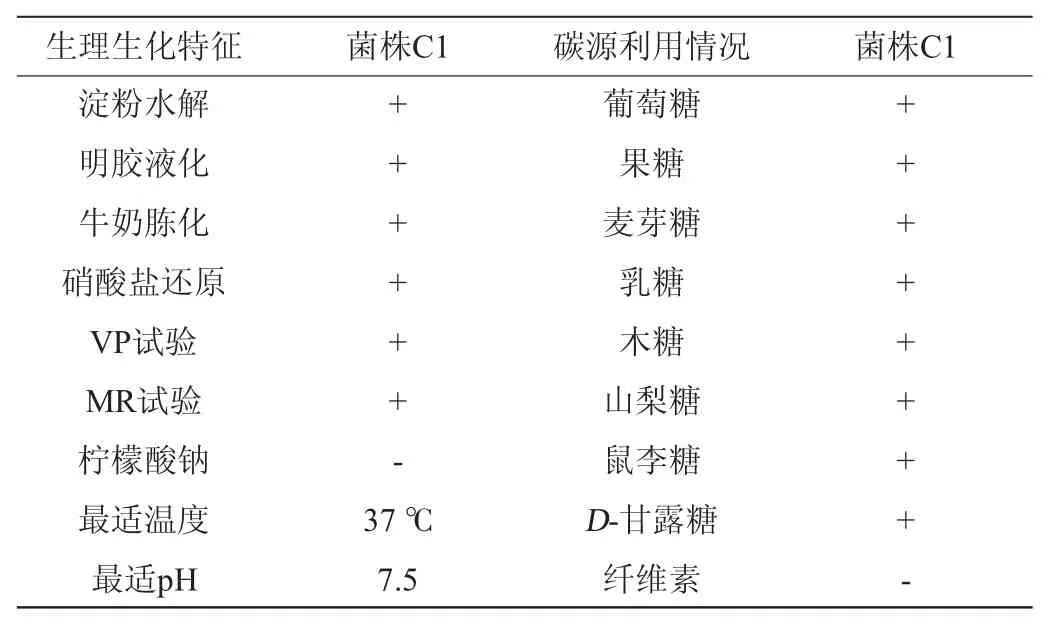
表1 菌株C1的生理生化特征及碳源利用情況Table 1 Physiological and biochemical characteristics and carbon source utilization ability of strain C1
2.1.3 分子生物學鑒定
菌株C1的PCR擴增產物采用1%瓊脂糖凝膠電泳進行純度及擴增片段大小的檢測,結果見圖2。由圖2可知,菌株C1僅在1 500 bp附近顯示一條明顯的條帶,說明擴增產物的純度較高,菌株C1的16S rDNA被成功進行PCR擴增。
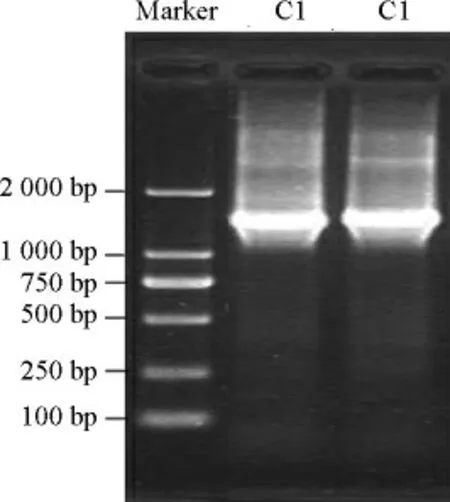
圖2 菌株C1的16S rDNA PCR擴增產物電泳圖Fig.2 Electrophoresis patterns of PCR amplification products of 16S rDNA for strain C1
將待鑒定菌株的16S rDNA序列與NCBI中相似度高的已知序列構建成系統發育樹,若兩者的16S rDNA同源性達到99%,則認為它們是同一種菌[15]。菌株C1的16S rDNA系統發育樹見圖3所示。由圖3可知,菌株C1與Bacillus cereus的親緣關系最近,同源性達到99%,結合菌落形態和生理生化特征的結果,可確定菌株C1為一株蠟樣芽孢桿菌(Bacillus cereus)。蠟樣芽孢桿菌因其高蛋白酶活性具有一定的應用價值。于平等[16]從土壤中篩選的蠟樣芽孢桿菌,能產中性蛋白酶,且耐熱性較好,應用前景廣闊。吳昱含等[17]從廢鉻污染的土壤中分離到一株產蛋白酶耐鉻的蠟樣芽孢桿菌,該菌株能有效清除含鉻污水中的蛋白質,清除率高達91.9%,達到制革污水清潔化的目的。

圖3 基于16S rDNA序列菌株C1的系統發育樹Fig.3 Phylogenic trees of strain C1 based on 16S rDNA sequences
2.2 不同培養基組成對蝦殼脫蛋白率的影響
2.2.1 葡萄糖添加量對蝦殼脫蛋白率的影響
