CYP79B2/B3基因在吲哚族硫苷與IAA代謝中的功能研究進展
2020-08-28 11:34:30孫夏莉臧運祥徐義康許雙雙
江蘇農業科學 2020年14期
孫夏莉 臧運祥 徐義康 許雙雙
摘要:吲哚族硫苷與吲哚-3-乙酸(indole-3-acetic acid,簡稱IAA)在十字花科植物生長發育和防御過程中發揮了重要作用。CYP79B2/B3基因催化生成的吲哚乙醛肟(IAOx)是這2條代謝途徑的共同前體物,因此,闡明CYP79B2/B3基因在吲哚族硫苷與IAA代謝平衡中的作用,對于十字花科植物生長發育調控及病蟲害防御研究具有重要意義。本文對吲哚族硫苷合成途徑、IAA合成途徑及CYP79B2/B3基因在2條代謝途徑中的功能研究進展進行了綜述。
關鍵詞: CYP79B2/B3;吲哚族硫苷;IAA;代謝途徑;功能分化
中圖分類號:Q945.18;S184?? 文獻標志碼: A
文章編號:1002-1302(2020)14-0050-05
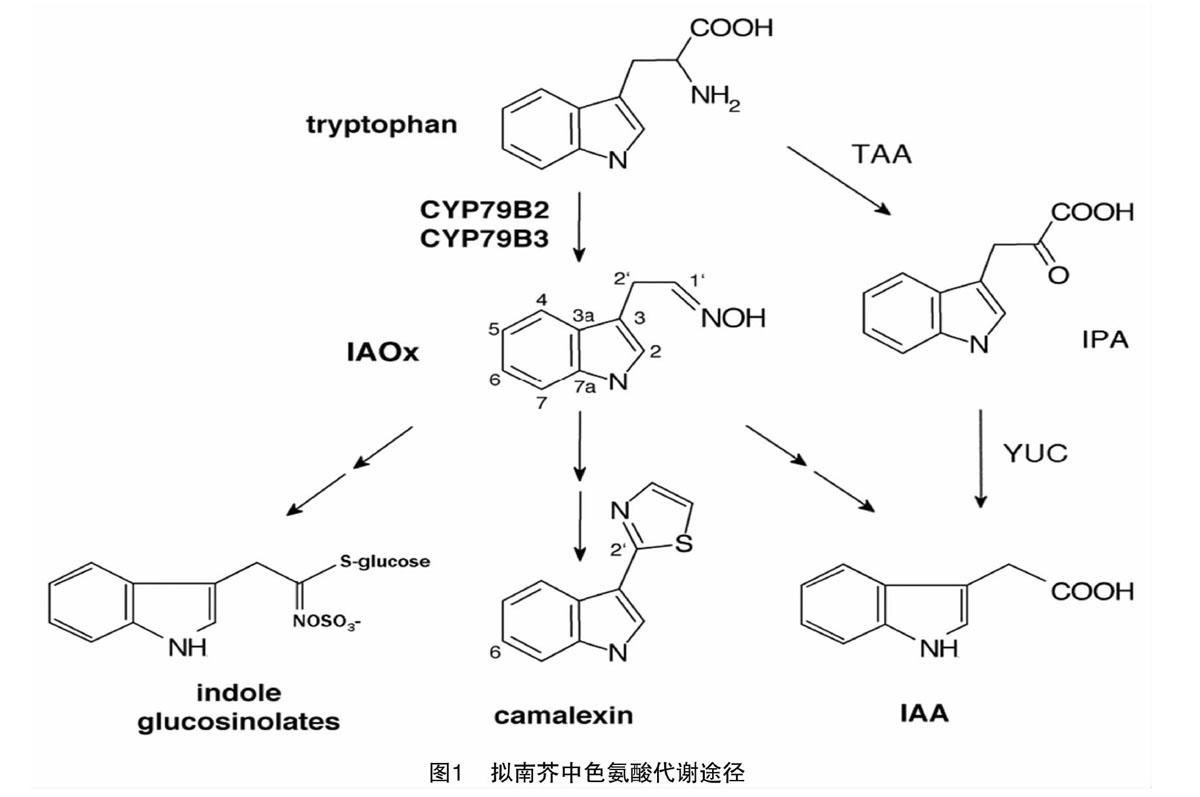
十字花科植物富含一類具有抗病蟲功能的重要次生代謝物質——硫代葡萄糖苷(簡稱硫苷),其水解產物具有較強的抗病蟲能力[1]。硫苷由硫代葡萄糖基、磺酸肟和源于不同氨基酸的R側鏈3個部分組成。根據側鏈基團的種類和性質不同,可將其分為脂肪族、芳香族和吲哚族硫苷3類[2]。吲哚族硫苷與吲哚-3-乙酸(indole-3-acetic acid,簡稱IAA)的合成具有共同的前體物——吲哚乙醛肟(IAOx),由CYP79B2/B3催化形成(圖1)[3]。模式植物擬南芥中有CYP79B2、CYP79B3等2個基因,而白菜等十字花科蔬菜中含有多個拷貝的CYP79B2/B3基因。因此,闡明CYP79B2/B3基因參與吲哚族硫苷與IAA代謝平衡的分子機制,可為十字花科植物生長發育調控及其對害蟲、病原菌的防御研究提供理論基礎。本文擬圍繞吲哚族硫苷與IAA代謝,對CYP79B2/B3基因在2條代謝途徑中的功能研究進展進行簡要綜述,以便為他人開展相關研究提供參考。
1 CYP79B2/B3基因在吲哚族硫苷代謝途徑中的功能
1.1 硫苷的作用
硫代葡萄糖苷是源于氨基酸的含氮硫陰離子的植物次級代謝產物,目前自然界中已發現200多種硫代葡萄糖苷,其中模式植物擬南芥中約有40種硫代葡萄糖苷[4-5]。硫苷水解產物,如異硫氰酸酯、腈類和硫代氰酸酯等對草食動物和多種微生物具有抵御作用,近年來又因個別組分的強效抗癌、抗氧化作用而引起植物學、醫學、生物學、食品科學等眾多學科研究者的關注。前人的研究結果表明,吲哚族硫苷對草食性動物具有威懾作用,并且有助于預防害蟲和影響昆蟲產卵,是構成植物防御體系不可或缺的一部分[6-8]。此外,吲哚族硫苷水解產物吲哚-3-甲醇可以通過控制Ⅰ相酶與Ⅱ相酶活性來預防前列腺癌、肺癌、膀胱癌、食道癌等,能通過多條途徑預防腫瘤發生,促進機體抗氧化物的生成[9-11]。
1.2 吲哚族硫苷的合成途徑
吲哚族硫苷的合成底物源于色氨酸。擬南芥中含有4種吲哚族硫苷,分別是吲哚-3-甲基硫苷、1-甲氧基-吲哚-3-甲基硫苷、4-羥基-吲哚-3-甲基硫苷和4-甲氧基-吲哚-3-甲基硫苷[12-14]。
