EDDS對Cd脅迫下三葉鬼針草幼苗NO產生的影響
2020-08-24 14:23:46丁浚剛陳銀萍楊波師小平頡海帆
生態科學 2020年4期
丁浚剛, 陳銀萍, 楊波, 師小平, 頡海帆
EDDS對Cd脅迫下三葉鬼針草幼苗NO產生的影響
丁浚剛, 陳銀萍*, 楊波, 師小平, 頡海帆
蘭州交通大學環境與市政工程學院, 蘭州 730070
通過室內盆栽試驗, 研究了40 mg·kg-1Cd(CdCl2·2.5 H2O)脅迫下, 不同濃度乙二胺二琥珀酸(EDDS)(0、0.5、1.5、2.5、5.0 mmol·L-1)單施及EDDS與一氧化氮(NO)供體硝普鈉(SNP)(0、0.25、0.5、1.0 mmol·L-1)聯合施加對三葉鬼針草(L)幼苗應激信號分子NO產生量和一氧化氮合酶(NOS)活性的影響。結果表明: 單施EDDS, 植株不同部位NO生成量隨EDDS濃度的升高呈增加趨勢, 5.0 mmol·L-1時達到最大; 0.5 mmol·L-1的EDDS可增強根、葉中NOS活性。在探究NO產生較多和NOS活性增強顯著的EDDS處理濃度與SNP聯合施加的研究中發現, 隨SNP濃度的升高, 根中NO生成量先升高后降低, 莖和葉中持續升高; 適宜濃度的SNP可進一步增強植株體內NOS活性。EDDS誘導NO的生成會被硝酸還原酶(NR)抑制劑(NaN3)和NOS抑制劑(L-NAME)抑制, 對EDDS處理下NOS活性影響較小。NO清除劑(c-PTIO)能有效清除部分NO, 增強根和葉中NOS活性。因此, 在Cd脅迫下, 適宜濃度的EDDS單施及與SNP聯合施加都會增加三葉鬼針草幼苗體內NO產生量。
EDDS; NO; 三葉鬼針草; Cd脅迫; NOS;
0 前言
植物修復是一種生態友好型原位綠色修復技術, 是重金屬污染土壤修復研究的熱點。但因普通植物生物量低、生長緩慢、對重金屬的積累具有專一性等缺點, 很大程度上限制了植物修復技術在重金屬污染土壤治理方面的推廣應用[1]。近年來, 螯合劑誘導植物提取技術受到極大關注[2], 研究證實, Cd、鉛(Pb)脅迫下, 適宜濃度的螯合劑可使土壤中水溶態重金屬含量增加, 同時苧麻(L.)、紫花苜蓿(L.)等對土壤中重金屬的吸收能力得到強化, 植物體內重金屬含量明顯提高, 土壤中重金屬的去除效率得到了提高[3–4]。研究發現, 植物激素、菌根等與螯合劑聯合施用, 不僅能夠促進植物在重金屬污染土壤中的正常生長, 提高植物抗性, 改善土質, 而且能夠強化植物對重金屬的吸收和富集, 大大提升了植物的修復效果[5]。
EDDS因其生物可降解, 對環境較友好, 且對植物和土壤微生物毒性較小, 作為一種可替代乙二胺四乙酸(EDTA)的螯合劑近年來廣泛用于污染土壤修復的研究[6]。NO是一種廣泛存在并參與植物應激反應的早期信號分子, 環境脅迫可導致植物細胞的NO爆發, 使植物細胞啟動一系列反應或與其它信號分子協同, 防衛和保護植物體免受脅迫損傷[7]。在重金屬脅迫條件下的研究表明, NO會參與抗逆生理過程, 誘導增強植物體內NO合成主要途徑NOS活性, 提升植物的抗逆性, 降低副產物的積累, 提高植物對環境的適應能力[8]。但NO能否像激素和菌根等一樣作為聯合物質協同螯合劑強化植物對重金屬的吸收和富集鮮有報道。
