紅鈴蟲對Bt棉花的抗性現狀及抗性機制
2020-08-21 05:31:30王金濤叢勝波
湖北植保 2020年4期
王 玲 王金濤 叢勝波 許 冬 萬 鵬
(農業部華中作物有害生物綜合治理重點實驗室/農作物病蟲草害防控湖北省重點實驗室/湖北省農業科學院植保土肥研究所武漢 430064)
紅鈴蟲(Pectinophora gossypiella),屬鱗翅目Lepidoptera麥蛾科Gelechiidae,是一種世界性的重要棉花害蟲,以其幼蟲危害棉蕾和花鈴,導致棉鈴內部的纖維受損或者造成蕾鈴與棉籽脫落,使棉鈴重量減輕、纖維品質下降甚至棉籽破碎[1]。2000年之前,紅鈴蟲在我國長江流域棉區常年為害成災,致使棉花產量損失10%~30%,嚴重影響了該區域的棉花生產[2]。
蘇云金芽孢桿菌(Bacillus thuringiensis,Bt),是一種革蘭氏陽性菌,在產孢過程中產生具有殺蟲活性的晶體蛋白[3]。自轉Bt基因棉花商業化種植之后,為紅鈴蟲等鱗翅目害蟲的防治提供了新的有效的手段。隨著Bt棉花的不斷種植,紅鈴蟲的種群數量得到了有效的控制[4]。然而,由于紅鈴蟲是寡食性害蟲,在棉田面臨著持續的選擇壓力,對Bt棉花存在抗性演化的風險。事實上,世界范圍內已發現一些田間紅鈴蟲種群對Bt棉花產生了實質抗性,其抗性問題嚴重威脅著Bt棉花的持續使用。
1 紅鈴蟲對Bt棉花的抗性現狀
我國長江流域自2000年開始種植轉Bt(Cry1Ac)基因棉花,隨著Bt棉花的大面積推廣應用,有效的抑制了紅鈴蟲的發生與危害。然而,該區域并沒有要求種植一定比例的非Bt棉花作為“庇護所”,隨著Bt棉種植面積的增加,到2006年,非Bt棉花的種植比例首次下降到20%以下[4]。到2008~2009年,非Bt棉花的種植比例降至12%。直至2010年開始,F2代Bt棉花的種植比例大幅度上升,為紅鈴蟲提供了23%的非Bt棉庇護所,此后非Bt棉花的比例常年維持在25%左右[4]。常年的抗性監測發現,2008~2010年長江流域棉區紅鈴蟲對Cry1Ac的敏感性相對于2005~2007年有所下降,表現出早期預警[5];而2011~2015年該區域紅鈴蟲對Cry1Ac的敏感性相對于2008~2010又明顯上升。此外,通過檢測7種已知的抗性等位基因,發現2012~2015年這7種抗性等位基因的基因頻率均處于較低水平,7種等位基因的總頻率從2012年的0.0105(0.0084~0.0132)下 降 到 2015 年 的 0.0046(0.0031~0.0067),下降了2.3倍[6]。