普通小麥-濱麥及其衍生系的染色體組成分析
2020-07-31 03:35:54楊曉菲王長(zhǎng)有陳春環(huán)田增榮吉萬全
麥類作物學(xué)報(bào) 2020年3期
楊曉菲,王長(zhǎng)有,陳春環(huán),田增榮,吉萬全
(1.渭南師范學(xué)院環(huán)境與生命科學(xué)學(xué)院,陜西渭南 714099; 2.旱區(qū)作物逆境生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室/西北農(nóng)林科技大學(xué)農(nóng)學(xué)院,陜西楊凌 712100)
賴草屬(LeymusHochst.)是一個(gè)從四倍體至十二倍體類型均有的多倍體物種,其中許多物種[如大賴草(L.racemusus)、羊草(L.chinensis)、多枝賴草(L.multicaulis)等]的染色體核型已經(jīng)非常清楚[1-2],而濱麥(Leymusmollis)是賴草屬中的一個(gè)四倍體物種,具有抗寒、抗旱、耐鹽堿[3]、莖稈粗壯和大穗[4]等優(yōu)良性狀,同時(shí)對(duì)小麥條銹病、葉銹病和白粉病等多種真菌病害免疫[5-6]。作為小麥改良育種的重要三級(jí)基因資源[7],其染色體核型的研究處于空白狀態(tài),且四倍體賴草屬植物基因組(染色體組)構(gòu)成目前尚未定論。濱麥基因組為NsNsXmXm,其中Ns基因組來自于新麥草(PsathyrostachysKeng)[8-10],而Xm基因組的來源目前尚未確定。在第2屆國(guó)際小麥族研討會(huì)上(1994年美國(guó)猶他州),基因組命名委員會(huì)建議使用NsXm作為四倍體賴草屬植物基因組的表示符號(hào),Xm代表一個(gè)未知的基因組直到試驗(yàn)驗(yàn)證為止[11]。
染色體組型分析可以初步判斷外源染色體是否存在和具體數(shù)目,是植物分類和遺傳研究的重要方法之一。本研究對(duì)濱麥染色體核型進(jìn)行初步分析并繪制濱麥染色體核型模式圖,同時(shí)通過分子標(biāo)記研究其可能的二倍體供體種并分析其與濱麥之間的親緣關(guān)系。