蝦殼中的蛋白質能為微生物的生長提供氮源,因此蝦殼發酵培養基需要添加適當的碳源。葡萄糖添加量對蠟樣芽孢桿菌和枯草芽孢桿菌發酵蝦殼的脫蛋白率的影響見圖4所示。由圖4可知,蝦殼發酵培養基中葡萄糖添加量為10~50g/L時,增加葡萄糖含量有利于蝦殼中蛋白質的溶出,從而提高蝦殼的脫蛋白率。當葡萄糖添加量為50~90g/L時,繼續增加葡萄糖添加量,2株芽孢桿菌發酵蝦殼的脫蛋白率反而下降。因此,對蠟樣芽孢桿菌和枯草芽孢桿菌而言,蝦殼發酵培養基中葡萄糖的添加量確定為50 g/L。此外,圖4表明蠟樣芽孢桿菌發酵蝦殼的脫蛋白能力高于枯草芽孢桿菌,在蝦殼發酵培養基中添加50 g/L的葡萄糖,蠟樣芽孢桿菌和枯草芽孢桿菌的脫蛋白率分別為80.4%、75.6%。
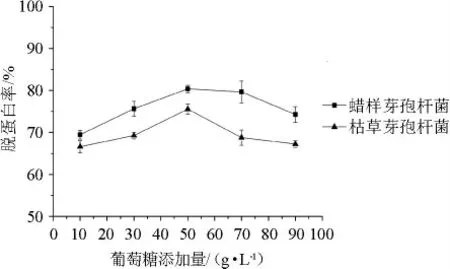
圖4 葡萄糖添加量對蝦殼脫蛋白率的影響Fig.4 Effect of glucose addition on the deproteinization rate of shrimp shell
2.2.2 蝦殼粉添加量對蝦殼脫蛋白率的影響
發酵培養基中蝦殼粉添加量對蠟樣芽孢桿菌和枯草芽孢桿菌發酵蝦殼的脫蛋白率的影響見圖5所示。由圖5可知,蝦殼粉添加量為20 g/L時,2株芽孢桿菌發酵蝦殼的脫蛋白率較高,分別達到85.1%、78.4%,蝦殼粉添加量為10~50 g/L時,蠟樣芽孢桿菌發酵蝦殼的脫蛋白率均高于枯草芽孢桿菌。蝦殼粉添加量高于20 g/L時,枯草芽孢桿菌發酵的脫蛋白率下降速率較大。因此,2株微生物的發酵培養基中,蝦殼粉添加量確定為20 g/L。

圖5 蝦殼粉添加量對蝦殼脫蛋白率的影響Fig.5 Effect of shrimp shell power addition on the deproteinization rate of shrimp shell
2.2.3 酵母膏添加量對蝦殼脫蛋白率的影響
酵母膏中含有蛋白質、維生素和必需氨基酸等營養物質,是一種非常理想的微生物發酵原料[18]。在發酵初期,蝦殼發酵培養基中能供微生物利用的可溶性蛋白質含量較低,不利于微生物生長。而添加一定量的酵母膏可能促進微生物的生長和脫蛋白作用。酵母膏添加量對蠟樣芽孢桿菌和枯草芽孢桿菌發酵蝦殼的脫蛋白率的影響見圖6。
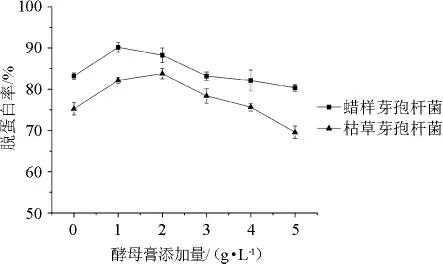
圖6 酵母膏添加量對蝦殼脫蛋白率的影響Fig.6 Effect of yeast extract addition on the deproteinization rate of shrimp shell
