基于線粒體能量代謝途徑的金針菇采后納米包裝保鮮機制
2020-09-11 05:37:26馬寧王鶴潼方東路趙立艷楊文建裴斐胡秋輝
中國農業科學 2020年16期
馬寧,王鶴潼,方東路,趙立艷,楊文建,裴斐,胡秋輝
(1南京財經大學食品科學與工程學院/江蘇省現代糧食流通與安全協同創新中心/江蘇省糧油品質控制及深加工技術重點實驗室,南京 210023;2南京農業大學食品科技學院,南京 210095)
0 引言
【研究意義】金針菇(Flammulina filiformis)又稱毛柄金錢菌,由于其美味和高營養特性,目前已成為世界第三大食用菌,不僅銷售量逐年增長,更有廣闊的開發前景[1-2]。然而采后金針菇含水量高、呼吸作用強、能量代謝旺盛[3],極易發生開傘、褐變和腐爛等品質劣變現象[4]。目前對于金針菇采后品質劣變的研究大多數集中于失重率、褐變度、開傘率等表觀指標,內在機理的研究尚缺乏,而線粒體是植物能量代謝及能量物質(adenosine triphosphate,ATP)生成的主要場所。有研究表明,能量狀態失衡是果蔬品質劣變的核心因素[5],由此推測金針菇采后品質劣變也可能是由能量狀態失衡引起[3],因此,基于線粒體探究金針菇冷藏過程的能量狀態很有必要,對金針菇保鮮技術體系研究具有重要意義。【前人研究進展】目前國內外對線粒體提取方法的研究有很多,主要有差速離心法[6-7]、不連續密度梯度離心法[8]及試劑盒法(如酵母試劑盒法[9]、普利萊試劑盒法[10])。密克等[8]利用蔗糖密度梯度離心法提取板栗疫病菌的線粒體,通過測定發現所制備的線粒體完整性好,且受到污染較少。ZHAO等[9]采用普利萊試劑盒提取的線粒體研究在大鼠中內質網和線粒體是否共同參與椎間盤突出的細胞凋亡和椎間盤退變。GUIROLA等[10]利用酵母線粒體提取試劑盒提取線粒體,進而研究釀酒酵母中線粒體內微量物質pif1對細胞內蛋白及微量元素的影響。納米材料近年來在食品包裝的應用研究中越來越受關注,常見的有金屬及其氧化物納米材料、二氧化硅及硅酸鹽納米材料、多糖基納米材料、蛋白基納米材料和復合納米材料[11]。在實驗室前期的研究中開發了一種添加了納米銀、納米二氧化鈦、納米凹凸棒土、納米二氧化硅的復合包裝材料[12],該包裝材料能有效抑制金針菇開傘、褐變、孢子增加等品質劣變,保鮮效果顯著[12-13]。研究表明,采后果蔬的質量下降與能量供應密切相關[14-15]。課題組前期對乙烯含量、活性氧積累和線粒體微觀結構等進行了研究,這些都與能量代謝有關[13,16]。另外對蛋白組學的檢測發現納米包裝組金針菇中參與碳水化合物代謝的相關蛋白在冷藏期間發生了顯著性差異表達[17]。研究發現ATP供應不足或生產效率下降會導致膜結構完整性破壞,并導致果蔬采后迅速衰老和質量下降[18-20]。闞娟等[21]報道線粒體復合體活性的下降也會導致以上的后果,并抑制能量物質合成,從而導致細胞能量供應不足。【本研究切入點】線粒體的研究(包括提取純化)目前均集中在人、動物或植物上,應用于金針菇的幾乎沒有。【擬解決的關鍵問題】通過對常見線粒體提取方法的對比,確定適用于金針菇線粒體的最優提取方法。同時,從線粒體角度闡明不同包裝對金針菇能量代謝的影響。此外,通過動態檢測金針菇在冷藏期間ATP代謝系統物質含量變化,探明納米包裝在貯藏中維持金針菇能量代謝水平的作用機制。
1 材料與方法
試驗于2019年1—11月在南京財經大學食品科學與工程學院進行。
1.1 試驗材料
新鮮金針菇(F-C01l)從南京金萬辰生物科技有限公司采購,1 h內低溫運輸至實驗室。篩選的金針菇大小為菌蓋直徑3—4 cm、重量為23—30 g、未開傘、沒有機械損傷。挑選后的金針菇進行液氮研磨,并用于后續線粒體提取方法篩選優化。金針菇菌種由同公司提供同品種菌種,培養至菌絲體時對菌絲進行過濾沖洗,將培養基沖洗干凈后進行凍干處理,最后使用液氮研磨供后續線粒體提取方法篩選優化。
1.2 儀器與試劑