Pedersen等[12]利用FISH技術(shù)結(jié)合重復(fù)序列pAsl和pHvG38,分辨出中國(guó)春和綿陽(yáng)11中的21對(duì)染色體。由于重復(fù)序列探針的準(zhǔn)備耗時(shí)費(fèi)力,部分寡核苷酸探針的發(fā)展為辨別染色體提供了一種更經(jīng)濟(jì)高效的方法。Tang等[13]研究認(rèn)為,兩種寡核苷酸探針pSc119.2和pTa-535代替重復(fù)序列pAsl和pHvG38做雙色FISH,同樣可以清晰準(zhǔn)確地識(shí)別小麥21對(duì)染色體。因此,本研究擬結(jié)合兩種寡核苷酸探針對(duì)普通小麥7182的標(biāo)準(zhǔn)核型進(jìn)行繪制,并利用FISH-GISH技術(shù)對(duì)普通小麥-濱麥高世代衍生系的染色體組成進(jìn)行分析,旨在為后續(xù)創(chuàng)制和鑒定出更多類型的普通小麥-濱麥種質(zhì)資源奠定基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料
研究材料包括普通小麥7182(Triticumaestivum,2n=6x=42,AABBDD),濱麥(L.mollis,2n=4x=28,NsNsXmXm),華山新麥草(Psathyrostachyshuashanica,2n=2x=14,NsNs),二倍體長(zhǎng)穗偃麥草(Thinopyrumelongatum,2n=2x=14,EeEe),擬鵝觀草(Pseudoroegeriaspicata,2n=2x=14,StSt)和百薩偃麥草(Thinopyrumbessarabicum,2n=2x=14,EbEb)以及普通小麥-濱麥高世代衍生系M47和M39。
1.2 試驗(yàn)方法
1.2.1 根尖培養(yǎng)與取材