由圖6可知,添加0~5 g/L的酵母膏,蠟樣芽孢桿菌對蝦殼的脫蛋白率均高于枯草芽孢桿菌,且酵母膏添加量為1 g/L時,蠟樣芽孢桿菌發酵蝦殼的脫蛋白率達到最高(90.1%)。在此基礎上增加酵母膏添加量,脫蛋白率不斷下降,可能原因在于較高含量的酵母膏雖然有利于微生物的生長,但是產蛋白酶能力下降。因此確定酵母膏添加量為1 g/L。
2.3 蝦殼發酵過程中蛋白酶活力及脫蛋白率的變化
枯草芽孢桿菌和蠟樣芽孢桿菌發酵蝦殼過程中,發酵液中蛋白酶活力及蝦殼脫蛋白率的變化見圖7所示。
由圖7(a)可知,2株菌的酶活力均呈現先升高后下降的趨勢,在發酵0~4 d,枯草芽孢桿菌的酶活力稍高,4 d后蠟樣芽孢桿菌的酶活力較高。枯草芽孢桿菌在發酵第4天達到最高酶活力154.6 U/mL,而蠟樣芽孢桿菌在發酵第5天的酶活力最高,達到220.8 U/mL,此時枯草芽孢桿菌的酶活力僅為145.7 U/mL。
由圖7(b)可知,在蛋白酶的作用下,2株菌均對蝦殼的脫蛋白率不斷增加,0~5 d增加較快,5 d后緩慢增長。發酵第5天,枯草芽孢桿菌和蠟樣芽孢桿菌對蝦殼的脫蛋白率分別達到80.4%、90.8%。蠟樣芽孢桿菌的脫蛋白作用明顯強于枯草芽孢桿菌。通過微生物發酵提取蝦殼中的甲殼素,脫蛋白率較高可以達到90%~95%[9],如灰色鏈霉菌發酵過程中產的蛋白酶對凡納濱對蝦蝦殼進行脫蛋白作用,脫蛋白率為91.1%[19];通過沙雷氏菌發酵處理南美白對蝦蝦殼,脫蛋白率達到94.5%[20]。
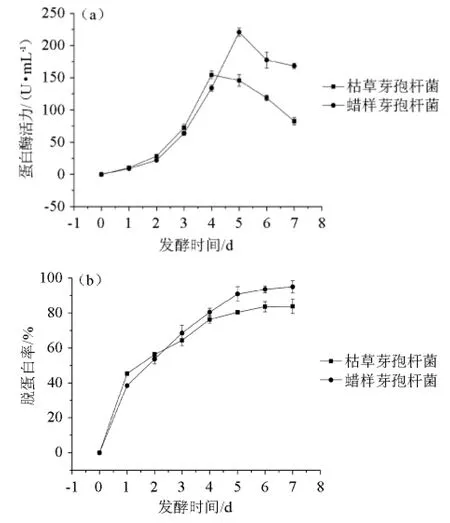
圖7 蝦殼發酵過程中蛋白酶活力(a)和脫蛋白率(b)的變化Fig.7 Variation of protease activity (a) and deproteinization rate (b)during shrimp shell fermentation process
3 結論
從蝦塘沉積物中分離到一株產蛋白酶的細菌C1,經鑒定其為蠟樣芽孢桿菌(Bacillus cereus)。與枯草芽孢桿菌相比,本研究篩選到的蠟樣芽孢桿菌對蝦殼的脫蛋白能力更強。當蝦殼發酵培養基中葡萄糖添加量為50 g/L時,蝦殼粉20 g/L,酵母膏1 g/L,蠟樣芽孢桿菌發酵蝦殼的脫蛋白率較高;此條件下經蠟樣芽孢桿菌發酵5 d,發酵液中的蛋白酶活力達到220.8 U/mL,蝦殼脫蛋白率達到90.8%,而同等條件下枯草芽孢桿菌的脫蛋白率僅為80.4%。本研究篩選的蠟樣芽孢桿菌,在發酵過程中能產生高活性的蛋白酶,高效脫除蝦殼中的蛋白質,在蝦殼甲殼素的生物提取工藝中具有較好的應用前景。