三葉鬼針草(L.)是種一年生草本植物, 由于其生物量大, 根系較發達, 對重金屬表現出較強的抗逆性、優良的富集性和轉移性, 近年來廣泛用于Cd、Pb污染的土壤修復研究中[9–10]。本文通過盆栽試驗研究不同濃度EDDS單施及與NO供體聯合作用對Cd脅迫下三葉鬼針草不同部位NO生成及NOS活性的影響, 分析EDDS聯合NO供體能否促進植物體內NO生成, 并淺探NO生成途徑, 為進一步研究EDDS與聯合物質是否可以通過促進植物體內NO的生成來提高植物抗性, 進而為強化植物對重金屬的吸收和富集提供理論基礎, 也可為促進三葉鬼針草在Cd污染土壤植物修復中的實際應用提供科學依據。
1 材料與方法
1.1 材料與試劑
三葉鬼針草種子購自泰山野菜種植基地; EDDS為麥卡希試劑, 現用現配; NR抑制劑NaN3、NOS抑制劑L-NAME、c-PTIO均購自Sigma公司; NOS和NO試劑盒均購自南京建成生物工程研究所; CdCl2·2.5 H2O、Na2[Fe(CN)5NO]·2H2O及其他藥品均為國產分析純。
1.2 試驗設計
試驗土壤基質采自甘肅省某縣周邊農田耕地(0—20 cm, Cd含量0.49 mg·kg-1), 自然風干后過2 mm篩備用。將2 kg備用土壤與設定污染物(以Cd Cl2·2.5H2O 模擬Cd脅迫, 濃度為40 mg·kg-1, 溶液形式加入到土壤中)充分混勻后裝入花盆(口徑20 cm、高度15 cm), 平衡21 d后, 于2016年3月20日選種播種, 盆栽置于溫度、光照適宜的培養架上進行培養, 發芽后間苗至每盆10株植物, 待三葉鬼針草生長60 d左右、長出6片真葉后, 于2016年5月20日前后進行如下處理:
①EDDS單施試驗設五個處理: 配制濃度為0、0.5、1.5、2.5、5.0 mmol·L-1的EDDS溶液(pH 7.0), 分別取各濃度溶液100 mL施入已長好植物的土壤中; ②選取NO產生量較多和NOS活性較強的EDDS處理濃度分別與不同濃度SNP(0、0.25、0.5、1.0 mmol·L-1)共同施加, 進行EDDS與SNP聯合作用試驗, 與0.1 mmol·L-1NaN3+0.25 mmol·L-1L-NAME共同施加, 進行NO主要合成途徑以及對NOS活性影響試驗, 與0.075 mmol·L-1c-PTIO共同施加, 進行EDDS誘導生成NO的清除及NOS活性影響試驗; 各處理對照為同濃度Cd脅迫、相同生長條件下施加同劑量超純水的處理(K(Cd))。上述處理各設置3個重復, 每個重復3個平行, 處理7 d后, 取現采鮮樣進行各指標測定。
1.3 試驗方法
使用南京建成NO試劑盒法測定NO含量: 稱取根、莖、葉1 g洗凈鮮樣, 加入提取液冰浴研磨, 加入混合試劑37℃標準水浴1 h, 依次加入試劑三、四, 3500 rpm離心10 min, 取樣加入顯色劑, 10 min后550 nm波長下測定并計算NO含量。南京建成NOS試劑盒法測定NOS活性: 稱取鮮樣0.5 g加入提取液進行冰浴研磨, 4℃下3500 rpm離心10 min, 依次加入說明書要求試劑, 530 nm波長下測定并計算NOS活性。
1.4 數據統計與分析
采用Excel和SPSS 24軟件進行數據處理和分析, 利用Origin 9軟件繪圖。采用單因素方差分析(One-way ANOVA)和Duncan法進行多重比較。
2 結果與分析
2.1 EDDS對Cd脅迫下三葉鬼針草幼苗NO產生的影響