將供試材料種子腹溝朝下,置于鋪有兩層濾紙的干凈培養(yǎng)皿中,加ddH2O濕潤(rùn)濾紙即可(無多余的水分流動(dòng)為宜)。將培養(yǎng)皿置于23~25 ℃恒溫培養(yǎng)箱中黑暗培養(yǎng),待根長(zhǎng)至3~5 cm時(shí),剪取長(zhǎng)約1 cm根尖放入濕潤(rùn)且打孔的普通離心管(1.5 mL)中并編號(hào)記錄,放入笑氣(N2O)罐中密封處理2 h,之后在離心管中加入300 μL 90% 冰乙酸,冰浴固定10 min后,ddH2O沖洗根尖3次,靜置,待離心管完全干燥后,加入70% 冰乙醇,置于-20 ℃冰箱保存?zhèn)溆谩?/p>
濱麥根尖于2015-2017連續(xù)三年在3月下旬至4月上旬取材兩到三次,挖取新生根并剪取約1 cm長(zhǎng)的根尖,置于冰水混合物中處理22~26 h,然后用卡諾氏固定液(無水乙醇∶冰乙酸=3∶1,v/v)于4 ℃冰箱固定48 h左右,用ddH2O將根尖沖洗至少3次,隨后加入70%冰乙醇,-20 ℃冰箱保存?zhèn)溆谩?/p>
1.2.2 根尖處理與制片
用ddH2O沖洗70%冰乙醇保存的根尖,用濾紙吸干水分,切取乳白色根尖部分置于裝有2% 纖維素酶和1%果膠酶混合酶液的0.5 mL離心管中,瞬時(shí)離心,待乳白色根尖沉入離心管底部,將離心管放入37 ℃水浴酶解55~60 min(不同材料酶解時(shí)間有所不同)。然后用70% 乙醇沖洗三次,用解剖針將浸潤(rùn)在100 μL 70% 乙醇中的根尖搗碎,6 000 r·min-1離心1~2 min,倒置離心管于濾紙上吸干水分,加40 μL預(yù)冷的純乙酸,高速瞬時(shí)渦旋3次,移液槍吸取10 μL懸濁液滴于干凈載玻片上(處于黑暗且濕潤(rùn)的盒子),5 min后待懸濁液緩慢均勻擴(kuò)散后于顯微鏡下鏡檢并拍照。
1.2.3 染色體核型分析與模式圖繪制
根據(jù)李懋學(xué)等[14]關(guān)于植物核型的分析標(biāo)準(zhǔn),選取30個(gè)以上染色體形態(tài)和分散較好的中期分裂相細(xì)胞進(jìn)行染色體數(shù)目的統(tǒng)計(jì),精選五個(gè)細(xì)胞進(jìn)行拍照。利用圖象處理軟件Adobe Photoshop CS6進(jìn)行細(xì)胞背景處理,測(cè)量各個(gè)染色體及各臂的相對(duì)長(zhǎng)度,進(jìn)行染色體的配對(duì),排列建立核型圖。