目前擬南芥中吲哚族硫苷的合成途徑已經較為清楚。先由分支酸鹽經鄰氨基苯甲酸酶(AS)催化反應形成鄰氨基苯甲酸,經過多步反應后形成吲哚,再經色氨酸合成酶B(TSB)催化形成色氨酸;然后由細胞色素P450酶CYP79B2和CYP79B3催化生成吲哚-3-乙醛肟(IAOx)[15];再然后,在單加氧酶CYP83B1的催化作用下形成不穩定的吲哚-3-甲基酸式硝基化合物,進入吲哚族硫苷合成途徑;隨后,通過非酶促反應或谷胱甘肽硫轉移酶(GSTF9和GSTF10)的作用獲得半胱氨酸或由谷胱甘肽(GSH)提供的硫,生成S-吲哚-3-甲基-硫代氧肟,經γ-谷氨酰多肽合成酶GGP1和C-S裂解酶SUR1(C-S lyase)催化生成硫代肟基酸[16-17];之后,在葡萄糖基轉移酶UGT74B1的催化作用下發生糖基化而形成脫硫硫苷[18];最后,經硫酸根轉移酶AtST5a酸化生成完整的吲哚族硫苷,其核心結構再經過羥基化、甲基化、葡萄糖基化等次級修飾而形成不同的吲哚族硫苷[19]。
硫苷的合成涉及許多因素,轉錄調控網絡復雜。研究發現,MYB類轉錄因子(MYB34、MYB51和MYB122)在參與硫苷生物合成的過程中能特異地調控吲哚族硫苷的合成。MYB34被認為是色胺代謝的正向調節因子,在過量表達突變體中發現CYP79B2、CYP79B3和CYP83B1的表達量均顯著增加,吲哚族硫苷的含量增加了約10倍[20-21]。當合成基因突變導致吲哚族硫苷缺失時,MYB34的轉錄水平大幅提升,說明MYB34在植物吲哚族硫苷含量缺失時被激活。機械損傷可以誘導MYB51過量表達,從而調控CYP79B2、CYP79B3、CYP83B1、UTG74B1、At ST5a等表達,引起植物體內吲哚族硫苷大量積累[22]。
MYB34、MYB51和MYB122能促進植物根、芽中吲哚族硫苷的生物合成,MYB122在此過程中可能協助MYB34起輔助作用。在myb34、myb51、myb122三重突變體中沒有檢測到吲哚族硫苷,說明這3個MYB因子對于吲哚族硫苷的合成是必不可少的[23]。有研究發現,IQD1和OBP2可能是間接調控因子,IQD1對吲哚族硫苷、脂肪族硫苷都具有正向調節作用,OBP2則是通過提高CYP83B1的表達量來增加吲哚族硫苷含量[24-26]。
1.3 CYP79B2/B3基因在代謝途徑中的功能
Hull等將擬南芥CYP79B2和CYP79B3基因轉入酵母后發現,這2個基因均能將色氨酸轉化為吲哚-3-乙醛肟,并且CYP79B2基因的過表達使得擬南芥產生抵抗毒性的色氨酸類似物[27]。Zhao等通過在擬南芥中過表達CYP79B2和CYP79B3基因發現,轉基因植株中吲哚族硫苷和生長素含量均顯著提高,而擬南芥CYP79B2/B3雙突變體中則完全缺失吲哚族硫苷,存在較低含量的生長素,說明這2個酶都能催化吲哚-3-乙醛肟合成生長素和吲哚族硫苷[28]。CYP79B2在氨基酸水平上與CYP79B3有85%的同源性。一般來說,CYP79B2在氨基酸水平上與其他CYP[STBX]79[STBZ]顯示出44%~67%的相似性,這可能解釋了兩者存在功能上的冗余[29]。研究發現,參與色氨酸代謝的CYP79B2、CYP79B3基因在 1 μmol/L 2,4-D上生長的擬南芥中被誘導表達,導致吲哚硫苷積累量增加。在生長素水平高的sur1中,CYP[STBX]79B2、CYP79B3[STBZ]被誘導,從而增加了IAOx產量,并增強了超根性表型,這說明CYP79B2、CYPB3酶通過催化生成IAOx進入吲哚族硫苷和IAA代謝途徑[30]。
Mikkelsen等指出,CYP79B2不僅在植物葉片、莖、花和根部可誘導,而且在植物根中的表達量最高;另外關于CYP79B2的GUS報告基因表達研究發現,GUS報告基因在植物幼苗期根系中有較高表達量,在子葉中部分表達,而在老葉中的表達很弱,CYP79B2的表達模式與擬南芥中吲哚族硫苷的分布非常對應[29]。擬南芥cyp79b2突變體中吲哚族硫苷含量比cyp79b3突變體高[28]。以上研究結果說明,與CYP79B2相比,在吲哚族硫苷合成過程中,CYP79B3可能發揮更重要的作用。