40 mg·kg-1Cd脅迫下, 隨著EDDS濃度增加不同部位NO生成量呈增加趨勢; 根中NO生成量顯著增加, 5.0 mmol·L-1時達到最大, 較K(Cd)增加了70.07%(P<0.05); EDDS濃度為2.5和5.0 mmol·L-1時莖中NO生成量增加顯著, 較K(Cd)分別增加234.18%和268.00%(P<0.05); 葉中NO生成量在EDDS濃度為2.5 mmol·L-1時達到最大, 較K(Cd)增加155.14%(P<0.05), 雖在5.0 mmol·L-1時出現下降態勢, 但仍高于K(Cd); EDDS濃度相同的條件下, 不同部位NO生成量根>莖>葉(圖1)。
Cd脅迫下, 相同濃度SNP處理下, 三葉鬼針草幼苗根和莖在5.0 mmol·L-1EDDS處理下生成的NO量高于2.5 mmol·L-1, 但隨SNP濃度的升高各部位總體變化趨勢相似(圖2)。根中NO生成量隨SNP濃度的升高呈現先升高后降低的趨勢, 在高濃度SNP與EDDS聯合作用時NO生成量會被抑制, 2.5 mmol·L-1EDDS與0.25 mmol·L-1SNP聯合作用時NO生成量達到最大, 較單施同濃度EDDS增加40.50%(P<0.05); 5.0 mmol·L-1EDDS與0.5 mmol·L-1SNP聯合作用時NO生成量達到最大, 較單施同濃度EDDS增加118.39%(P<0.05)(圖2A)。
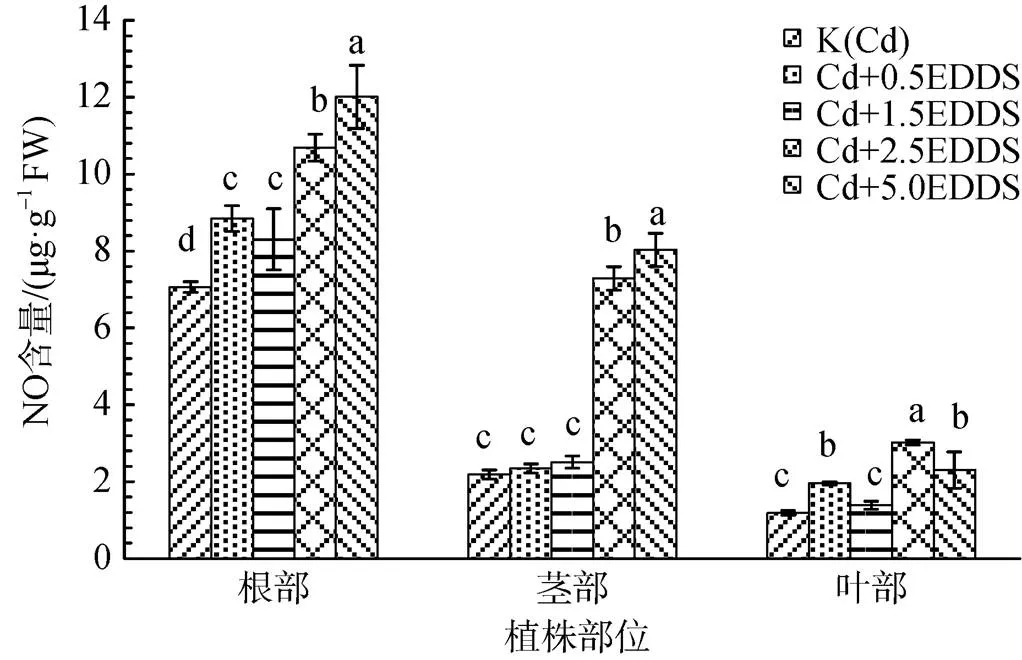
不同小寫字母表示各處理間差異顯著( P<0.05)。下同。
Figure 1 Effect of EDDS on NO production inseedlings
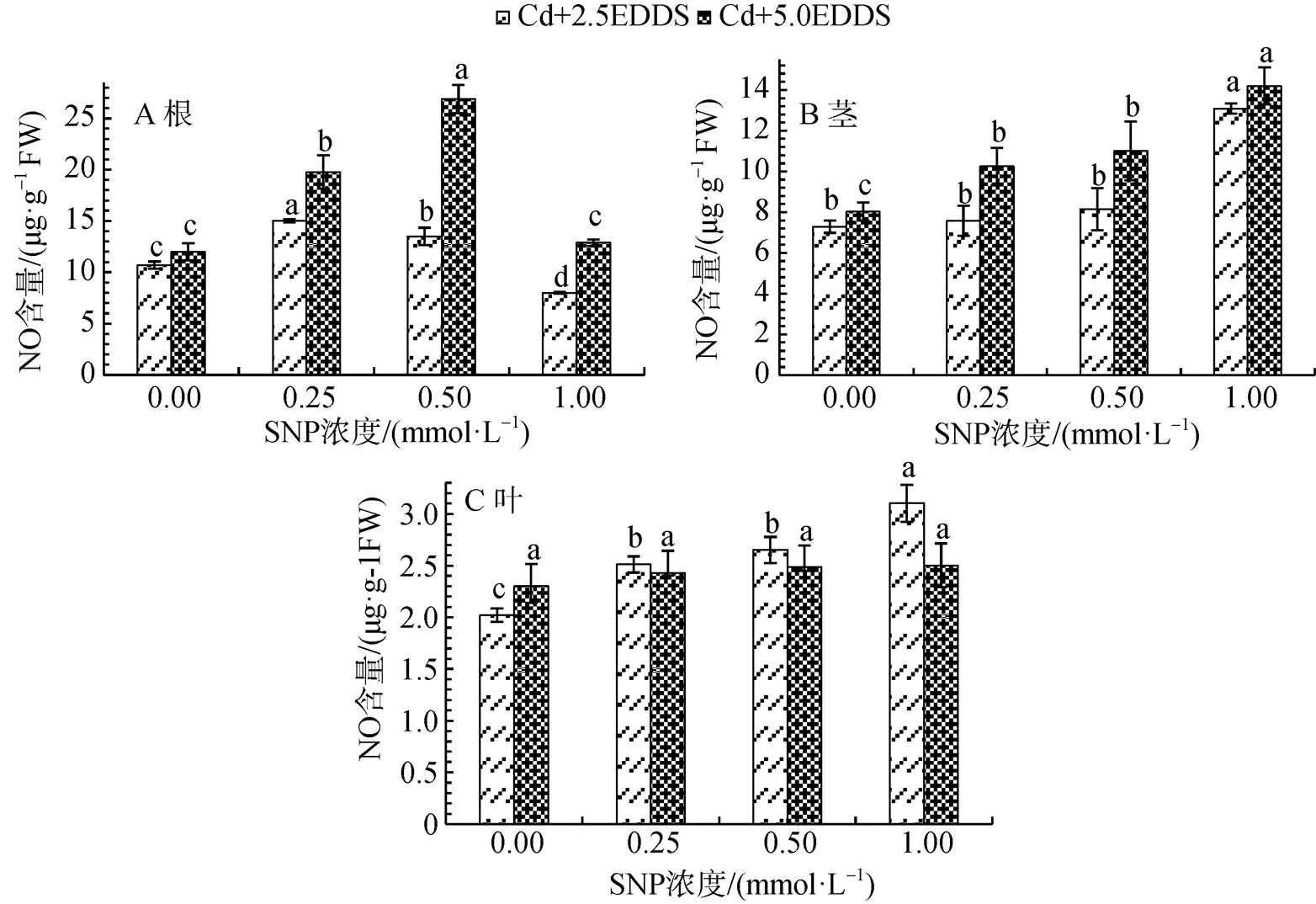
圖 2 EDDS聯合SNP對Cd脅迫下三葉鬼針草幼苗不同部位NO生成的影響
Figure 2 Effects of EDDS combined with SNP on NO production in different parts ofseedlings
莖部NO生成量隨SNP濃度的升高呈現持續升高的趨勢。當2.5和5.0 mmol·L-1EDDS分別與1.0 mmol·L-1SNP聯合作用時NO生成量分別達到最大值, 較單施EDDS分別增加79.54%和76.82% (P<0.05)(圖2B)。
葉部NO生成量隨SNP濃度的升高呈現持續升高的趨勢。2.5 mmol·L-1EDDS與1.0 mmol·L-1SNP聯合作用NO生成量達到最大值, 較單施EDDS增加53.61%(P<0.05); 而5.0 mmol·L-1EDDS與不同濃度SNP聯合作用時NO生成量雖有增加但不顯著(圖2C)。
對2.5 mmol·L-1EDDS處理組添加NaN3和L-NAME發現, 兩種抑制劑能有效降低根和莖中由誘導產生的NO產生量; c-PTIO的施加能有效清除植株體內生成的部分NO, 根中NO生成量較K(Cd)顯著降低(P<0.05), 莖中NO生成量較K(Cd)無顯著變化(P>0.05)。兩種抑制劑的添加對葉中誘導產生的NO生成量影響不顯著, 而c-PTIO的添加可顯著降低部分EDDS誘導產生的NO(圖3)。
2.2 EDDS對三葉鬼針草不同部位NOS活性的影響
探究EDDS對三葉鬼針草幼苗NOS活性影響的研究發現: 0.5 mmol·L-1EDDS使得根和葉中NOS活性顯著增強, 較K(Cd)分別提高14.88%和30.57% (P<0.05); 5.0 mmol·L-1EDDS處理顯著增強莖中NOS活性, 較K(Cd)提高48.00%(P<0.05), 而對根中NOS活性產生明顯的抑制作用(圖4)。綜合結果表明, 低濃度EDDS顯著增強根和葉中NOS活性, 對莖影響較小; 高濃度EDDS抑制根中NOS活性, 增強莖中NOS活性, 對葉影響較小。