利用Excel對(duì)染色體的各種參數(shù)進(jìn)行數(shù)據(jù)分析,并繪制出整套染色體的核型模式圖。按照Stebbins[15]的核型標(biāo)準(zhǔn)進(jìn)行核型分類。
1.2.4 SSR和EST分子標(biāo)記分析
分子標(biāo)記分析共選用825對(duì)SSR引物和86對(duì)EST引物,SSR和EST引物序列均來自GrainGenes數(shù)據(jù)庫(kù)(http://wheat.pw.usda.gov/GG2/index.shtml),由奧科鼎盛生物科技有限公司(北京)合成。采用改良后的CTAB法對(duì)供試材料進(jìn)行總基因組DNA的提取。PCR反應(yīng)體系為10 μL,包括1.0 μL PCR Buffer 緩沖液(含Mg2+)、1.0 μL 0.5 μmol·L-1primers(正、反向引物各0.5 μL)、0.8 μL 0.2 mmol·L-1dNTPs混合液、模板DNA 1.0 μL(200 ng·μL-1)、0.1 μL Taq DNA 聚合酶(5 U·μL-1),ddH2O補(bǔ)充至 10.0 μL。反應(yīng)在熱循環(huán)儀Bio-Rad S1000TM型(California,USA)上進(jìn)行。PCR產(chǎn)物用8%非變性聚丙烯酰胺凝膠檢測(cè),電泳步驟參照吳金華等[16]的方法,凝膠經(jīng)硝酸銀染色顯影后置于燈箱上觀察、照相并統(tǒng)計(jì)帶型。
1.2.5 雙色FISH-GISH技術(shù)
參照Tang 等[13]的方法,利用英駿(Invitrogen)生物技術(shù)有限公司(上海)合成的兩種寡核苷酸探針Oligo-pTa535(Tamra-red)、Oligo-pSc119.2(FAM-green)序列,以及濱麥葉片總基因組DNA為探針,先后對(duì)根尖體細(xì)胞進(jìn)行熒光原位雜交(FISH)和基因組原位雜交(GISH)分析。濱麥葉片gDNA的提取參照改良后的CTAB法[17]進(jìn)行。使用OLYMPUS BX53(日本)熒光顯微鏡鏡檢,利用系統(tǒng)軟件DP800 CCD、cellSens Standaed 1.8進(jìn)行拍照并用圖像處理軟件Adobe Photoshop CS6分析圖像。