這說明“庇護所”對延緩紅鈴蟲對Bt棉花的抗性演化是十分重要的,2006~2009年期間由于非Bt棉的比例極低,導致早期預警的發生,但隨著2010年F2代Bt棉的大面積種植,提供了充足的非Bt棉庇護所,反轉了紅鈴蟲對Bt棉花的早期抗性,同時還減少了化學農藥的使用[4]。因此,到目前為止,我國長江流域紅鈴蟲對Bt棉花仍然保持著較好的敏感性(表1)。
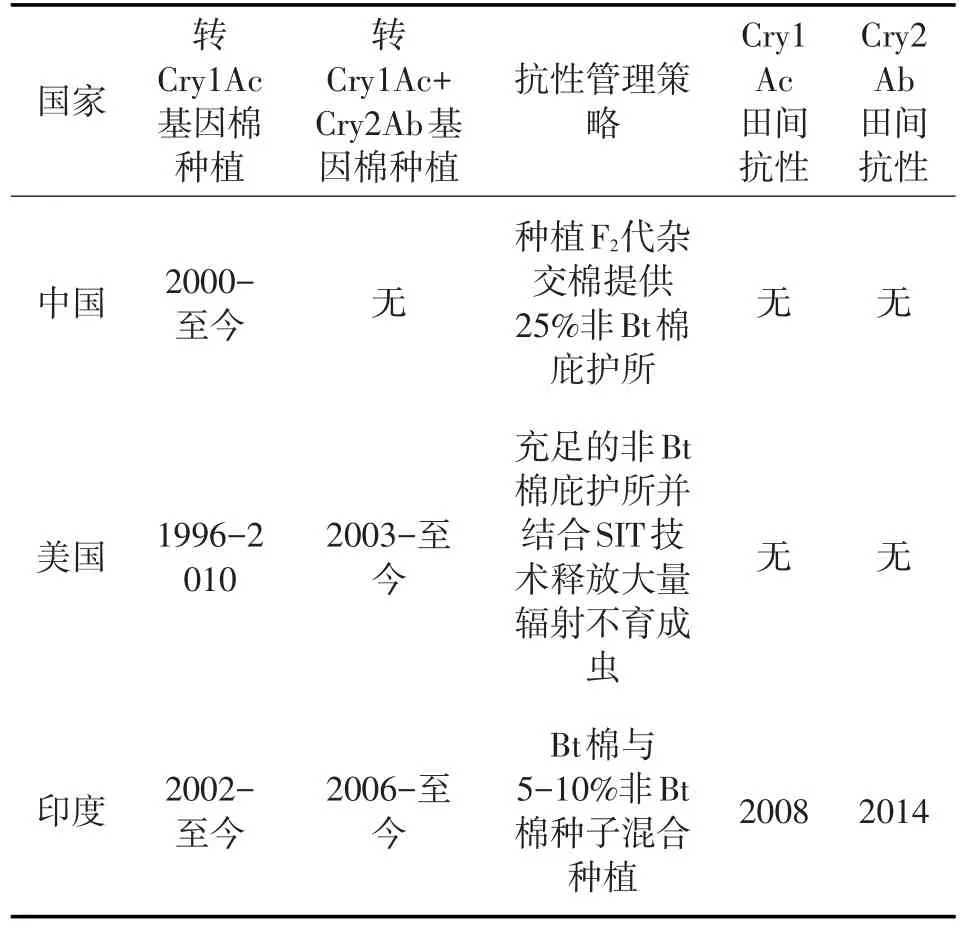
表1 三個國家Bt棉花的種植及紅鈴蟲田間抗性現狀
美國亞利桑那州在引進Bt棉花之前,紅鈴蟲的種群密度常年較高。自1996年開始種植轉Bt(Cry1Ac)基因棉花,該區域的棉花種植者一直遵循法定的“庇護所”策略,部分田塊用于種植非Bt棉花,使得該區域非Bt棉花的種植比例每年達25%以上[7]。到2005年,Bt棉花造成了紅鈴蟲種群密度的區域性下降和廣譜殺蟲劑噴灑量的減少[8]。2003年,美國開始逐漸從單一種植轉Bt(Cry1Ac)單價棉轉向同時種植單價棉和轉Bt(Cry1Ac+Cry2Ab)雙價棉。常年對亞利桑那州田間種群進行室內生物測定及抗性等位基因檢測,發現1997年~2005年,該區域的田間紅鈴蟲種群對Cry1Ac的敏感性并沒有下降(表1)[7]。從2006年開始,美國開始施行紅鈴蟲根除計劃,通過在田間釋放大量輻射不育的成蟲來延緩紅鈴蟲對 Bt棉花的抗性[7,9]。自此,亞利桑那州的紅鈴蟲種群數量急劇下降:田間非Bt棉上的紅鈴蟲幼蟲從2005年的15.3%下降到2009年的0.012%,下降了99.92%;而信息素誘集的成蟲數量從2005年的每周26.7只下降至2009年的0.0054只,下降了99.98%[9]。2010年~2018年,亞利桑那州田間棉鈴上未發現紅鈴蟲幼蟲;2013年~2018年,田間未發現野生的紅鈴蟲成蟲[10]。其他州的情況與亞利桑那州類似,最終美國農業部于2018年10月宣布紅鈴蟲已在美國被根除[10]。