CL21R型微量臺式冷凍離心機,德國 Thermo Electron LED公司;SX500型快速自動高壓滅菌儀,日本鹿兒島地震保險公司;HH2型數顯恒溫水浴鍋,國華電器有限公司;Axlo Vert.A1型倒置式顯微鏡,德國卡爾蔡司股份公司;THZ-C型恒溫振蕩器,上海璽袁科學儀器有限公司;FreeZone 2.5升臺式凍干機,北京照生行儀器設備有限公司;日立 JEM-1011型透射電子顯微鏡,日本Hitachi公司;Clark氧電極,漢莎科學儀器有限公司;ELX800型酶標儀,美國Bio-Tek公司;1260系列高效液相色譜儀,美國Agilent公司。
乙二胺四乙酸二鈉試劑(EDTA)、甘露醇、Tris-HCl試劑、牛血清蛋白(BSA)、蔗糖、β-巰基乙醇、苯甲基黃酰氟(PMSF)、健那綠、麥芽糖、葡萄糖、酵母提取物、蛋白胨、MgSO4·7H2O、KH2PO4、瓊脂粉、馬鈴薯葡萄糖瓊脂培養基(PDA)、Gluta固定液、二甲砷酸鈉緩沖液、鋨酸、丙酮、Epon-812環氧樹脂、磷酸氫二鉀、磷酸二氫鉀、甲醛(分析純)、三磷酸腺苷、二磷酸腺苷、一磷酸腺苷、乙醇脫氫酶(ADH)活性檢測試劑盒、線粒體呼吸鏈復合體Ⅳ活性檢測試劑盒、超氧化物歧化酶(SOD)活性檢測試劑盒、線粒體呼吸鏈復合體I活性檢測試劑盒、線粒體呼吸鏈復合體 III活性檢測試劑盒均采購于北京索萊寶有限公司;酵母線粒體提取試劑盒采購于北京百奧萊博科技有限公司;線粒體-胞漿蛋白制備試劑盒采購于北京普利萊基因技術有限公司。
1.3 試驗設計
1.3.1 線粒體的提取試驗 改進普利萊法(Improved Pulilai Kit Method,IPKM):由于傳統普利萊法沒有對于細胞壁破碎的過程,更適用于動物細胞,因此根據吳媛等[22]的方法改進,先對金針菇細胞進行破壁處理,再進行線粒體的提取。具體步驟如下:將樣品浸沒至巰基化合物中,在32℃下靜置15—30 min。10 000 r/min離心1 min,棄上清,沉淀重懸于山梨醇,每克樣品加入蝸牛酶30—40 mg,37℃保溫1 h。800×g離心5 min,將上清液丟棄,沉淀物用PBS洗3次,并于4℃、800×g離心5—10 min收集沉淀。加入1.5 mL冰預冷的Mito-Cyto Buffer重懸沉淀,4℃、800×g離心5 min,收集上清再次800×g離心5 min,收集上清后12 000×g再次離心10 min,管底沉淀即為線粒體。
不連續密度梯度離心法(discontinuous density gradient centrifugation method,DDGCM):參照藺凱麗等[23]的方法,將樣品提取液以 1﹕6的比例向樣品中加入4℃預冷提取液(內含1 mmol·L-1EDTA、0.3 mol·L-1甘露醇、50 mmol·L-1Tris-HCl、0.2% BSA、2 mmol·L-1β-巰基乙醇),在組織勻漿機中勻漿3次,15 s/次。將勻漿液1 500×g離心5 min,取上清液3 000×g離心5 min,再取上清液12 000×g離心15 min,沉淀為粗線粒體。將沉淀分別用3種濃度的2 mL洗滌液洗滌(由于洗滌液的濃度高低影響線粒體的產量,因此采用3種濃度的洗滌液,并最終取最佳的一組與另兩種方法組進行比較),首先配置初始濃度洗滌液:內含 1 mmol·L-1EDTA、0.3 mol·L-1甘露醇、10 mmol·L-1Tris-HCl、0.5 mmol·L-1PMSF,然后將上述濃度洗滌液分別稀釋200倍和400倍,從而得到3個濃度的洗滌液,以下分別用 DDGCM-1、DDGCM-200、DDGCM-400來表示。洗滌過后12 000×g離心15 min,得沉淀為粗線粒體。將粗線粒體用懸浮液(內含1 mmol·L-1EDTA、0.3 mol·L-1甘露醇、10 mmol·L-1Tris-HCl)懸浮。將粗線粒體懸浮液小心鋪在 15%﹕23%﹕32%﹕60%蔗糖不連續密度梯度上。4℃下134 000×g離心45 min。取32%和60%之間不透明淺白色線粒體環,用2倍體積懸浮液洗滌。4℃下12 000×g離心20 min,得到的沉淀即為高純度線粒體。