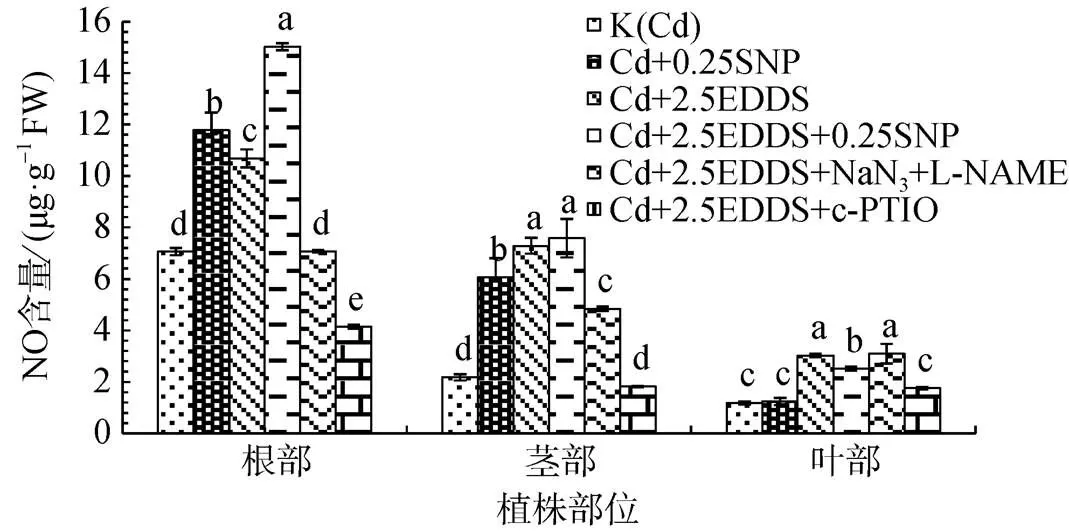
圖3 EDDS與NO產生劑、抑制劑和清除劑聯合作用對三葉鬼針草幼苗NO生成的影響
Figure 3 Effects of EDDS combined with NO producers, inhibitors and scavengers on NO production inseedlings
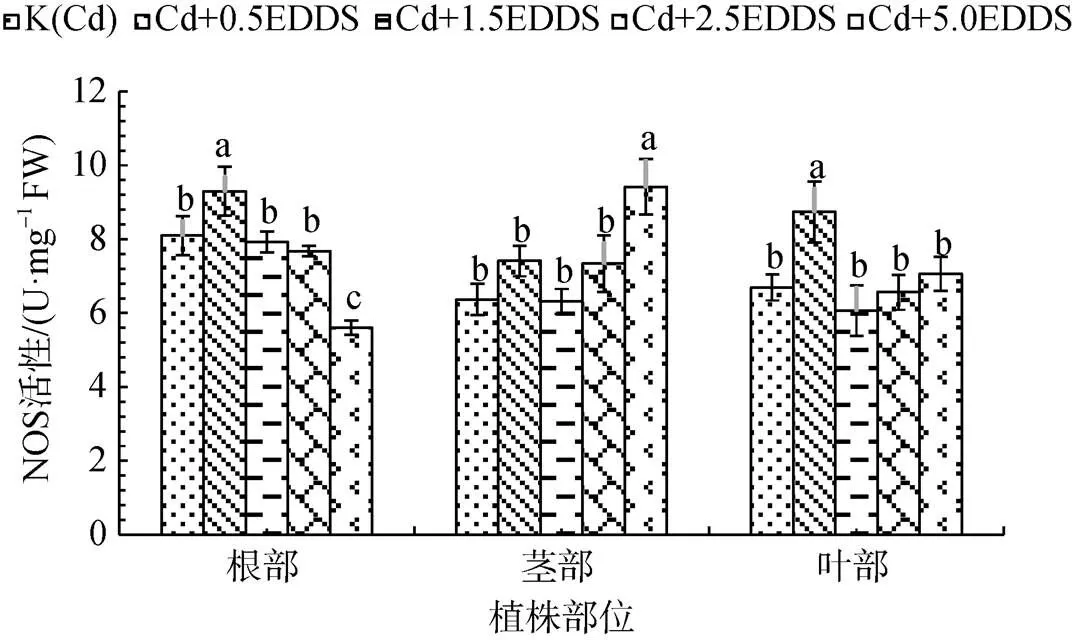
圖4 不同濃度EDDS對三葉鬼針草幼苗NOS活性的影響
Figure 4 Effect of different concentrations of EDDS on NOS activities inseedlings
選取對三葉鬼針草幼苗不同部位NOS活性誘導作用較顯著的0.5 mmol·L-1EDDS與不同濃度SNP聯合處理幼苗發現: 三葉鬼針草不同部位NOS活性隨著SNP濃度的增加, 呈現出先升高后降低的趨勢, 且在0.25 mmol·L-1時各部位NOS活性均最強, 且顯著高于K(Cd)(P<0.05)。高濃度SNP對根中NOS活性影響不顯著; SNP濃度變化對莖部NOS活性影響較小; 0.25 mmol·L-1時, 葉中NOS活性較K(Cd)提高71.82%(P<0.05), 而施加1.0 mmol·L-1SNP處理時, NOS活性受到了顯著抑制(P>0.05)(圖5)。
在0.5 mmol·L-1EDDS作用下, NaN3和 L- NAME的添加對三葉鬼針草幼苗NOS活性影響較小, c-PTIO的添加可顯著增強根中NOS活性(P<0.05), 對莖中NOS活性具有明顯的抑制作用, 對葉中NOS活性影響甚微(圖6)。
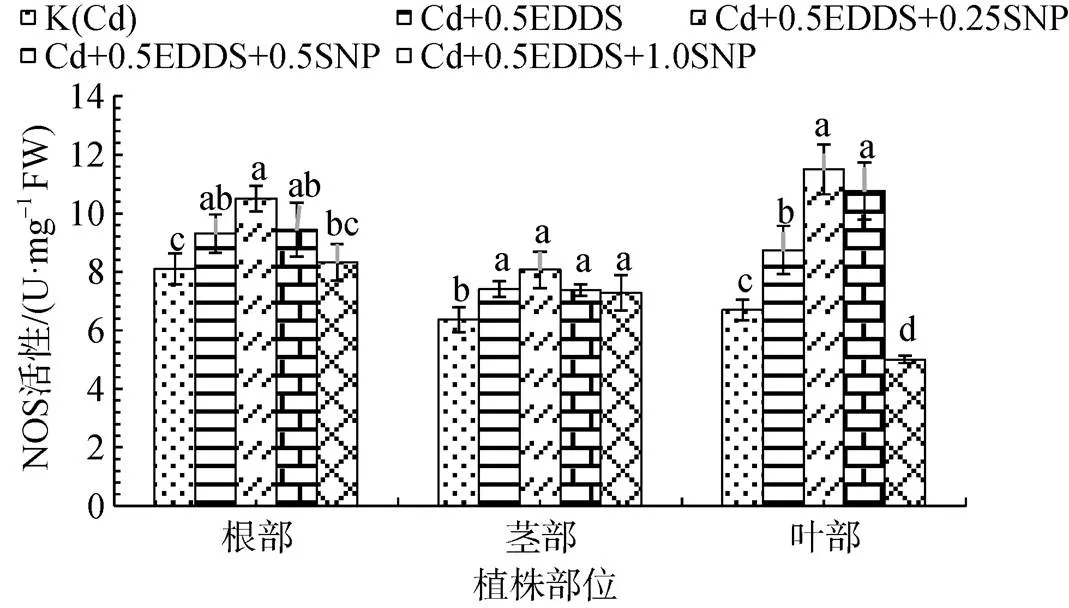
圖5 不同濃度SNP與0.5 mmol·L-1EDDS共同施加對三葉鬼針草不同部位NOS活性的影響
Figure 5 Effect of different concentrations of SNP and 0.5 mmol·L-1EDDS on NOS activities in different parts ofseedlings
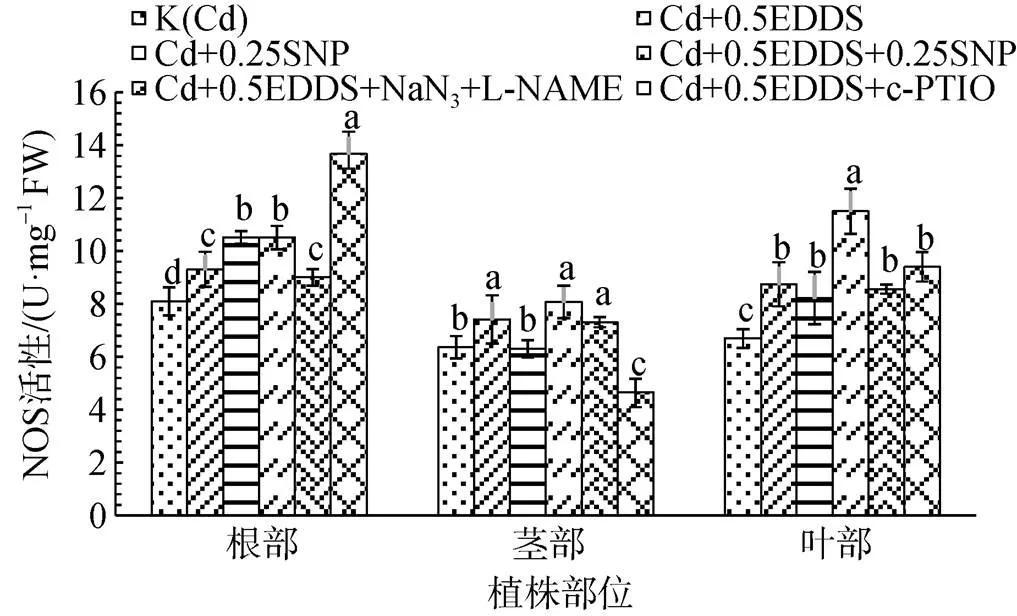
圖6 EDDS與NO產生劑、抑制劑和清除劑聯合作用對三葉鬼針草幼苗NOS活性的影響
Figure 6 Effect of EDDS combined with NO generator, inhibitor and scavenger on NOS activity ofseedlings
3 討論
3.1 EDDS對三葉鬼針草不同部位NO含量的影響
螯合誘導植物修復是通過螯合劑活化并釋放土壤中固相重金屬, 提高土壤中液相重金屬離子濃度, 解吸吸附在土壤顆粒表面的重金屬, 從而提高植物修復效率[11]。適宜濃度的EDDS可以有效促進土壤中碳酸鹽結合態、鐵錳氧化物結合態等形態的金屬離子向可交換態轉化, 促進植物對金屬離子的吸收[12], 并通過調整植物根系各化學成分、可溶性糖、可溶性蛋白質、丙二醛等的含量以及根系對礦物質元素的吸收和分配等來增強根系對重金屬的耐受能力, 提升植物對重金屬的吸收和富集能力[13]。研究證實, 植物體內NO主要通過改變重金屬的分布和誘導抗性反應來參與植物對重金屬脅迫的抗性[7]。