2 CYP79B2/B3[STHZ]基因在IAA代謝途徑中的功能
2.1 IAA合成途徑
吲哚-3-乙酸是植物體內最主要的天然活性生長素,調節、參與植物的生長發育和多種生理過程,特別是植物細胞生長和分裂、植株向地性和向光性的形成、主側根及下胚軸生長、維管束發育和花器官形成等形態構建過程[31]。按照合成底物的不同,IAA合成途徑被分為依賴于色氨酸的途徑和不依賴于色氨酸的途徑[32-33]。在十字花科植物中,IAA合成主要依賴色氨酸途徑,依據中間產物又可分為吲哚丙酮酸(indole-3-pyruvic acid,簡稱IPA)途徑、吲哚乙醛肟(indole-3-acetaldoxime,簡稱IAOx)途徑、色胺(tryptamine)途徑和吲哚乙酰胺(indole-3-acetamide,簡稱IAM)途徑4條支路。遺傳學和酶學研究發現,TAA和YUC基因在合成植物IAA的過程中是必需的,TAA[STBX]1[STBZ]是吲哚丙酮酸合成途徑中的關鍵基因,其編碼蛋白催化色氨酸轉化為吲哚-3-丙酮酸,最后生成IAA,YUCCA是色胺途徑中的限速酶,可經羥基化形成N-羥基色胺,脫羧后形成IAOx,最后由多步反應轉化為生長素。IAA的合成途徑復雜,且途徑之間存在交叉。IPA途徑被證實是擬南芥中主要的IAA生物合成途徑[34]。吲哚乙醛肟途徑被研究人員普遍認為不是植物體內主要的IAA合成途徑,卻是十字花科植物防御體系中的一個重要調控環節,與吲哚族硫苷的合成存在相關性。
2.2 CYP79B2/B3基因在吲哚乙醛肟代謝途徑中的功能
前人的研究結果表明,CYP[STBX]79B2[STBZ]基因的過表達會使植物體內IAA含量明顯提高,但在cyp79B2和cyp79B3雙突變體中,植物則表現出幼苗下胚軸變短、長勢低矮的表型,體內IAA含量較低,合成速率降低[35]。擬南芥中吲哚乙醛肟途徑中IAA的生物合成過程已經明確,即CYP79B2/B3蛋白將色氨酸轉化為吲哚-3-乙醛肟,再經過吲哚-3-乙醛生成IAA,或者生成吲哚-3-乙腈,在腈水解酶作用下生成IAA[21,36]。
此外,研究還發現CYP79B2/B3雙突變體在 26 ℃ 條件下時IAA活性顯著下降,突變體植株呈現生長素缺乏表型[37];當溫度升高至29 ℃時,在剝離的胚軸中IAA的合成速率大幅度提高。這說明在擬南芥中,吲哚乙醛肟途徑的IAA合成還受到特定溫度的影響[38]。
有研究發現,將擬南芥CYP79B2基因與CYP79B3基因分別轉入大白菜中后,在轉CYP79B2基因的試驗中由于IAA的過量積累而未能獲得轉基因植株,而在轉CYP[STBX]79B3[STBZ]的試驗中獲得了轉基因植株,但表現出IAA過量合成的性狀[39-41]。以上結果說明,在IAA合成的過程中,與CYP79B3基因相比,CYP79B2基因可能發揮更重要的作用。而當這2個基因與吲哚族硫苷合成途徑中的CYP83B1基因一起被轉入大白菜中時,獲得的轉基因植株積累的吲哚族硫苷和表現型與對照相似。以上結果表明,CYP83B1基因的高效表達可以將大量IAOx引入吲哚族硫苷合成途徑,從而避免IAA的過量合成。
3 吲哚族硫苷與IAA代謝的關系