酵母線粒體提取試劑盒方法(yeast mitochondrial extraction kit method,YMEKM):參照北京百奧萊博科技有限公司提供的酵母線粒體提取試劑盒說明書進行提取。
1.3.2 不同包裝袋貯藏保鮮試驗 冷藏試驗中將金針菇在(4±1)℃和90%相對濕度下于黑暗中預冷24 h,并將樣品(250±5 g)隨機包裝到納米包裝袋和普通PE包裝袋中進行貯藏,每個處理組3個平行樣品,對各包裝組金針菇分別進行21袋的包裝處理。然后將樣品在(4±1)℃和90%相對濕度下存儲21 d。冷藏期間每3 d利用優化得到的方法對各處理組樣品進行線粒體提取,并進行生理生化分析。本研究中,所用的納米包裝袋是參考筆者實驗室FANG等[24]的生產工藝得到。按質量比取納米粉體15%(其中納米銀30%、納米二氧化鈦35%、納米凹凸棒土 25%、納米二氧化硅 10%)、低密度聚乙烯46%、低密度線性聚乙烯22%、分散劑10%、潤滑劑5%和偶聯劑2%制得納米母粒,并按質量比取經上述制備的納米母粒7.5%、防霧劑母粒8%和低密度線性聚乙烯84.5%制得納米保鮮膜。普通PE包裝袋是沒有納米復合母料納米粉的聚乙烯袋,其尺寸和厚度(25 cm×25 cm,40 μm),與納米包裝袋相同。
1.4 檢測方法
1.4.1 線粒體結構完整性的檢測 乙醇脫氫酶(ADH)活性的測定:參照ADH活性檢測試劑盒說明書進行測定。
細胞色素C氧化酶活性的測定:參照線粒體呼吸鏈復合體Ⅳ活性檢測試劑盒進行測定。
呼吸速率的測定:參照高永彬[25]的方法,使用Clark(Hansatech Ltd, Hardwick industrial Estate, King’s Lynn, Norfold, UK)公司的氧電極裝置分別對各組提取的線粒體、金針菇子實體以及菌絲體進行呼吸氧消耗的測定,測定過程中保持室內 25℃恒溫狀態。(1)線粒體呼吸速率的測定:測定前,用反應液進行校正,待溫度恒定。取100 μL線粒體懸浮液定容到1 mL反應液中(反應液內含0.3 mol·L-1甘露醇、2 mmol·L-1MgSO4、5 mmol·L-1KH2PO4、10 mmol·L-1TES、10 mmol·L-1NaCl、0.1%(w/v)BSA,加入終濃度為 10 mmol·L-1琥珀酸鹽、5 mmol·L-1抗壞血酸、10 μmol·L-1ATP、0.5 mmol·L-1ADP、1 mmol·L-1NADH、25 μmol·L-1細胞色素 C 等底物和 0.05%(w/v)曲拉通 100),并迅速轉移至反應室中。開始測定,待耗氧速率穩定時開始記錄。(2)金針菇子實體呼吸速率的測定:測定前,用超純水進行校正,待溫度恒定。取兩個菌蓋直徑為0.3 cm,菌柄長1.5 cm的金針菇子實體加入反應室并使反應室內超純水定容到 1 mL,開始測定,待耗氧速率穩定時開始記錄。(3)金針菇菌絲體呼吸速率的測定:測定前,用CYM培養基(內含1%麥芽糖、2%葡萄糖、0.2%酵母提取物、0.2%蛋白胨、0.05% MgSO4·7H2O、0.46% KH2PO4)進行校正,待溫度恒定。取0.5 mL菌絲體定容到1mL培養基中,并迅速轉移至反應室中。開始測定,待耗氧速率穩定時開始記錄。
透射電鏡觀察:參照TARZE等[26]的方法,取純化后的線粒體少許,在2.5% Gluta固定液于4℃固定2 h以上,用0.2 mol·L-1二甲砷酸鈉緩沖液洗兩次,每次15 min,再用1%的鋨酸(以0.2 mol·L-1二甲砷酸鈉配制)于4℃下固定2 h。之后進行脫水處理:4℃下分別在30%、50%和70%丙酮浸15 min,然后在室溫下分別在80%和90%丙酮中浸一次,再在95%丙酮中浸兩次,最后在100%丙酮浸兩次,以上每次均浸15 min。脫水后的樣品用Epon-812環氧樹脂包埋并進行切片,挑選出厚度在50—70 nm的切片置于銅網上,用醋酸雙氧鈾和檸檬酸鉛雙重染色,最后在日立JEM-1011型透射電子顯微鏡下觀察拍照。