印度自2002年開始種植轉Bt(Cry1Ac)基因棉,2006年開始引進轉Bt(Cry1Ac+Cry2Ab,Bollgard II)雙價棉。雖然印度基因工程批準委員會(GEAC)要求種植20%非Bt棉花作為庇護所以延緩抗性發展,但印度農戶并沒有嚴格執行。2014年,印度轉Bt棉花的種植面積已達棉花種植總面積的 95%,而 Bt棉中 96%為轉 Bt(Cry1Ac+Cry2Ab)雙價棉,而轉Bt(Cry1Ac)單價棉僅占4%[11](Choudhary和Gaur,2015年)。與美國和中國對Bt棉花的持續敏感性不同,在印度,由于非Bt棉庇護所非常稀少,紅鈴蟲迅速發展出對Bt棉花的實際抗性[12,13]。2008年,首次記錄了印度古吉拉特邦田間紅鈴蟲種群對Cry1Ac產生抗性(表1)[14]。2014年,來自印度中部的8個田間種群對Cry1Ac和Cry2Ab的平均抗性倍數分別為310和78,而且轉Bt(Cry1Ac+Cry2Ab)雙價棉棉鈴上的幼蟲侵染率(52%)高于非Bt棉上的幼蟲侵染率(27%)[13]。到 2015年,紅鈴蟲對轉Bt(Cry1Ac+Cry2Ab)雙價棉產生抗性在印度棉區已經普遍存在[13]。
2 紅鈴蟲對Bt的抗性機制
紅鈴蟲對Bt的抗性機制目前可以歸為兩類,一種是對Cry1Ac的抗性,目前已報道的所有室內或者田間紅鈴蟲Cry1Ac抗性品系或者抗性個體均有鈣粘蛋白基因(PgCad1)介導,尚未發現其他基因介導紅鈴蟲對Cry1Ac的抗性。總共報道了17種不同的PgCad1抗性等位基因[15-22],這些抗性等位基因全部涉及PgCad1的突變,只有r17還涉及了PgCad1的表達量下調(表2)。在這17種抗性等位基因中,有9種抗性等位基因(r5、r7-r12、r15、和r17)發生了可變剪切或錯誤剪切,產生2~4種不同的轉錄本[18,21,22];說明可變剪切或錯誤剪切在紅鈴蟲PgCad1抗性等位基因中是普遍存在的。抗性等位基因r3、r15和r16均發生了轉座子的插入,其中r3等位基因在第21個外顯子插入一個4739 bp的非LTR反轉錄轉座子[23],r16等位基因在第20個外顯子插入一個1545 bp的退化轉座子[20],r15等位基因最為特殊,在第28個外顯子插入了一個3370 bp的片段,該插入片段是一種MITE轉座子,且MITE轉座子中包含了另外2次轉座子插入事件,插入了SINE與RTE這2種轉座子[21],這說明轉座子插入也是導致紅鈴蟲PgCad1突變的重要因素。所有的PgCad1抗性等位基因均編碼產生結構不完整的鈣粘蛋白,涉及除胞內區之外的各個區域,包括信號肽、鈣粘蛋白重復區域、近膜區以及跨膜區的變異,以鈣粘蛋白重復區域的變異為主,這些結構不完整的鈣粘蛋白與Cry1Ac的結合能力下降或者定位錯誤,從而導致紅鈴蟲對Cry1Ac產生抗性[19-21]。此外,抗性等位基因r17的轉錄水平相對于敏感型PgCad1顯著下調79~190倍,這也是導致紅鈴蟲對Cry1Ac產生抗性的重要原因[22]。