IAA的另一條合成途徑是色氨酸經由TAAs(色氨酸氨基轉移酶)與YUCCAs催化形成[34,40-41]。它與CYP79B2/CYP79B3催化的IAA合成途徑互補,所以在擬南芥cyp[STBX]79b2/cyp79b3[STBZ]雙突變體中仍能檢測到較高含量的IAA。然而,在擬南芥cyp[STBX]79b2/cyp79b3[STBZ]雙突變體中檢測不到吲哚族硫苷或亞麻薺素,表明由CYP79B2/B3催化的吲哚族硫苷或亞麻薺素的合成途徑是唯一的[28,42]。
吲哚族硫苷合成后大部分儲存在植物細胞組織液泡中,由于不具有揮發性,又與黑芥子酶存在空間上的隔離,因此結構較為穩定。當遭遇動物啃食或機械外力造成細胞破碎時,會與特定蛋白體中的黑芥子酶發生不可逆的水解反應,產生不同種類的生物活性物質。溫度、壓強、酸堿性和時間等多個因子對硫苷降解都有影響[43-44]。有研究發現,在酸性條件下(如pH值為2~5時)且存在亞鐵離子時,主要產生腈類物質,吲哚-3-乙腈經腈水解酶水解反應,最后生成吲哚-3-乙酸。
在生物合成途徑中,IAOx是主要代謝產物和次級代謝產物的重要分支點,其下游基因(CYP83B1、CS-裂合酶基因或UGT74B1基因)的丟失會導致IAA過量產生[45-46]。目前在擬南芥中已經鑒定出生長素含量增加的突變體,并且發現sur1可以編碼C-S裂解酶,參與吲哚族硫苷的合成[39]。而與之相似的sur2則可編碼CYP83B1。在sur2cyp83B1突變體中,IAOx轉化吲哚族硫苷的途徑被阻斷,導致IAOx更多地向IAA轉化[47]。以上分析說明,在吲哚族硫苷和IAA的合成過程中存在競爭反應底物的關系。近年來的生化分析結果表明,在擬南芥中大部分吲哚-3-乙醛肟是在CYP[STBX]79[STBZ]B途徑中產生的,推測吲哚-3-乙醛肟的合成與轉變為生長素的過程是耦合在一起的,而與吲哚-3-乙醛肟轉變為硫代葡萄糖苷的過程不是耦合在一起的[48]。色胺途徑產生的色胺以及植物體內形成的N-羥基色胺可抑制CYP83B1酶的活性,降低吲哚族硫苷的生成量,于是吲哚-3-乙醛肟參與到生長素合成過程中。吲哚族硫苷在Fe2+存在的條件下可水解生成吲哚-3-乙腈,經腈水合酶催化生成IAA,并誘導CYP83B1/sur2和UGT74B1基因的表達,說明IAA與吲哚族硫苷合成有協同調節作用[49]。
4 展望
亞麻薺素只存在于擬南芥、薺菜等少數十字花科植物中,在大白菜植株中尚未檢測到亞麻薺素。因此,大白菜可以作為研究吲哚族硫苷與IAA代謝平衡調控的模式植物。以吲哚族硫苷與IAA代謝的共同前體物吲哚乙醛肟為切入點,闡明CYP79B2/B3基因參與吲哚族硫苷(參與植物的病蟲害防御反應)與IAA(調控植物的生長發育過程)代謝平衡的分子機制,可為十字花科植物生長發育調控及其對害蟲、病原菌的防御研究提供理論基礎。
參考文獻:
[1]Grubb C D,Abel S. Glucosinolate metabolism and its control[J]. Trends in Plant Science,2006,11(2):89-100.
[2]Klopsch R,Witzel K,Brner A,et al. Metabolic profiling of glucosinolates and their hydrolysis products in a germplasm collection of Brassica rapa turnips[J]. Food Research International,2017,100(3):392-403.
[3]Bende J,Celenza J L. Indolic glucosinolates at the crossroads of tryptophan metabolism[J]. Phytochemistry Reviews,2009,8(1):25-37.