1.4.2 線粒體功能活性的檢測 超氧化物歧化酶(SOD)活性的測定:參照SOD活性檢測試劑盒進行測定。
健那綠染色線粒體計數:參照高永超等[27]的方法,取線粒體懸浮液,滴一滴于載玻片,標記涂片,滴一滴0.02%健那綠染液,蓋上蓋片染色20 min。放于倒置顯微鏡下觀察并拍照。線粒體呈藍綠色顆粒狀。拍照通過ImageJ軟件進行計數。
1.4.3 能量水平和能荷值測定 參考 WU等[28]的方法測定三磷酸腺苷(adenosine triphosphate,ATP)、二磷酸腺苷(adenosine diphosphate,ADP)、一磷酸腺苷(adenosine monophosphate,AMP)含量。標準溶液的制備:將 ATP、ADP、AMP標準溶液分別稀釋為6.25、12.5、25、50和 100 mg·L-1的5個濃度梯度,得到的溶液用1 mol·L-1KOH調至pH 6.5—6.8并定容,經0.45 μm微孔濾膜過濾。樣品的制備:取0.5 g金針菇樣品,加入6 mL冰冷的0.6 mol·L-1高氯酸冰浴研磨并靜置10 min,3 000 r/min離心10 min,上清液與標液進行同樣的調 pH和過濾處理,濾液用于ATP、ADP和 AMP含量測定,以上操作均在 4℃條件下完成。制備好的標準溶液及樣品在下述色譜條件下上機進樣,并記錄各標準品及樣品峰面積。以不同濃度各標樣峰面積y為縱坐標,以標樣質量濃度x為橫坐標,繪制標準曲線,回歸方程及相關系數見表1。色譜條件:色譜柱(Eclipse plus C-18,5 μm×150 mm×4.6 mm);流動相:A:0.06 mol·L-1的K2HPO4+0.04 mol·L-1的 KH2PO4(pH 7.0),B:100%甲醇;柱溫:30℃;流速:1 mL·min-1;檢測波長:254 nm;洗脫條件:ATP:50%A,50%B;ADP、AMP:95%A,5%B。洗脫時間:4 min。
能量負荷值測定:能量負荷值=(ATP+1/2ADP)/(ATP+ADP+AMP)。
1.4.4 線粒體關鍵復合體酶活性的測定 線粒體復合體I、Ⅲ、Ⅳ活性的測定:利用1.3.1中對比確定的最佳線粒體提取方法對金針菇線粒體進行提取,并參照北京索萊寶科技有限公司線粒體復合體 I、Ⅲ、Ⅳ活性檢測試劑盒中說明書提供的方法進行活性測定。
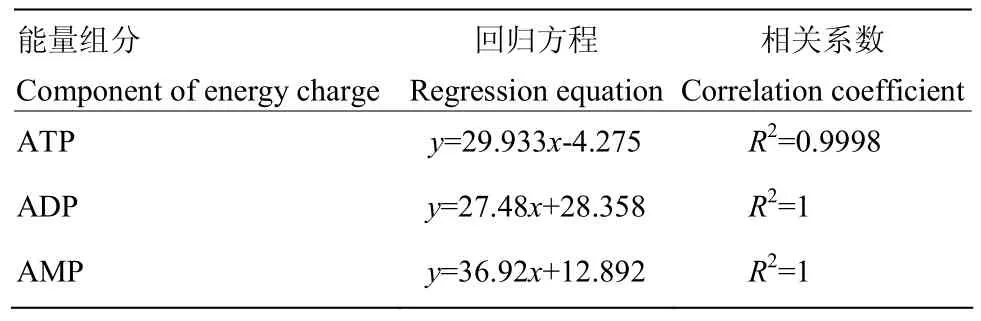
表1 ATP、ADP 和 AMP 的標準曲線回歸方程和相關系數Table 1 Standard curve line regression equations and correlation coefficients of ATP, ADP and AMP
1.5 數據處理與分析
使用Excel進行數據處理,并用Origin 8.5繪圖,SPSS Statistics17.0進行顯著性分析,顯著性水平設置為P<0.05。
2 結果
2.1 不同提取方法對金針菇線粒體結構完整性的影響
2.1.1 乙醇脫氫酶活性 乙醇脫氫酶(alcohol dehydrogenase,ADH)和細胞色素C氧化酶(cytochrome c oxidase,COX)是分別穩定存在于胞液及線粒體內的標志性蛋白,當線粒體膜發生破碎時,二者會分別向線粒體及胞液中流動,因此,通過測定它們在線粒體及胞液中的活性,可以來反映線粒體膜的破損情況[20]。在不連續密度梯度離心法(DDGCM)中,使用的洗滌液濃度高低會影響線粒體的產量,因此本研究使用 3種濃度的洗滌液(DDGCM-1、DDGCM-200、DDGCM-400)進行篩選以確定最佳提取參數。

圖1 金針菇子實體所提線粒體(A)及胞液(B)中乙醇脫氫酶活性Fig.1 Activity of alcohol dehydrogenase in mitochondria (A) and cytosol (B) of F. filiformis fruiting body