紫花苜蓿和東南景天(Hance)在低濃度Cd脅迫下, 內源NO含量急劇上升, 而隨著Cd濃度的升高, NO含量顯著下降, 維持在略高或持平于空白對照, 在清除NO后植物的正常生長會受到明顯的抑制, 并且活性氧(ROS)的積累會明顯增加, 細胞損傷程度加重[14–15]。本研究結果表明, Cd脅迫下, 植物體內不同部位NO含量隨著EDDS濃度的升高而升高, 一方面可能是隨著EDDS濃度的增加使得土壤中可被植物利用的重金屬形態含量增加, 促進了植物對重金屬的吸收[16], 導致在一定重金屬濃度范圍下植物體內NO生成量增加, 以減輕重金屬對植物的毒害作用[17]; 另一方面, 高濃度EDDS會對植物的正常生長產生抑制作用[18], 結合本試驗室已有研究成果[10], EDDS濃度不超過2.5 mmol·L-1時, 三葉鬼針草能正常生長, 吸收富集轉運重金屬能力較強, 而超過2.5 mmol·L-1時, 植物正常生長受到抑制, 植物對Cd的吸收量減少, 土壤中Cd含量與對照組相比無顯著差異, 說明高濃度EDDS對三葉鬼針草產生脅迫作用, 使得植物體內在NO含量積累升高, 而過多產生的NO與ROS反應產生毒性更強的氧化物如過氧化亞硝酸(ONOO-), 對細胞膜的損傷程度更大, 這與滕玉瑾等[19]的研究結果相似。
外源NO能夠增強植物對重金屬脅迫抗性, 一方面是通過增強植物細胞的抗氧化系統或直接清除ROS, 另一方面是通過影響植物對重金屬的吸收以及重金屬在植物細胞內的分布[20]。外源NO供體SNP的施加使植物體內NO生成增加, 一方面是由于植物吸收SNP后在植物體內本身釋放NO[21], 另一方面, L-精氨酸作為植物體內通過NOS途徑合成NO的主要前體物質, 其含量變化對植物體內NO含量影響較為明顯[22], 而已有研究證實, 在銅(Cu)脅迫條件下, 施加SNP可以提高植物葉和根中L-精氨酸含量, 促進其向NO合成方向進行[23]。本研究表明, 在Cd脅迫條件下, 不同濃度的SNP與EDDS共同施加, 三葉鬼針草不同部位NO生成量會進一步增加, 隨SNP濃度的增加, 莖和葉中NO生成量呈持續增加態勢, 而根中NO生成量呈先增加后降低趨勢, 其原因可能是在高濃度EDDS存在條件下, SNP濃度超過0.25 mmol·L-1時增強了重金屬對植物根部生長的抑制作用, 且隨著SNP濃度的升高, 抑制作用越明顯[18,24]。
c-PTIO是NO的清除劑, 可有效清除植物內源NO[25], NaN3和L-NAME分別是NR和NOS的抑制劑[26]。研究表明, 植株體內NO釋放量與NOS活性呈正相關, 添加適宜濃度SNP可以不同程度誘導植株體內NOS活性, 從而增加植株體內NO含量, NOS抑制劑L-NAME可以明顯抑制NO的產生[23,27]; 本研究用EDDS聯合NaN3和L-NAME處理Cd污染土壤中種植的三葉鬼針草幼苗, 發現三葉鬼針草不同部位NO含量相對于施加EDDS和SNP聯合施加處理組都有所下降; 經c-PTIO處理后, 根部NO含量明顯低于K(Cd)以及其他處理組, 葉和莖中NO生成量均有所降低, 說明在EDDS和SNP誘導作用下能夠生成較多的NO, 而兩種酶抑制劑能夠在一定程度上抑制這種作用, c-PTIO則能夠有效清除部分NO。楊建軍等[28]的研究證實了鹽脅迫條件能夠使黃瓜(L.)幼苗體內NO生成量增加, NO清除劑c-PTIO以及NaN3和L-NAME都可以不同程度地降低NO含量, 這與本研究結果一致。
3.2 EDDS對三葉鬼針草不同部位NOS活性的影響