[4]Ishida M,Hara M,Fukino N,et al. Glucosinolate metabolism,functionality and breeding for the improvement of Brassicaceae vegetables[J]. Breeding Science,2014,64(1):48-59.
[5]Kliebenstein D J,Kroymann J,Brown P,et al. Genetic control of natural variation in Arabidopsis glucosinolate accumulation[J]. Plant Physiology,2001,126(2):811-825.
[6]Kim J H,Jander G. Myzus persicae (green peach aphid) feeding on Arabidopsis induces the formation of a repellent indole glucosinolate[J]. The Plant Journal,2007,49(6):1008-1019.
[7]Bednarek P,Pislewska-Bednarek M,Svatos A,et al. A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense[J]. Science,2009,323(5910):101-106.
[8]Clay N K,Adio A M,Denoux C,et al.Glucosinolate metabolism required for an Arabidopsis innate immune response[J]. Science,2009,323(5910):95-101.
[9]Murillo G,Mehta R G. Cruciferous vegetables and cancer prevention[J]. Nutrition & Cancer,2001,41(1/2):17-28.
[10]Mithen R F,Dekker M,Verkerk R,et al.Review:the nutritional significance,biosynthesis and bioavailability of glucosinolates in human food[J]. Journal of the Science of Food & Agriculture,2000,80(7):967-984.
[11]Fahey J W,Zhang Y,Talalay P,et al. Broccoli sprouts:an exceptionally rich source of inducers of enzymes that protect against chemical[J]. PNAS,1997,94(19):10367-10372.
[12]Chen I,Mc Dougal A,Wang F,et al. Aryl hydrocarbon receptor rmediated antiestrogenic and antitumorigenic activity of diindolylmethane[J]. Carcinogenesis,1998,19:1631-1639.