由圖1可見,在子實體組,DDGCM和酵母試劑盒法(YMEKM)所提取的線粒體中ADH活性均比改進普利萊法(IPKM)的活性高。其中,雖然DDGCM未稀釋洗滌液組所提取的線粒體中 ADH活性最低,但仍然高于IPKM。另外,IPKM所得胞液中的ADH活性最高,與各組都存在顯著性差異。由圖2可見,在菌絲體組,雖然各方法得到的胞液中 ADH活性結果與子實體組相同,但是線粒體ADH活性中YMEKM所得與DDGCM-200相比無顯著性差異,而IPKM所得活性仍然最低。因此,相比之下,IPKM 提取得到的線粒體受胞質污染最輕。
2.1.2 細胞色素C氧化酶活性 由圖3可見,在子實體組中,DDGCM提取的線粒體中 COX活性隨著洗滌液濃度的降低而降低,盡管未稀釋洗滌液組在3個梯度中COX活性最高,并且顯著高于YMEKM,但顯著低于IPKM所提取線粒體中的COX活性。另外,胞液中的COX活性結果顯示YMEKM最高,DDGCM其次,IPKM 的最低,并且三者均有顯著性差異。綜上所述,IPKM 所提取的線粒體受胞質污染最輕,與2.1.1得到的結果一致。此外,由圖4可見,在菌絲體組中,所得結果與子實體趨勢一致。
2.1.3 線粒體呼吸速率 細胞色素C氧化酶損失會直接導致線粒體呼吸能力下降,呼吸速率降低[29]。因此,為了進一步驗證分離線粒體的呼吸能力,本研究使用氧電極對線粒體呼吸速率進一步評估。通過測定線粒體提取前后的呼吸速率,對比各方法在提取后的呼吸速率差異來評估它們對于線粒體膜的損傷程度。由圖 5可知,無論是子實體還是菌絲體中,IPKM所提的線粒體與未提取前差距最小,且顯著高于其他兩種方法所提線粒體的呼吸速率,證明該方法提取得到的線粒體能更好地維持其功能活性。雖然使用多個呼吸鏈底物配制線粒體反應液,但提取的線粒體呼吸速率與細胞內呼吸相比較低,這可能與反應液模擬細胞環境存在誤差有關。隨著菌絲生長,呼吸速率呈上升趨勢。在不同生長時期,菌絲的呼吸速率增速不同[30],因此與子實體相比,菌絲體的呼吸速率相對較慢。

圖2 金針菇菌絲體所提線粒體(A)及胞液(B)中乙醇脫氫酶活性Fig.2 Activity of alcohol dehydrogenase in mitochondria (A) and cytosol (B) of F. filiformis mycelium
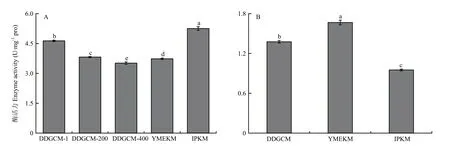
圖3 金針菇子實體所提線粒體(A)及胞液(B)中細胞色素C氧化酶活性Fig.3 Activity of Cytochrome C oxidase in mitochondria (A) and cytosol (B) of F. filiformis fruiting body
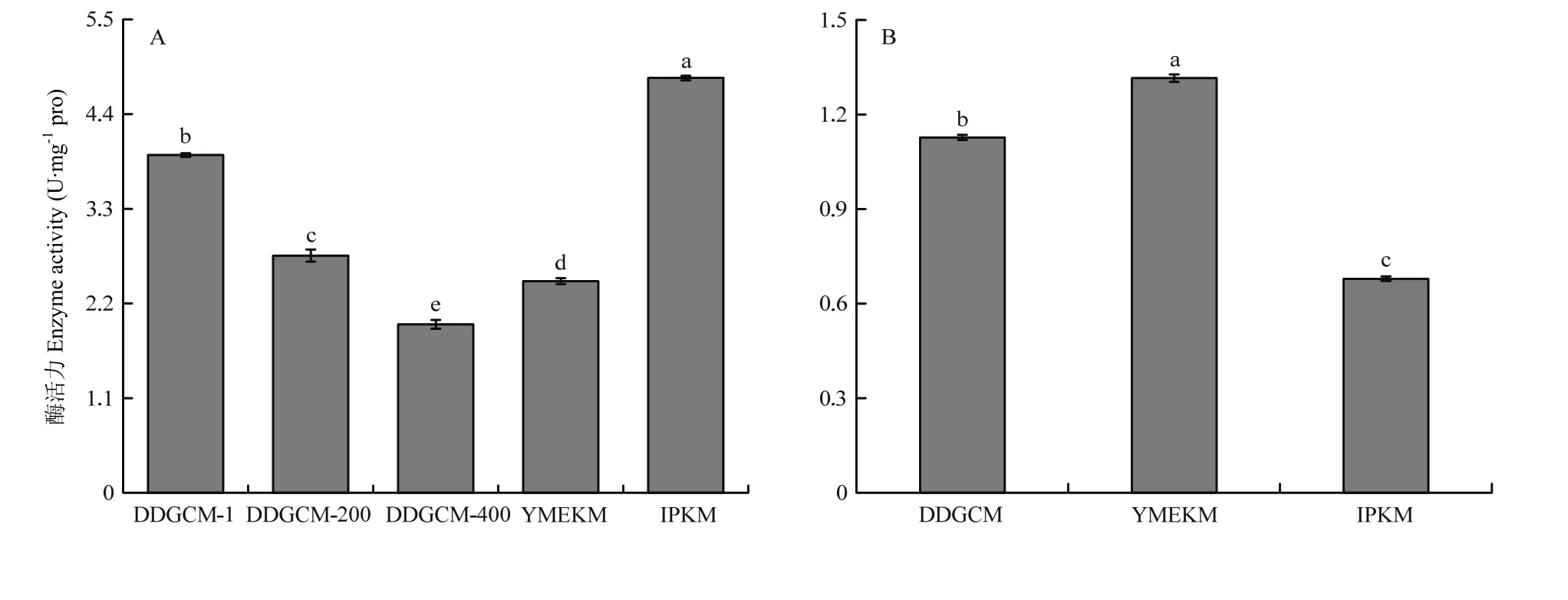
圖4 金針菇菌絲體所提線粒體(A)及胞液(B)中細胞色素C氧化酶活性Fig.4 Activity of Cytochrome c oxidase in mitochondria (A) and cytosol (B) of F. filiformis mycelium
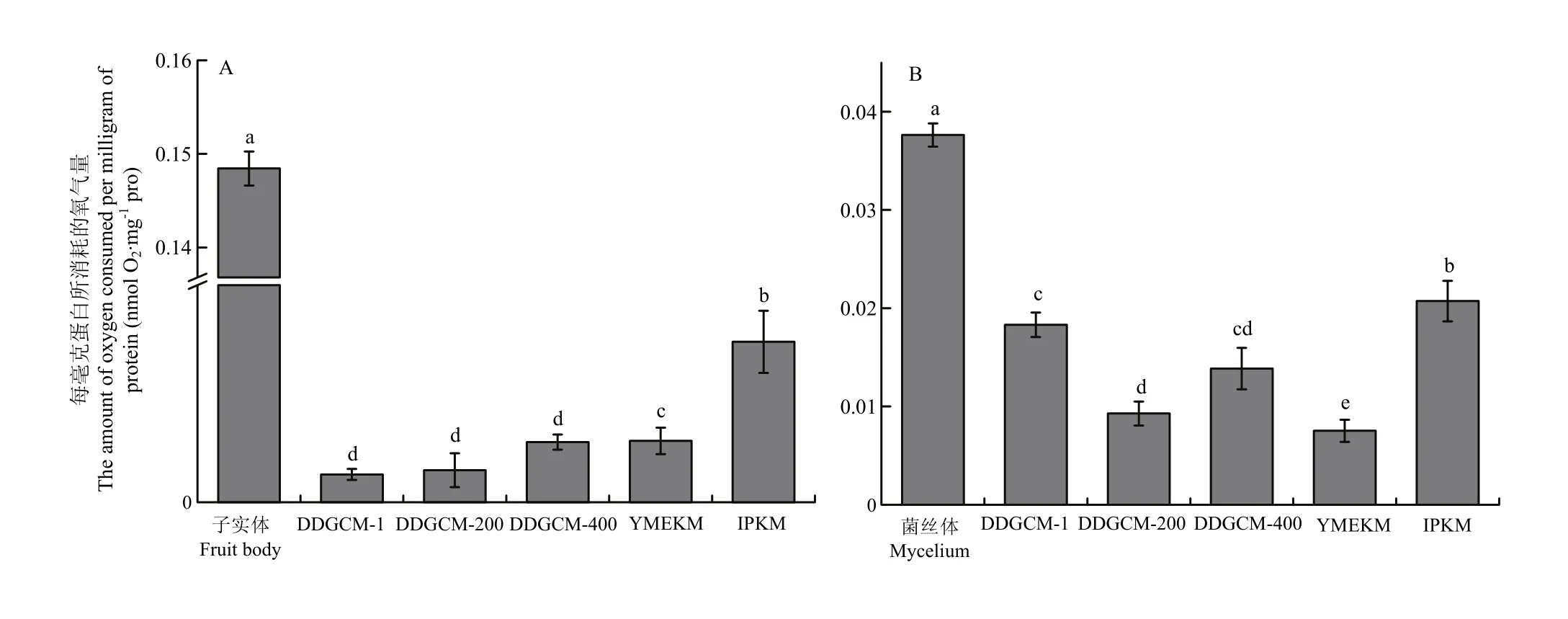
圖5 金針菇子實體(A)、菌絲體(B)線粒體的呼吸氧消耗Fig.5 The number of active mitochondria extracted from the fruit body (A) and mycelium (B) of F. filiformis
2.1.4 透射電鏡觀察線粒體 在圖 6的顯微鏡照片中,所有樣品組中都可見典型的橢圓形線粒體。其中,DDGCM隨著洗滌液的逐級稀釋,具有完整結構的線粒體也越來越少,并且腫脹現象也越來越嚴重。YMEKM制備的線粒體腫脹并表現出線粒體膜破碎嚴重,線粒體膜碎片在該組圖片中清晰可見。而 IPKM提取得到的線粒體雖然也存在少量的碎片,但得到的形態結構完整的線粒體數量最多,與其他組樣品形成明顯對比。