植物體內的內源NO主要由NOS和NR催化合成以及非酶促途徑合成[29]。由NOS介導時以L-精氨酸為底物, 以NAD(P)H為電子供體生成NO[7,30]; 此外, 植物細胞質的NR和根部特有的質膜亞硝酸還原酶( Ni-NOR)也參與NO的生成, NR和Ni-NOR以NAD(P)H作為電子供體進行還原反應將亞硝酸鹽(NO2-)還原生成NO[22]。研究證實, 在Cd脅迫下, 適宜濃度的SNP配合24-表油菜素內酯(EBR)可以刺激花生(L.)體內的NR和NOS, 提高酶的活性, 配合植物體內其他生理變化, 來減輕Cd在花生體內的負面效應[31]。本研究表明, 低濃度EDDS可以誘導三葉鬼針草根、莖、葉中NOS活性, NO含量也隨之升高, 說明低濃度EDDS處理的三葉鬼針草在Cd脅迫下, NOS途徑會對NO的生成產生影響, 而高濃度的EDDS對根部NOS活性產生了明顯的抑制作用; 陳銀萍等[8]的研究結果表明, 隨著Pb脅迫濃度的增加, 三葉鬼針草不同部位NOS酶活性呈現先升高后降低的趨勢。在高濃度EDDS處理時, 根中NOS活性雖然受到明顯的抑制并低于K(Cd), 而NO含量持續升高, 說明此時, 三葉鬼針草根中NOS途徑不是合成NO的唯一途徑; St?hr等[32]的研究發現, 煙草()根部存在特有的Ni-NOR, 參與NO的生成。在低濃度EDDS處理時, 施加SNP, 可以增強NOS活性, 但高濃度SNP對根和莖中NOS活性影響較小, 該結果與劉建新等[33]研究結果相似。低濃度的EDDS與L-NAME 和NaN3聯合處理時根中NOS活性較SNP、EDDS及二者聯合處理組有所降低, 莖和葉中也呈現出類似的變化趨勢, 而在施加c-PTIO后根部NOS活性明顯升高, 葉中較K(Cd)也有顯著的升高, 這與c-PTIO導致鹽處理小麥(L.)根中NOS類似蛋白活性增強[20]和c-PTIO使機械損傷下蘆葦()愈傷組織NOS活性增強[34]的研究結果相似。而莖中呈現出明顯的抑制狀態, 原因可能是同種植物不同部位NOS對外界干擾的響應機理存在差異, 具體原因需進一步研究。
4 結論
Cd脅迫下, EDDS能誘導三葉鬼針草幼苗NO生成量的增加, 低濃度EDDS提升了根和葉中NOS活性, 且NO含量增加, 高濃度EDDS對根中NOS活性產生抑制, 但NO含量升高, 證明NOS途徑不是根部NO合成的唯一途徑; 低濃度EDDS與SNP聯合作用可進一步提升根部NOS活性, 且在一定濃度范圍內NO的生成量也會增加。兩種抑制劑的添加進一步證實了EDDS對NO生成能產生積極的誘導作用。NO的清除能刺激NOS活性, 證實了在一定條件下, NOS途徑是植物體內NO生成的途徑之一, 但在不同部位表現出不同的響應, 出現這種現象的原因需進一步研究。
[1] 聶亞平, 王曉維, 萬進榮, 等. 幾種重金屬(Pb、Zn、Cd、Cu)的超富集植物種類及增強植物修復措施研究進展[J]. 生態科學, 2016, 35(02): 174–182.
[2] 袁金瑋, 陳笈, 陳芳, 等. 強化植物修復重金屬污染土壤的策略及其機制[J]. 生物技術通報, 2019, 35(01): 126– 136.
[3] 劉金, 殷憲強, 孫慧敏, 等. EDDS與EDTA強化苧麻修復鎘鉛污染土壤[J]. 農業環境科學學報, 2015, 34(7): 1293–1300.
[4] 任麗娟, 柴琳琳, 郭昱, 等. 螯合劑對鎘在土壤和紫花苜蓿中積累的影響研究[J]. 新疆大學學報(自然科學版), 2015, (3): 330–335.