[13]Kliebenstein D J,Gershenzon J,Mitchell-Olds T,et al.Comparative quantitative trait loci mapping of aliphatic,indolic and benzylic glucosinolate production in Arabidopsis thaliana leaves and seeds[J]. Genetics,2001,159(1):359-370.
[14]Brown P D,Tokuhisa J G,Reichelt M,et al.Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana[J]. Phytochemistry,2003,62(3):471-481.
[15]李一蒙,陳亞州,閻秀峰. 植物中的吲哚族芥子油苷與生長素代謝途徑的關系[J]. 植物生理學報,2009,45(2):195-201.
[16]Mikkelsen M D,Naur P,Halkier B A,et al. Arabidopsis mutants in the C-S lyase of glucosinolate biosynthesis establish a critical role for indole-3-acetaldoxime in auxin homeostasis[J]. Plant Journal,2004,37(5):770-777.
[17]Bak S,Feyereisen R.The involvement of two P450 enzymes CYP83B1 and CYP83A1 in auxin homeostasis and glucosinolate biosynthesis[J]. Plant Physiology,2001,127(1):108-118.
[18]Petersen B L,Andréasson E,Bak S,et al. Characterization of transgenic Arabidopsis thaliana with metabolically engineered high levels of p-hydroxybenzylglucosinolate[J]. Planta,2001,212(4):612-618.
[19]Snderby I E,M Burow,Rowe H C,et al. A complex interplay of three R2R3-MYB transcription factors determines the profile of aliphatic glucosinolates in Arabidopsis[J]. Plant Physiology,2010,153(1):348-368.
[20]Bender J,Fink G R.A Myb homologue,ATR1,activates tryptophan gene expression in Arabidopsis[J]. PNAS,1998,95(10):5655-5660.
[21]Celenza J L,Quiel J A,Smolen G A,et al. The Arabidopsis ATR1 Myb transcription factor controls indolic glucosinolate homeostasis[J]. Plant Physiology,2005,137(1):253-262.
[22]Gigolashvili T,Berger B,Mock H,et al.The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana[J]. Plant Journal,2007,50(5):886-901.
[23]Hai D U,Ran F,Liu J,et al. Genome-wide expression analysis of glucosinolate biosynthetic genes in Arabidopsis across diverse tissues and stresses induction[J]. Scientia Acricultura Sinica,2016,49(15):2879-2897.
[24]Schweizer F,Fernándezcalvo P,Zander M,et al. Arabidopsis basic helix-loop-helix transcription factors MYC2,MYC3,and MYC4 regulate glucosinolate biosynthesis,insect performance,and feeding behavior[J]. Plant Cell,2013,25(8):3117-3132.
[25]Skirycz A,Reichelt M,Burow M,et al.DOF transcription factor AtDof1.1 (OBP2) is part of a regulatory network controlling glucosinolate biosynthesis in Arabidopsis[J]. Plant Journal,2006,47(1):10-24.
[26]Levy M,Wang Q,Kaspi R,et al. Arabidopsis IQD1,a novel calmodulin-binding nuclear protein,stimulates glucosinolate accumulation and plant defense[J]. Plant Journal,2005,43(1):79-96.
[27]Hull A K,Vij R,Celenza J L,et al. Arabidopsis cytochrome P450s that catalyze the first step of tryptophan-dependent indole-3-acetic acid biosynthesis[J]. PNAS,2000,97(5):2379-2384.
[28]Zhao Y,Hull A K,Gupta N R,et al. Trp-dependent auxin biosynthesis in Arabidopsis:involvement of cytochrome P450s CYP79B2 and CYP79B3[J]. Genes Dev,2002,16(23):3100-3112.
[29]Mikkelsen M D,Hansen C H,Wittstock U,et al. Cytochrome P450 CYP79B2 from Arabidopsis catalyzes the conversion of tryptophan to indole-3-acetaldoxime,a precursor of indole glucosinolates and indole-3-acetic acid[J]. Journal of Biological Chemistry,2000,275(43):33712-33717.
[30]Mikkelsen M D,Halkier B A. Modulation of CYP[STBX]79[STBZ] genes and glucosinolate profiles in Arabidopsis by defense signaling pathways[J]. Plant Physiology,2003,131(1):298-308.
[31]Bandurski R S,Cohen J D,Slovin J P,et al. The plant hormones:auxin biosynthesis and metabolism[M]. Berlin,Heidelberg:Springer-Verlag,1995:39-65.