2 結(jié)果與分析
2.1 濱麥染色體核型分析及核型模式圖
鏡檢結(jié)果表明,濱麥根尖的細(xì)胞染色體數(shù)目為28條,即2n=4x=28(圖1a)。利用圖像處理軟件Adobe Photoshop CS6對(duì)染色體及各臂的相對(duì)長(zhǎng)度進(jìn)行測(cè)量,根據(jù)測(cè)量結(jié)果進(jìn)行配對(duì)和排序,繪制濱麥染色體核型圖(圖1b)。計(jì)算染色體長(zhǎng)臂與短臂相對(duì)長(zhǎng)度的比值并確定每一對(duì)染色體所屬類型(表1),繪制出濱麥染色體核型模式圖(圖1c)。依據(jù)核型標(biāo)準(zhǔn)推測(cè)出濱麥的核型公式為:2n=4x=28=22m(6sat)+6sm,其中m、sm和sat分別代表中間著絲粒染色體、亞中間著絲粒染色體和具有隨體的染色體。
2.2 濱麥染色體核型的歸屬
根據(jù)濱麥各對(duì)染色體的臂比(表1,圖1c),發(fā)現(xiàn)中間著絲粒染色體和亞中間著絲粒染色體分別有11對(duì)和3對(duì)。依據(jù)Stebbins[15]的染色體核型標(biāo)準(zhǔn)分類方案,即最長(zhǎng)與最短染色體之間的長(zhǎng)度比值<2記為A級(jí),比值在2~4之間記為B級(jí),比值>4記為C級(jí);臂比(長(zhǎng)臂/短臂)>2的比例為0記為I級(jí),0.01~0.5記為II級(jí),0.51~0.99記為Ⅲ級(jí),1.0記為Ⅳ級(jí)。本研究中濱麥最長(zhǎng)和最短染色體的比值為1.98,記為A級(jí);染色體臂比>2的比例為0.07,記為II級(jí)。因此,濱麥染色體的核型類型為IIA。
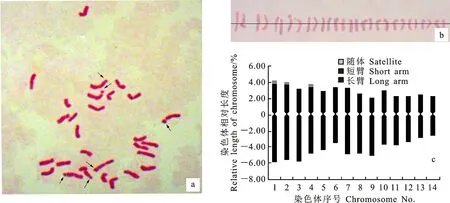
箭頭指示濱麥染色體的隨體。
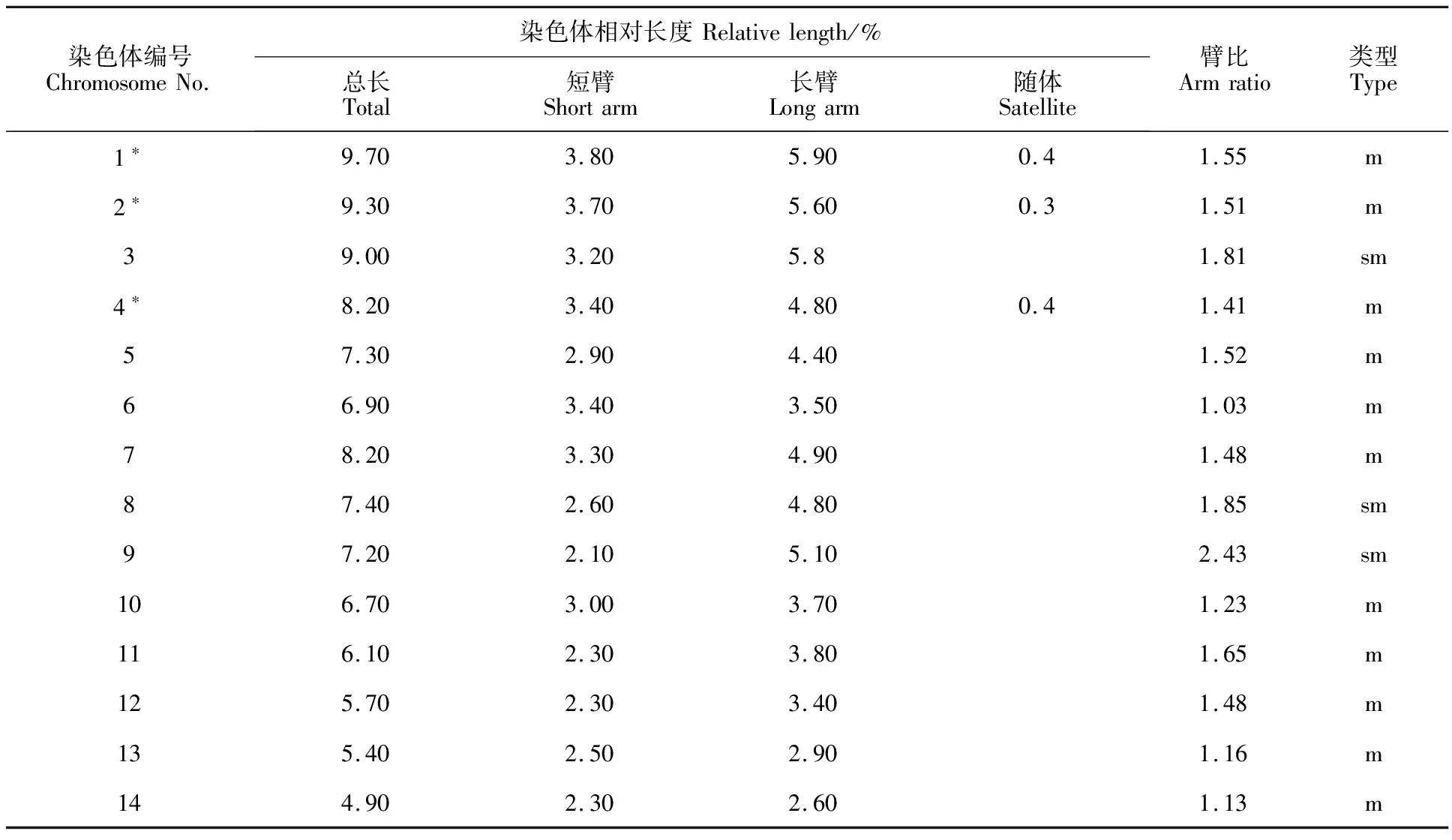