[5] MOHD I, SAHI V. Promising role of plant hormones in translocation of lead inshoots[J]. Environmental Pollution, 2008, 153(1): 29–36.
[6] 劉曉娜, 趙中秋, 陳志霞, 等. 螯合劑、菌根聯合植物修復重金屬污染土壤研究進展[J]. 環境科學與技術, 2011, 34(12H): 127–133.
[7] 張玲玲, 吳丹, 趙子捷, 等. 植物一氧化氮信號分子的研究進展[J]. 植物學報, 2017, 52(3): 337–345.
[8] 陳銀萍, 柯昀琪, 楊志娟, 等. 鉛脅迫下三葉鬼針草內源一氧化氮的生成及其對氧化損傷的緩解效應[J]. 植物科學學報, 2018, 36(2): 264–272.
[9] 諶金吾, 孫一銘, 王鳳英, 等. 三葉鬼針草毛狀根的誘導及其對重金屬Cd、Pb蓄積[J]. 環境科學學報, 2015, 35(5): 1596–1602.
[10] 楊波, 陳銀萍, 柯昀琪, 等. EDDS對Cd脅迫下三葉鬼針草生長和抗氧化酶系統及Cd積累的影響[J]. 農業環境科學學報, 2018, 37(05): 41–48.
[11] 鄭君健, 劉杰, 張學洪, 等. 重金屬污染土壤植物修復及強化措施研究進展[J]. 廣東農業科學, 2013, 40(18): 159– 164.
[12] 祝方, 商執峰, 陳雨, 等. 螯合劑-大白菜修復電子垃圾拆解場地土壤中鋅與磷酸酶活性的研究[J]. 安全與環境工程, 2014, 21(3): 41–45.
[13] 袁菊紅, 胡綿好. EDDS處理對硒脅迫下彩葉草根系FTIR-ATR、SEM-EDXS特征及生理特性的影響[J]. 植物科學學報, 2014, 32(6): 620–629.
[14] 蘧苗苗, 陳銀萍, 蘇向楠, 等. 鎘脅迫下紫花苜蓿幼苗內源一氧化氮和活性氧的生成[J]. 廣西植物, 2016, 36(12): 1483–1491.
[15] HU Yan, LU Lingli,TIAN Shengke,et al. Cadmium-induced nitric oxide burst enhances Cd tolerance at early stage in roots of a hyperaccumulatorpartially by altering glutathione metabolism[J]. Science of The Total Environment, 2019, 650: 2761–2770.
[16] 胡亞虎, 魏樹和, 周啟星, 等. 螯合劑在重金屬污染土壤植物修復中的應用研究進展[J]. 農業環境科學學報, 2010, 29(11): 2055–2063.
[17] HUANG Meiyu, AI Honglian, XU Xiaoxiang, et al. Nitric oxide alleviates toxicity of hexavalent chromium on tall fescue and improves performance of photosystem II [J]. Ecotoxicology and environmental safety, 2018, 164: 32–40.
[18] 陳立, 王丹, 龍嬋, 等. 三種螯合劑對芥菜修復鈾鎘復合污染土壤的影響[J]. 農業環境科學學報, 2018,37 (08): 146–153.