[32]Bartel B,Leclere S,Magidin M,et al. Inputs to the active indole-3-acetic acid pool:de novo synthesis,conjugate hydrolysis,and indole-3-butyric acid β-oxidation[J]. Journal of Plant Growth Regulation,2001,20(3):198-216.
[33]Normanly J,Bartelt B. Redundancy as a way of life:IAA metabolism[J]. Current Opinion in Plant Biology,1999,2(3):207-213.
[34]Mashiguchi K,Tanaka K,Sakai T,et al. The main auxin biosynthesis pathway in Arabidopsis[J]. PNAS,2011,108(45):18512-18517.
[35]Quittenden L J,Davies N W,Smith J A,et al. Auxin biosynthesis in pea:characterization of the tryptamine pathway[J]. Plant Physiology,2009,151(3):1130-1138.
[36]Tam Y Y,Normanly J. Overexpression of a bacterial indole-3-acetyl-l-aspartic acid hydrolase in Arabidopsis thaliana[J]. Physiologia Plantarum,2010,115(4):513-522.
[37]Nemoto K,Hara M,Suzuki M,et al. The NtAMI[STBX]1[STBZ] gene functions in cell division oftobacco BY-2 cells in the presence of indole-3-acetamide[J]. Febs Letters,2009,583(2):487-492.
[38]Pollmann S,Düchting P,Weiler E W. Tryptophan-dependent indole-3-acetic acid biosynthesis by‘IAA-synthase proceeds via indole-3-acetamide[J]. Phytochemistry,2009,70(4):523-531.
[39]Tao Y,Ferrer J L,Ljung K,et al. Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants[J]. Cell,2008,133(1):164-176.
[40]Won C,Shen X,Mashiguchi K,et al. Conversion of tryptophan to indole-3-acetic acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis[J]. PNAS,2011,108(45):18518-18523.
[41]Zhao Y D. Auxin biosynthesis:a simple two-step pathway converts tryptophan to indole-3-acetic acid in plants[J]. Molecular Plant,2012,5(2):334-338.
[42]Glawischnig E,Hansen B G,Olsen C E,et al. Camalexin is synthesized from indole-3-acetaldoxime,a key branching point between primary and secondary metabolism in Arabidopsis[J]. PNAS,2004,101(21):8245-8250.
[43]Wittstock U,Halkier B A. Glucosinolate research in the Arabidopsis era[J]. Trends in Plant Science,2002,7(6):263-270.
[44]陳凌云,余芳潔,陳君杰,等. 植物硫代葡萄糖苷二次修飾及調控的研究進展[J]. 江蘇農業科學,2019,47(4):22-29.
[45]Bttcher C,Westphal L,Schmotz C,et al. The multifunctional enzyme CYP71B15 (PHYTOALEXIN DEFICIENT3) converts cysteine-indole-3-acetonitrile to camalexin in the indole-3-acetonitrile metabolic network of Arabidopsis thaliana[J]. Plant Cell,2009,21(6):1830-1845.
[46]Barlier I,Kowalczyk M,Marchant A,et al. The SUR[STBX]2[STBZ] gene of Arabidopsis thaliana encodes the cytochrome P450 CYP83B1,a modulator of auxin homeostasis[J]. PNAS,2000,97(26):14819-14824.
[47]張海峰,袁 晶,汪俏梅. 植物激素與芥子油苷在生物合成上的相互作用[J]. 細胞生物學雜志,2005,27(4):423-426.
[48]王家利,劉冬成,郭小麗,等. 生長素合成途徑的研究進展[J]. 植物學報,2012,47(3):292-301.
[49]阮 穎,周樸華,劉春林. 硫代葡萄糖苷合成核心途徑與植物生長素微調[J]. 生命科學研究,2006(增刊3):25-29.
收稿日期:2019-08-20
基金項目:國家自然科學基金(編號:31572130、31000916);浙江省自然科學基金(編號:LY14C150005、Y3090538)。
作者簡介:孫夏莉(1990—),女,浙江舟山人,碩士研究生,研究方向為蔬菜品質調控與分子機理。E-mail:1367201782@qq.com。
通信作者:臧運祥,博士,教授,碩士生導師,研究方向為園藝作物品質生理、生物技術與分子育種。E-mail:yxzang@zafu.edu.cn。