2.2 不同提取方法對金針菇線粒體功能活性的影響
2.2.1 超氧化物歧化酶活性 超氧化物歧化酶(superoxide dismutase,SOD)是細胞中重要的抗氧化酶,分為3大類,分別為Cu/Zn-SOD、Mn-SOD、Fe-SOD。其中存在于線粒體內膜基質中的是 Mn-SOD,與線粒體的呼吸代謝有關[31],并維持線粒體膜的完整性和正常功能[32]。在不連續密度梯度離心法中,隨著洗滌液濃度的降低,所提線粒體中SOD活性逐漸降低,因此,選取未稀釋洗滌液組所提取的線粒體中SOD活性與其他兩個方法進行對比。由圖7可見,IPKM所提取的線粒體中SOD活性最高,酵母試劑盒法提取的線粒體中SOD活性最低,子實體組與菌絲體的趨勢一致。
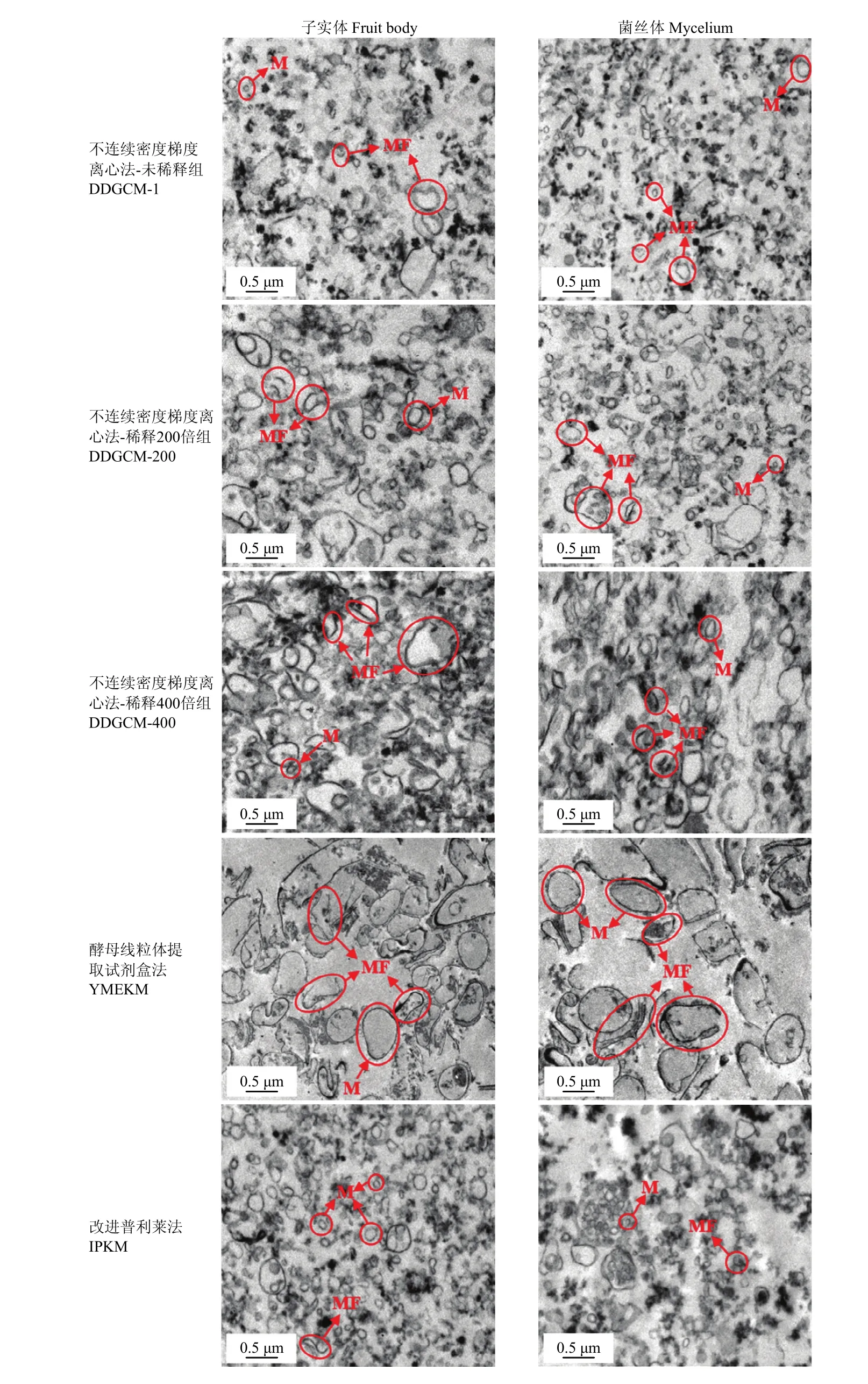
圖6 透射電鏡觀察線粒體微觀結構Fig.6 TEM observation of mitochondrial microstructure
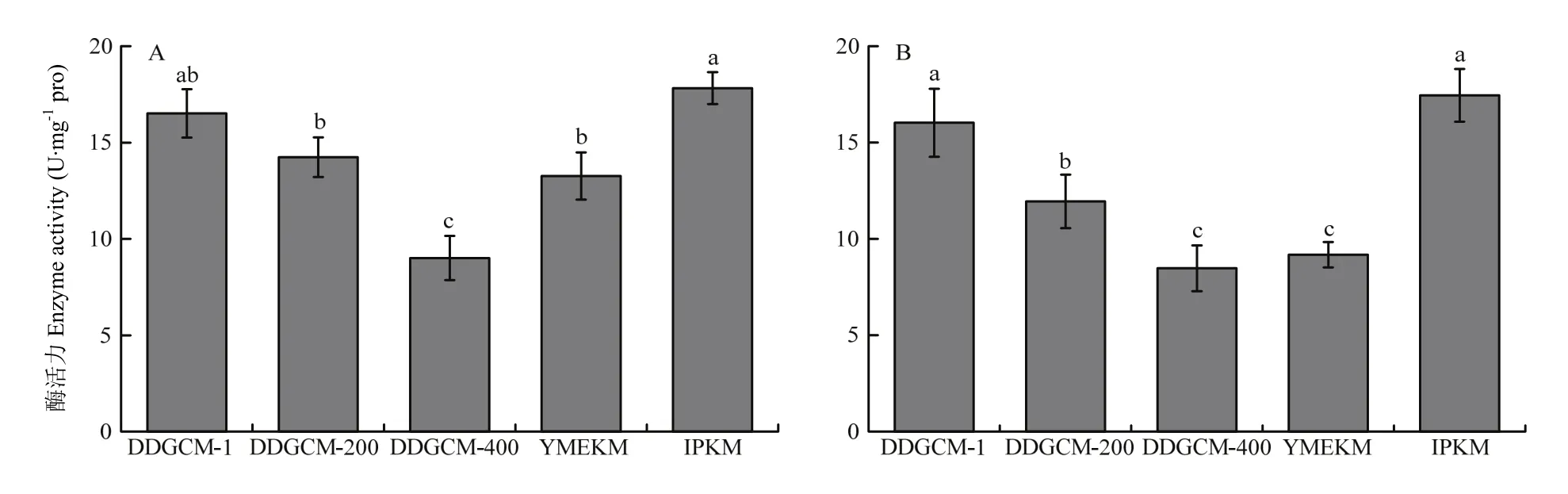
圖7 金針菇子實體(A)及菌絲體(B)所提線粒體中超氧化物歧化酶活性Fig.7 Activity of Superoxide dismutase in mitochondria of F. filiformis fruiting body (A) and mycelium (B)

圖8 健那綠染色活性線粒體Fig.8 Janus green B staining active mitochondria
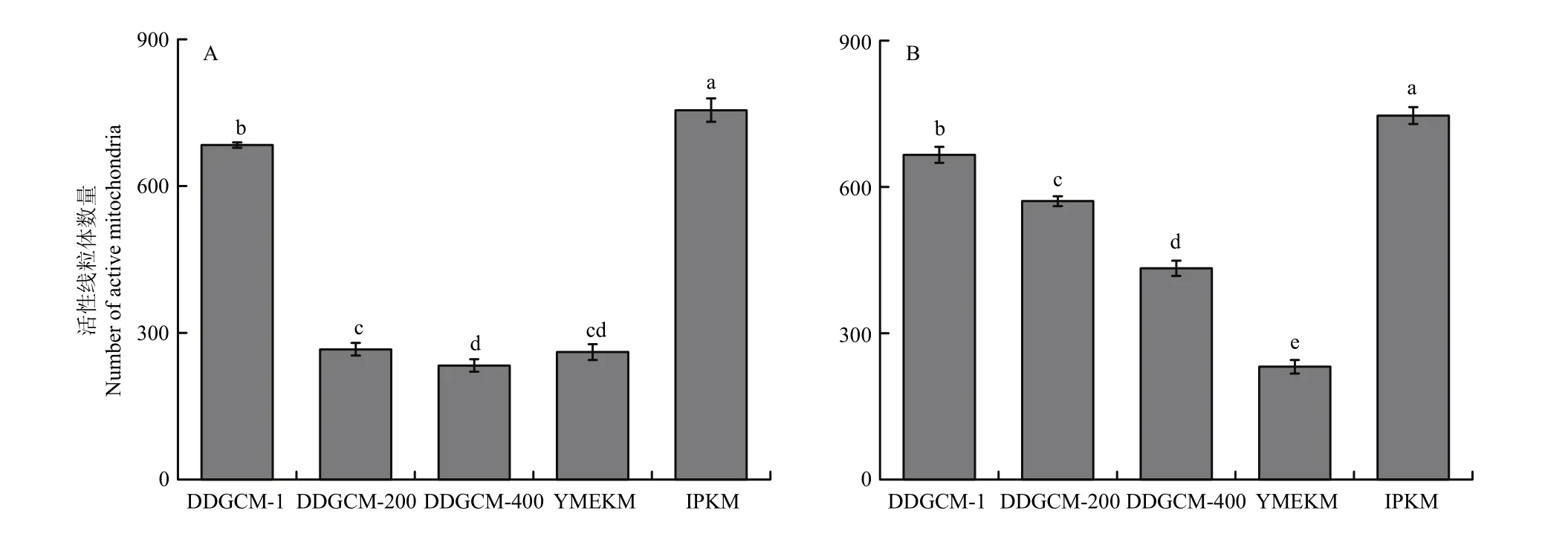
圖9 各提取方法的金針菇子實體(A)及菌絲體(B)中活性線粒體數量Fig.9 The number of active mitochondria extracted from the fruit body (A) and mycelium (B) of F. filiformis by each extraction method
2.2.2 健那綠染色顯微鏡拍照及計數 由圖8、9可見,在子實體中,DDGCM隨著洗滌液稀釋倍數的升高,所得活性線粒體數量逐漸減少。未稀釋洗滌液組的活性線粒體數量在三者中最多,而IPKM所得活性線粒體數量整體最多,顯著高于其他兩組。在菌絲體中,仍然是IPKM所得活性線粒體數量最多,與其他組均具有顯著差異;而YMEKM得到的活性線粒體數量最低。
2.3 不同包裝金針菇冷藏期間外觀評估
由圖10可見,與納米包裝相比,普通PE包裝內表面的水分凝結現象更明顯,分布更均勻。此外,在冷藏21 d后,兩個包裝處理組都呈現出一定程度的褐變及子實體開傘現象,而普通 PE包裝處理組的金針菇較納米包裝組更嚴重。
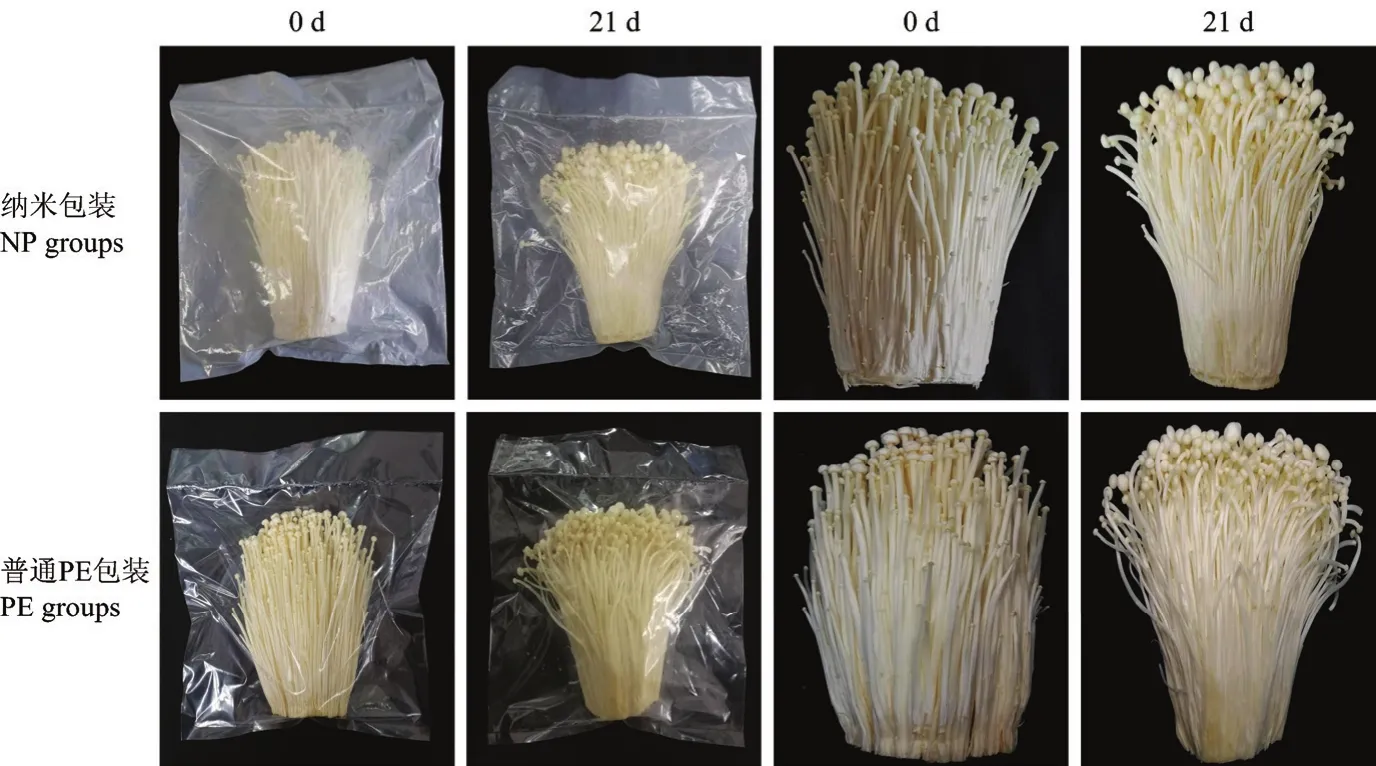
圖10 不同包裝對金針菇冷藏期間外觀質量的影響Fig.10 Effects of different packaging materials on appearance quality of F.filiformis during refrigeration
2.4 不同包裝金針菇冷藏期間能量水平和能荷值
由圖 11可見,在貯藏期間,納米包裝金針菇的ATP含量始終顯著高于其他處理組,AMP含量始終顯著低于其他處理組,表明納米包裝能有效控制ATP含量的降低,同時還抑制了 AMP含量的增加。在整個貯藏過程中,隨著ATP、ADP和AMP含量的變化,金針菇的能荷值也發生變化,而且在貯藏期間納米包裝組的能荷值始終顯著高于普通PE包裝組(P<0.05)。
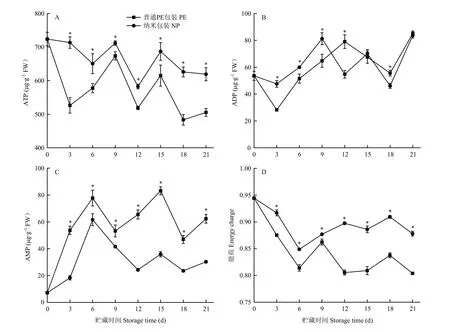