表1 濱麥染色體的相對(duì)長(zhǎng)度(總長(zhǎng)、長(zhǎng)短臂)、臂比及類型
2.3 濱麥染色體組供體種的初步分析
分子標(biāo)記擴(kuò)增結(jié)果表明,456對(duì)標(biāo)記(包括379對(duì)SSR引物和77對(duì)EST引物)在二倍體華山新麥草、二倍體長(zhǎng)穗偃麥草、百薩偃麥草、擬鵝觀草和四倍體濱麥中表現(xiàn)多態(tài)性(部分結(jié)果如圖2),58對(duì)標(biāo)記在濱麥和四種二倍體物種中擴(kuò)增帶型完全一致,其余311對(duì)標(biāo)記在濱麥、華山新麥草、二倍體長(zhǎng)穗偃麥草、擬鵝觀草和百薩偃麥草中無擴(kuò)增條帶或者擴(kuò)增條帶模糊。濱麥染色體組供體種分子標(biāo)記的初步分析結(jié)果表明,在多態(tài)性標(biāo)記中有157對(duì)SSR標(biāo)記和27對(duì)EST標(biāo)記在華山新麥草和濱麥間擴(kuò)增出相同帶型;有52對(duì)SSR標(biāo)記和15對(duì)EST標(biāo)記在百薩偃麥草和濱麥間擴(kuò)增出相同帶型;有62對(duì)SSR標(biāo)記和21對(duì)EST標(biāo)記在二倍體長(zhǎng)穗偃麥草和濱麥間擴(kuò)增出相同帶型;有67對(duì)SSR標(biāo)記和24對(duì)EST標(biāo)記在擬鵝觀草和濱麥間擴(kuò)增出相同帶型。這說明華山新麥草、百薩偃麥草、二倍體長(zhǎng)穗偃麥草和擬鵝觀草與濱麥擴(kuò)增出相同帶型的標(biāo)記分別占多態(tài)性標(biāo)記數(shù)的40.35%、14.69%、18.20% 和19.96%。
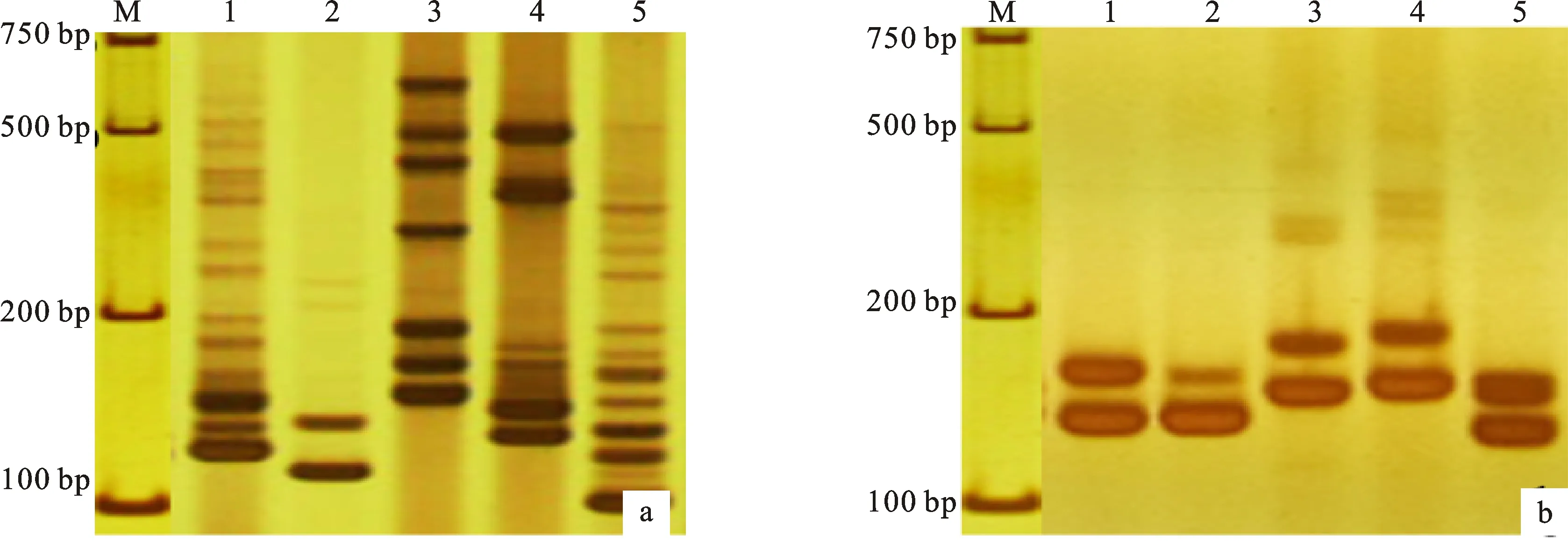
a:Xgpw7574-5B標(biāo)記的PCR結(jié)果;b:Xgwm674-3A標(biāo)記的PCR結(jié)果;M:Marker;1:濱麥;2:華山新麥草;3:百薩偃麥草;4:二倍體長(zhǎng)穗偃麥草;5:擬鵝觀草。