[19] 滕玉瑾, 李翠祥, 馬婷, 等. 鹽脅迫下小麥幼苗NO和胞間ROS的相互作用及其對元素含量的調節[J]. 植物生理學報, 2015, (11): 1837–1845.
[20] 夏海威, 施國新, 黃敏, 等. 一氧化氮對植物重金屬脅迫抗性的影響研究進展[J]. 生態學報, 2015, 35(10): 3139– 3147.
[21] 張敏, 姜春輝, 李曉云, 等. 外源NO對銅脅迫下番茄幼苗根系構型及其超微結構的影響[J]. 植物生理學報, 2012, 48(11): 1105–1112.
[22] CANTóN F R , SUáREZ M F , CáNOVAS F M. Molecular aspects of nitrogen mobilization and recycling in trees[J]. Photosynthesis Research, 2005, 83(2): 265–278.
[23] 王建, 竇巧惠, 于世欣, 等. 外源NO對Cu脅迫下番茄幼苗L-精氨酸代謝的影響[J]. 生態毒理學報, 2015, 10(3): 112–122.
[24] 武佳葉, 郝瑞芝, 曹媛, 等. NO對鉛脅迫下玉米根尖細胞的影響[J]. 中國細胞生物學學報, 2013, (4): 469–475.
[25] 周萬海, 師尚禮, 寇江濤. 一氧化氮對NaCl脅迫下苜蓿種子萌發的影響[J]. 核農學報, 2012, 26(4): 710–716.
[26] 王瑋, 趙方貴, 侯麗霞, 等. NO參與AM真菌與煙草共生過程[J]. 生態學報, 2013, 33(23): 7583–7589.
[27] 張洪艷, 王文霞, 尹恒, 等. 殼寡糖誘導植物防御反應中一氧化氮信號的研究[J]. 中國生物工程雜志, 2011, 31(2): 18–22.
[28] 楊建軍, 張國斌, 郁繼華, 等. 鹽脅迫下內源NO對黃瓜幼苗活性氧代謝和光合特性的影響[J]. 中國農業科學, 2017(19).
[29] WOJTASZEK P . Nitric oxide in plants - To NO or not to NO[J]. Phytochemistry, 2010, 31(33): 1–4.
[30] RíOLA DEL, F JAVIER C, BARROSO J B. Nitric oxide and nitric oxide synthase activity in plants[J]. Phytoche-mistry, 2004, 65(7): 783–792.
[31] DOBRIKOVA A G , VLADKOVA R S , RASHKOV G D , et al. Effects of exogenous 24-epibrassinolide on the photosyn-thetic membranes under non-stress conditions[J]. Plant Physiology and Biochemistry, 2014, 80: 75–82.
[32] STIHR C. Generation and possible roles of NO in plant roots and their apoplastic space[J]. Journal of Experimental Botany, 2003, 53(379): 2293–2303.
[33] 劉建新, 王金成, 劉秀麗. 鑭脅迫下燕麥幼苗精氨酸代謝對外源NO的響應[J]. 中國稀土學報, 2018, 36(02): 236–246.
[34] ZHAO Mingui, CHEN Lei, ZHANG Lili, et al. Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in[J]. Plant Physiology, 2009, 151(2): 755–767.
Effects of EDDS on NO production in the seedlings ofLunder Cd stress
DING Jungang, CHEN Yinping*, YANG Bo, SHI Xiaoping, XIE Haifan
School of Environmental and Municipal Engineering, Lanzhou Jiaotong University, Lanzhou 730070
An indoor pot experiment was conducted to study the effects of EDDS (0, 0.5, 1.5, 2.5, 5.0 mmol.L-1) and EDDS combined with nitric oxide (NO) donor sodium nitroprusside (SNP) (0, 0.25, 0.5, 1.0 mmol.L-1) on stress signal molecule(NO) and nitric oxide synthase (NOS) activity production ofL. seedlings under 40 mg·kg-1Cd (CdCl2.2.5 H2O) stress. The results showed that NO production in different parts ofseedlings increased with the increase of EDDS concentration, and reached the maximum at 5.0 mmol·L-1; 0.5 mmol·L-1EDDS enhanced nitric oxide synthase (NOS) activity in roots and leaves. Selecting EDDS treatment concentration with more NO production and significant increase in NOS activity to combined with SNP to treatseedlings showed that NO production in roots increased first and then decreased, but increased continuously in stems and leaves with the increase of SNP concentration. The NOS activity in the plant could be further enhanced by appropriate concentration of SNP.The NO induced by EDDS was inhibited by nitrate reductase (NR) inhibitor (NaN3) and NOS inhibitor (L-NAME), and NaN3and L-NAME had little effect on NOS activity. The NO scavenger (c-PTIO) effectively removed some of the NO and enhanced NOS activity in roots and leaves.Therefore, under the stress of Cd, the NO production inL. seedlings could be increased by single application of EDDS and combined application of SNP with suitable concentration.
EDDS;NO;L.;Cd stress;NOS
10.14108/j.cnki.1008-8873.2020.04.003
丁浚剛, 陳銀萍, 楊波, 等. EDDS對Cd脅迫下三葉鬼針草幼苗NO產生的影響[J]. 生態科學, 2020, 39(4): 19–25.
DING Jungang, CHEN Yinping, YANG Bo, et al. Effects of EDDS on NO production in the seedlings of Bidens pilosa L. under Cd stress[J]. Ecological Science, 2020, 39(4): 19–25.
Q494
A
1008-8873(2020)04-019-07
2019-10-07;
2019-12-27
國家自然科學基金項目(31560161;31260089;31640012)
丁浚剛(1992—), 男, 在讀碩士, 主要從事環境生態學研究, E-mail: 1937657901@qq.com
陳銀萍(1974—) , 女, 博士, 教授, 主要從事植物逆境生理生態學研究, E-mail: yinpch@mail.lzjtu.cn
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