圖11 不同包裝對金針菇冷藏期間ATP含量(A)、ADP含量(B)、AMP含量(C)、能荷(D)的影響Fig.11 Effect of different packaging materials on changes of ATP content (A), ADP content (B), AMP content (C) and energy charges (D) of F.filiformis during refrigeration
2.5 不同包裝金針菇冷藏期間線粒體關鍵復合體酶活性
由圖 12可見,在貯藏過程中,線粒體復合體 I和復合體Ⅲ活性均呈現先升高再降低的趨勢且兩個包裝組變化趨勢一致。對于線粒體復合體Ⅳ活性,兩個包裝組都在0—6 d呈現先下降的趨勢,普通PE包裝組樣品的復合體活性在貯藏第9天時達到峰值,隨后逐漸下降,而納米包裝組的線粒體復合體Ⅳ活性則在貯藏第15天時才達到峰值然后下降。在整個貯藏期間,納米包裝組的3個線粒體復合體酶活性均顯著高于普通PE包裝組,說明納米包裝可以在貯藏期間使線粒體復合體酶維持高活性,從而維持金針菇的能量代謝水平。
3 討論
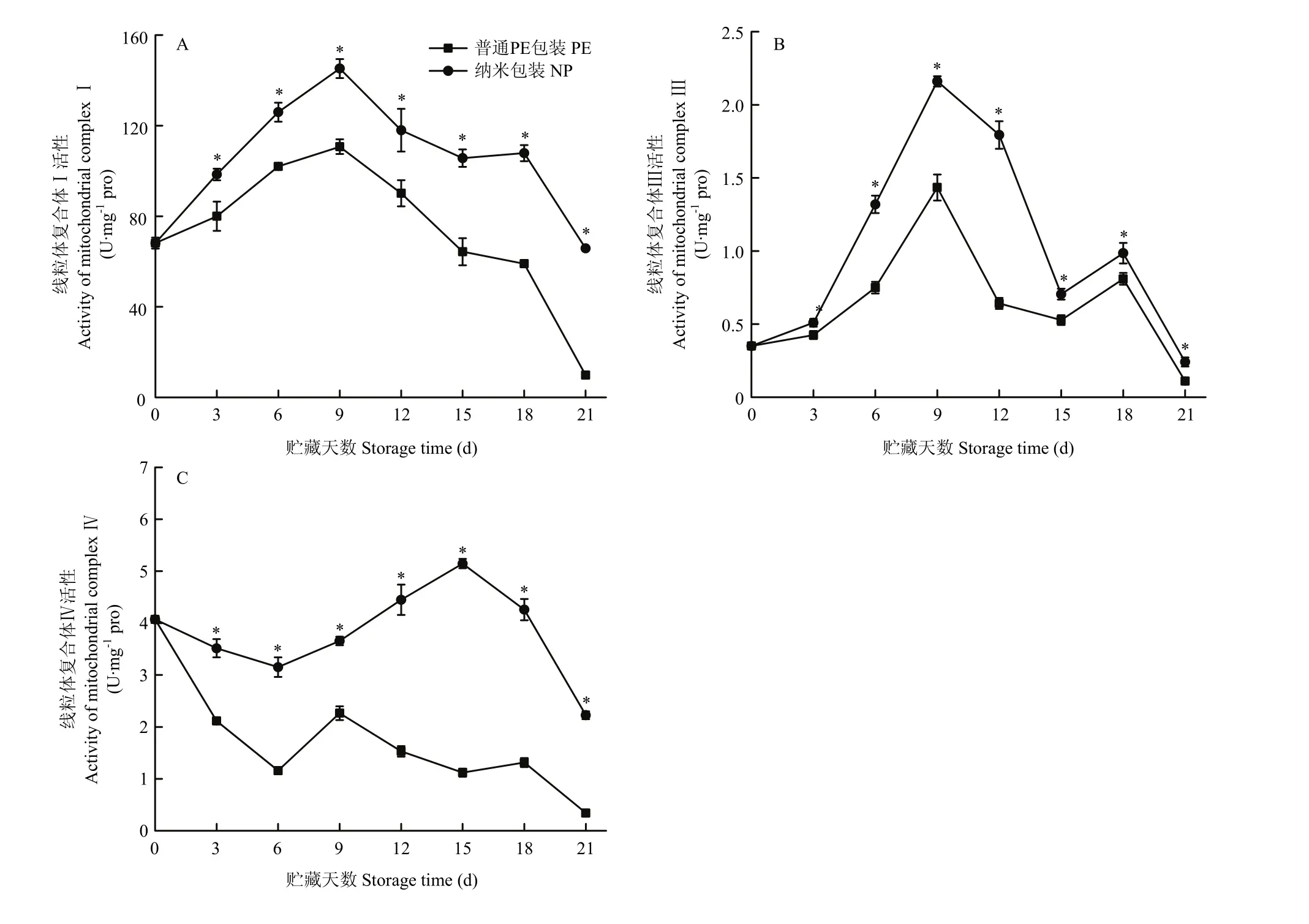
圖12 不同包裝對金針菇冷藏期間線粒體復合體I(A)、復合體Ⅲ(B)和復合體Ⅳ(C)活性的影響Fig.12 Effects of different packaging materials on changes of mitochondrial complex I (A), complex Ⅲ (B) and complex Ⅳ (C) of F.filiformis during refrigeration
線粒體是能量代謝的主要場所,它為細胞活動提供了80%的能量,其中三羧酸循環發生于線粒體基質中,而電子傳遞鏈和氧化磷酸化的發生位點在線粒體內膜上[21]。提取高純度的線粒體,是獲得高質量線粒體蛋白的重要前提,有助于分子生物學以及蛋白質組學的進一步研究,為后續線粒體功能的研究及其蛋白質組的分析奠定了良好的基礎[33]。然而在不同物種的細胞中,細胞結構有所差別,因此,針對不同物種細胞有不同的分離線粒體的方法[34-36]。本研究結果表明改進普利萊法所得的線粒體受胞質污染程度最低,更好地保持了線粒體的膜完整和活性。酵母線粒體提取試劑盒長時間的提取分離操作會對細胞壁、線粒體結構和活性造成損傷,而且過高或過低的緩沖液 pH會引起線粒體腫脹和膜破碎[37],進而降低了線粒體活性。不連續蔗糖密度梯度離心法屬于傳統方法,盡管線粒體在分離過程中會從破碎的細胞中迅速分離出來,但在將線粒體分散到懸浮介質中之前,線粒體會與液泡的內容物有短暫的接觸,其中一些水解酶將附著在線粒體外膜上。因此,線粒體外膜有被這些污染性的水解酶裂解的風險[38]。與此同時,多次的高速離心同樣會使部分線粒體膜破損,并且高濃度蔗糖引起的滲透損傷也是造成線粒體內膜破損的重要原因[39]。相比之下,改進普利萊法能通過有效的破壁方法和較少的離心步驟得到結構較完整且活性較好的線粒體,而整個分離過程的關鍵點是對金針菇細胞壁的破碎及細胞膜的裂解。真菌的細胞壁主要由葡聚糖、幾丁質和蛋白質組成[40],其中幾丁質與植物中的纖維素類似,形成堅硬的微纖維,強化細胞壁的強度,并且與α-1,3-葡聚糖一起排列[41-42]。因此,破壞幾丁質結構是破壁處理的關鍵。而蝸牛酶內含有多種酶,能夠高效降解幾丁質[43]。對于改進普利萊法,分離前用巰基化合物增強胞內蛋白穩定性的同時[44],在等滲條件下使用蝸牛酶進行酶解破壁,為得到大量完整的線粒體奠定了基礎。