2.4 普通小麥7182的FISH標(biāo)準(zhǔn)核型圖
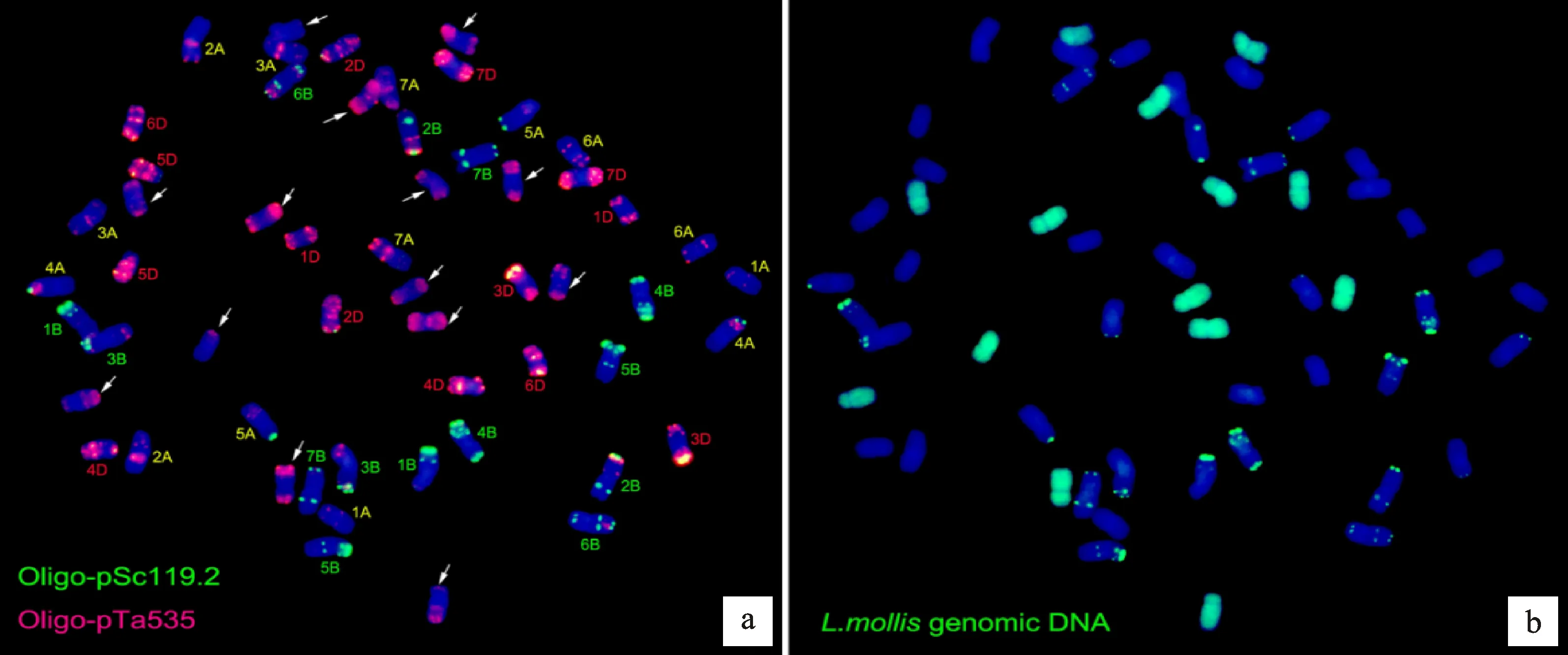
傅 杰等[18]通過組織培養(yǎng)獲得普通小麥-濱麥的雜種F1植株,經(jīng)秋水仙素處理后與普通小麥品系7182回交獲得了雜種后代。本研究結(jié)合兩種寡核苷酸探針pSc119.2和pTa-535,采用雙色FISH技術(shù)繪制出普通小麥7182所有染色體的FISH標(biāo)準(zhǔn)核型圖(圖3)。
紅色:Oligo-pTa535探針;綠色:Oligo-pSc119.2探針;藍(lán)色:DPAI復(fù)染。
2.5 普通小麥-濱麥高世代衍生系M47和M39的染色體組成
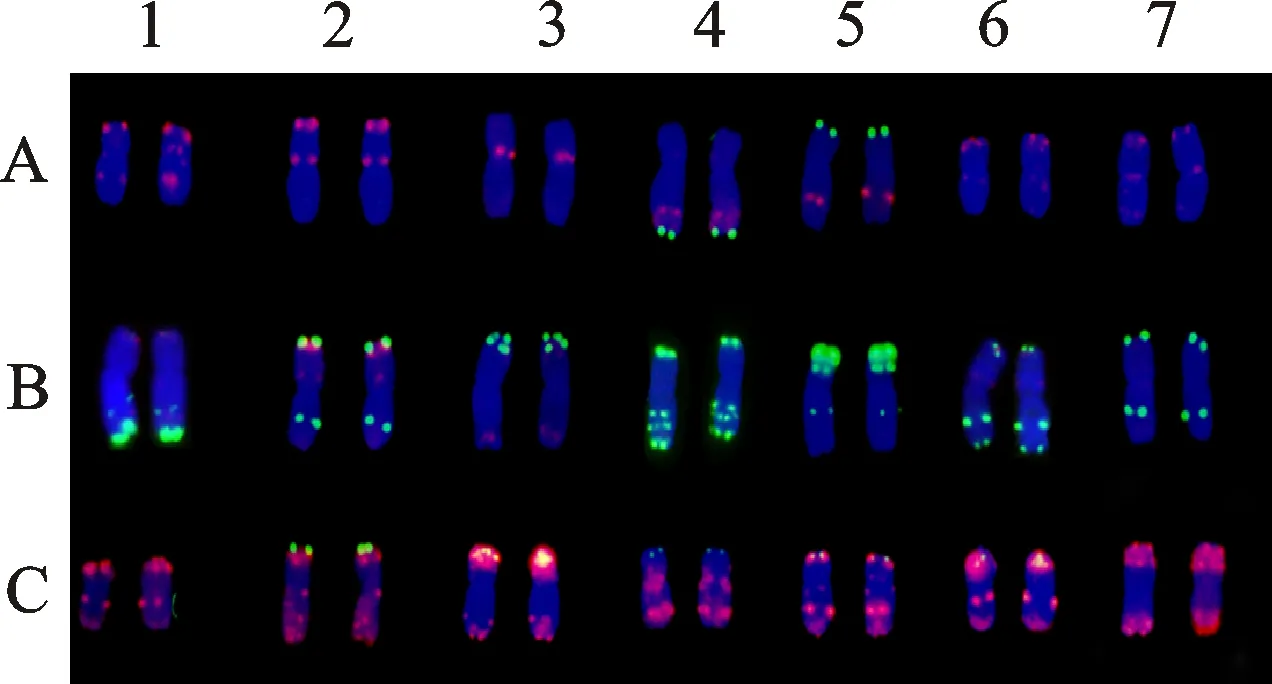
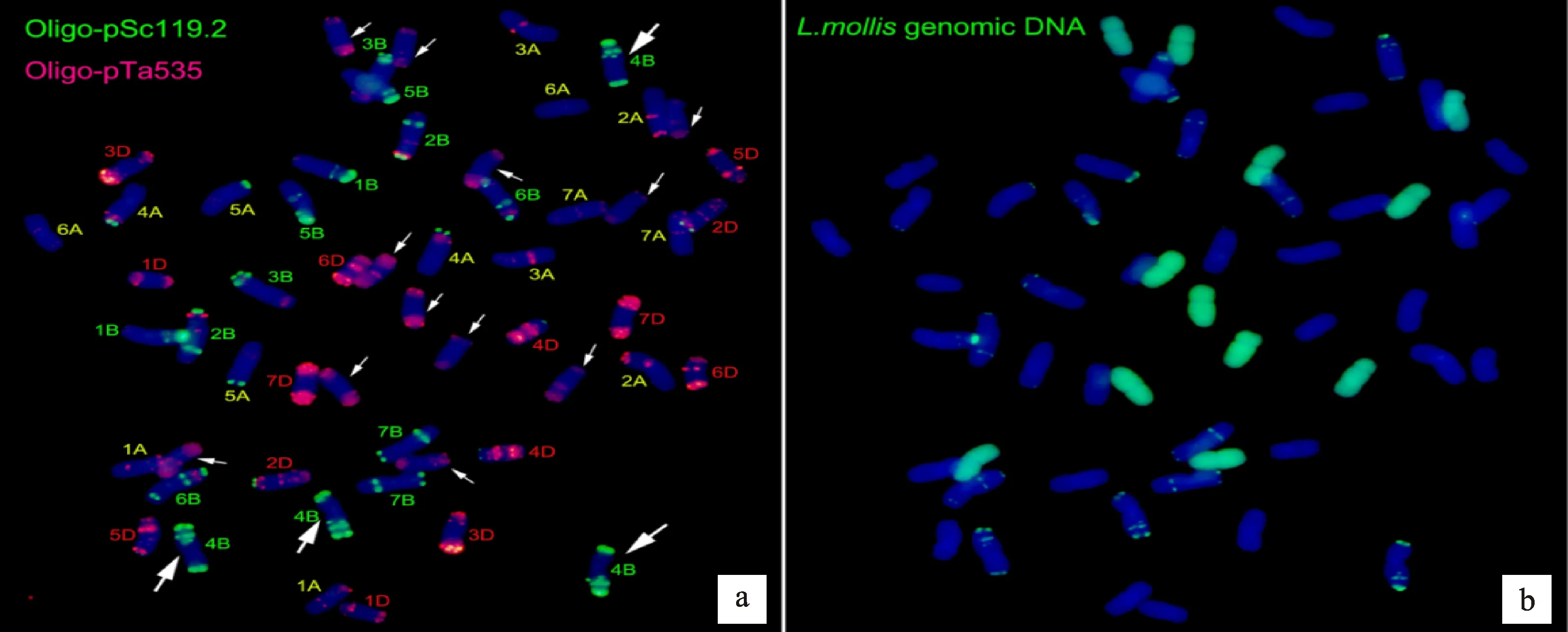
利用寡核苷酸探針pSc119.2和pTa-535對(duì)高世代衍生系M47和M39進(jìn)行順序雙色FISH-GISH鑒定。細(xì)胞學(xué)觀察結(jié)果表明,M47和M39體細(xì)胞染色體數(shù)目均為56條,不同的是,M47中有42條普通小麥染色體,14條雙末端紅色信號(hào)染色體(圖4a);而M39中有44條普通小麥染色體,其中包含4條4B小麥染色體,僅有12條雙末端紅色信號(hào)染色體(圖5a),因此,初步推斷雙末端紅色信號(hào)染色體為濱麥染色體。為了進(jìn)一步確認(rèn)雙末端紅色染色體的來源,利用濱麥的總基因組DNA為探針進(jìn)行GISH鑒定,結(jié)果表明,雙末端紅色信號(hào)染色體為濱麥染色體(圖4b、5b)。
a:利用寡核苷酸探針Oligo-pTa535(紅色)和Oligo-pSc119.2(綠色)對(duì)M47的FISH分析,顯示42條小麥染色體(已標(biāo)出)及14條雙末端紅色信號(hào)染色體(白色箭頭所指);b:利用濱麥總基因組DNA 為探針(綠色)在同一張片子上對(duì)M47的GISH分析。以DAPI為復(fù)染劑(藍(lán)色)。