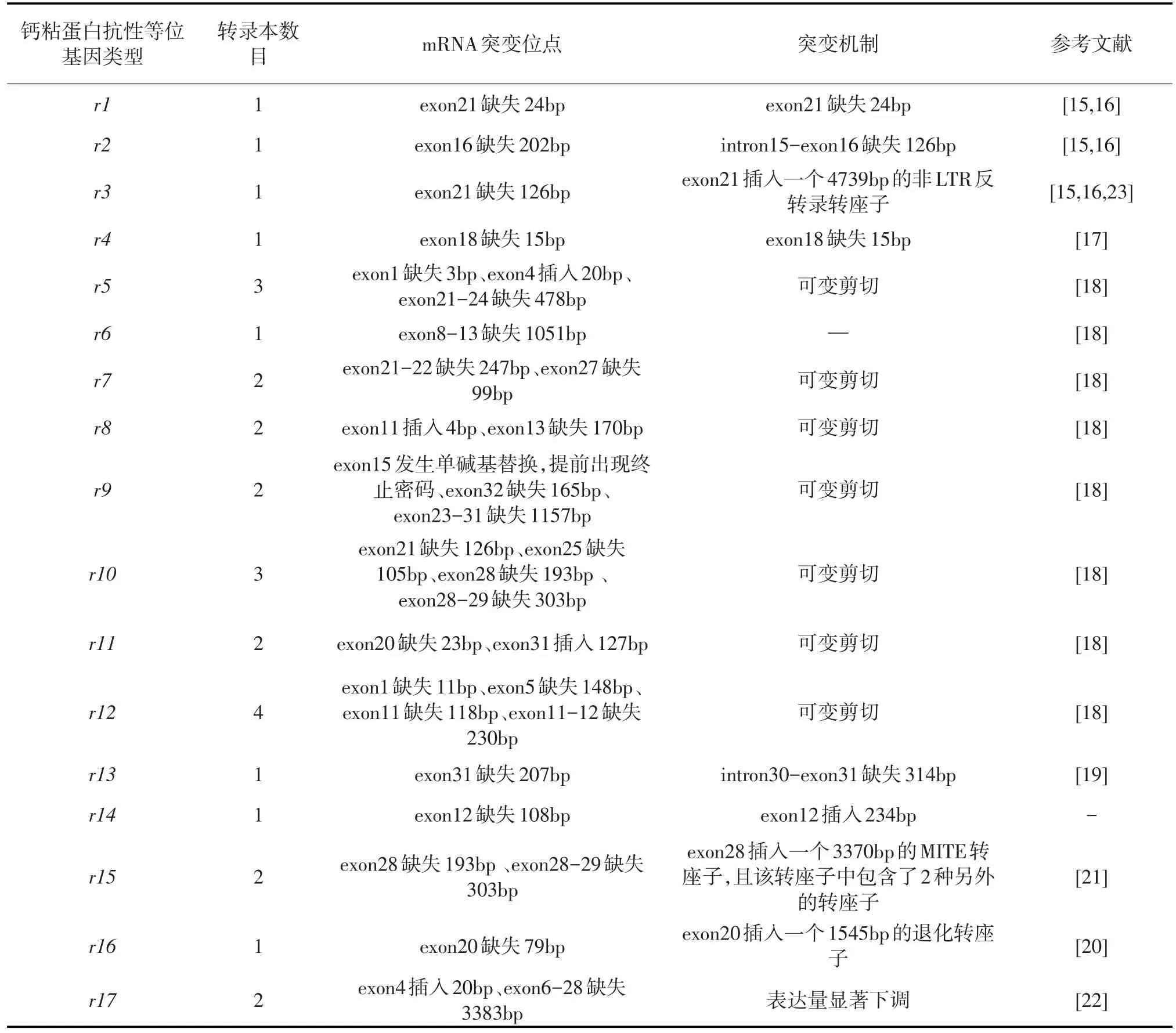
表2 紅鈴蟲鈣粘蛋白抗性等位基因及其介導Cry1Ac抗性的機制
另一種是對Cry2Ab的抗性,與ABC轉運蛋白ABCA2基因(PgABCA2)的變異相關。美國室內篩選的Cry2Ab抗性品系及印度田間的Cry2Ab抗性個體,均有由PgABCA2錯誤剪切導致[24]。完整的內含子被保留、5′或3′的剪切位點被替換以及外顯子跳躍是導致PgABCA2發生錯誤剪切的主要原因,目前以及發現20種以上不同的剪切形式,主要涉及外顯子4、6、8、14與20,其中錯誤剪切導致外顯子6缺失在室內抗性品系與田間抗性個體中均有發生[24]。值得一提的是,在紅鈴蟲中,Cry1Ac和Cry2Ab之間的交叉抗性是不對稱的,PgCad1突變的Cry1Ac抗性品系對Cry2Ab幾乎沒有交互抗性[19-21],而Cry2Ab抗性品系卻對Cry1Ac存在交互抗性[25]。
3 展望
在中國、美國和印度,紅鈴蟲對Bt棉花的抗性演化存在驚人的差異,這說明合理有效的抗性治理策略對延緩害蟲的抗性發展至關重要。制定合理有效的抗性治理策略,依賴于對Bt與害蟲互作機制的掌握。就紅鈴蟲而言,目前無論是室內的抗性品系,還是田間的抗性個體,都由PgCad1的不同突變形式介導Cry1Ac抗性,這說明PgCad1在Cry1Ac對紅鈴蟲的作用過程中是一個極為敏感的步驟,容易發生變異而導致抗性的發生。雖然來自三個國家的紅鈴蟲Cry1Ac抗性品系或個體都與PgCad1突變相關,但PgCad1突變的形式并不完全相同,這可能是由于三個國家Bt棉花的種植模式不同,紅鈴蟲受到的篩選壓力并不相同,從而導致田間種群的抗性演化有所差異。此外,PgCad1與PgABCA2的突變形式如此多樣化,也說明了田間抗性的發展是極其復雜的,田間很可能還存在更多的突變形式;也可能存在其他抗性基因介導紅鈴蟲對Cry1Ac或者Cry2Ab的抗性甚至介導二者的不對稱交互抗性,只有進一步深入了解這些抗性基因的類型及其對Bt抗性的貢獻大小與機制,才能有針對性的制定合理的抗性治理策略以繼續延緩紅鈴蟲對Bt棉花抗性的發展。