有研究表明金針菇表面結構薄,不能有效阻止其表面水分的迅速散失[45]。而采后的金針菇子實體或包裝袋內表面會因貯藏環境溫差出現水汽凝結現象,該現象有利于微生物的繁殖、侵染和傳播[46]。納米包裝材料可以有效控制金針菇采后蒸騰作用,減少包裝內部水汽凝結,這與前期研究結果一致[13]。筆者實驗室前期對冷藏期間不同包裝金針菇的孢子數量檢測發現,第15天無包裝組和普通PE包裝組孢子數量分別達到4.84×1011和4.09×1011/g cap,而納米包裝組只有3.44×1011/g cap,顯著低于其他兩組(P<0.05),表明納米包裝材料能有效調節包裝袋內濕度和氣體成分,減輕水汽凝結,控制微生物生長,減緩金針菇腐爛酸敗。冷藏期間納米包裝材料可以抑制金針菇的菌柄伸長、開傘、褐變,該現象是采后金針菇后熟作用的重要表現[47]。在冷藏期間,金針菇后熟作用主要由包裝內的溫度、氣體成分以及乙烯含量等因素所影響。有研究表明,金針菇在采后菌絲能產生高濃度的乙烯,促進其呼吸作用,從而加快其衰老速度[48]。前期對納米包裝在冷藏期間的氣體成分進行了分析,結果未檢測出乙烯,表明納米包裝材料中聚合的納米顆粒可能會吸收金針菇釋放的乙烯[22]。此外,納米包裝可以通過抑制金針菇的呼吸速率,減少活性氧積累,從而保護線粒體結構,維持金針菇的能量代謝水平[13]。
已有研究表明果蔬的生理失調和褐變與細胞能量的變化有關[49]。線粒體是細胞中進行生物氧化和產生ATP的主要場所。因此,果蔬組織及線粒體中主要能量物質的變化已成為細胞能量代謝研究的主要觀察指標[50]。ATP是至關重要的能源,保障果蔬組織正常的生理代謝功能[51]。同時,能荷值的高低決定了線粒體磷酸化活性的高低。筆者實驗室前期研究中發現納米包裝材料可以有效防止線粒體結構損傷[3],而線粒體結構受損時會導致磷酸化活性降低,進而誘導ATP和能荷低水平表達。而除了自然衰老劣變,線粒體的提取過程也會導致細胞裂解和線粒體結構損傷,這一點在線粒體提取方法篩選試驗中已得到驗證。這也能解釋前人在貯藏試驗中使用差速離心法提取線粒體時,測得的線粒體主要復合體活性指標并沒有明顯波動的原因[3]。電子傳遞鏈主要由線粒體復合體I(NADH脫氫酶)、線粒體復合體Ⅲ(細胞色素b/c1氧化還原酶)和線粒體復合體Ⅳ(細胞色素C氧化酶)組成,為細胞提供ATP發揮關鍵作用[14]。鑲嵌于線粒體內膜上的線粒體復合體,它們的活性高低是表征線粒體功能變化的重要指標,同時也是三羧酸循環和氧化磷酸化等能量代謝過程中的關鍵酶,可以反映ATP合成情況和線粒體內膜損傷程度[52]。本研究結果表明,納米包裝組金針菇具有更高水平的ATP和能荷值,并且納米包裝延遲了能荷的下降;而ATP代謝系統各物質含量在金針菇冷藏期間均呈現波動變化,這是由于食用菌屬于呼吸躍變型果蔬[53]。對于呼吸躍變型果蔬,研究人員常常把其呼吸躍變的高峰期作為水果由成熟走向衰老的轉折期。有研究表明,能荷的降低也是衰老的體現[54]。因此,納米包裝材料在冷藏期間通過抑制ATP水平的下降和維持高水平的能荷值來延緩金針菇的衰老。與此同時,ATP供應不足可直接引起線粒體腫脹和嵴斷裂,并進一步抑制線粒體的氧化磷酸化過程,這一現象在線粒體主要復合體活性的測定中得到了證實。在線粒體主要復合體中,線粒體復合體Ⅳ是線粒體呼吸和三羧酸循環(tricarboxylic acid cycle,TCA)中的關鍵酶[55],為氧化磷酸化提供能量,其活性高低直接影響到線粒體呼吸鏈中的電子傳遞。低活性的線粒體復合體Ⅳ會使ATP生成減少、能荷降低,從而導致細胞所需能量供應不足。此外,呼吸鏈障礙在導致氧化不完全的同時也會造成活性氧生成增多,以上均會導致細胞死亡[56]。在冷藏過程中,納米包裝顯著延緩了該酶活性的下降趨勢,與能荷值變化表現一致,進一步證明納米包裝材料延緩了金針菇氧化磷酸化速率的下降,保障了細胞ATP供應,維持基本能量代謝需要。對于不同包裝金針菇線粒體差異蛋白表達譜以及能量代謝調控路徑中關鍵性蛋白還不明確,后續還需利用線粒體蛋白組,從分子水平上進一步闡明納米包裝對金針菇能量和呼吸代謝的作用途徑與機制。
4 結論
通過分析線粒體的結構完整性和功能活性,確定改進普利萊法最適合金針菇線粒體的提取。在此基礎上,利用改進普利萊法對不同包裝的金針菇線粒體進行提取,證明納米包裝通過維持高水平的ATP含量與線粒體復合體I、III活性,延緩能荷與線粒體復合體IV的下降,從而更好地保持了金針菇的能量狀態并延緩了衰老。