a:利用寡核苷酸探針Oligo-pTa535(紅色)和Oligo-pSc119.2(綠色)對(duì)M39的FISH分析,顯示44條普通小麥染色體(已標(biāo)出)包含4條4B染色體(白色大箭頭所指)及12條雙末端紅色信號(hào)染色體(白色小箭頭所指);b:利用濱麥總基因組DNA 為探針(綠色)在同一張片子上對(duì)M39的GISH分析。DAPI為復(fù)染劑(藍(lán)色)。
3 討 論
3.1 濱麥核型及染色體組構(gòu)成分析
智 力[19]于2000年報(bào)道了窄穎賴草(L.angustus)、粗穗賴草(L.crassiuculus)、若羌賴草(L.ruoqiangensis)、大賴草(L.ruoqiangensis)和羊草(L.chinensis)等5個(gè)不同賴草屬物種的染色體核型;楊瑞武等[1]于2004年報(bào)道了十一個(gè)賴草屬四倍體物種,包括多枝賴草(L.multicaulis)、分枝賴草(L.ramosus)、沙生賴草(L.arenarius)、鹽生賴草(L.salinus)、密穗賴草(L.condensatus)、新生賴草(L.innovatus)、灰賴草(L.cinereus)、柴達(dá)木賴草(L.pseudoracemosus)、無芒賴草(L.triticoides)、阿克摩林賴草(L.akmolinensis)及賴草(L.secalinus)的染色體核型。分析比較得知,上述賴草屬不同物種均具有隨體染色體,且多數(shù)為中部著絲粒染色體,每個(gè)物種具有數(shù)量不等的近中部著絲粒染色體,其中個(gè)別物種還有少數(shù)的近端部著絲粒染色體。從核型的分類類型來看,除多枝賴草和沙生賴草核型為IIB外,其余物種的核型均為IIA。而葛榮朝等[20]發(fā)現(xiàn),多枝賴草染色體核型中未出現(xiàn)隨體染色體,這與楊瑞武[1]的研究結(jié)果不一致,造成結(jié)果差異的原因可能與試驗(yàn)材料來源和所采用的試驗(yàn)方法不同以及試驗(yàn)操作過程中的誤差等有關(guān)。
本研究對(duì)四倍體濱麥進(jìn)行了核型分析,核型公式為2n=4x=28=22m(6sat)+6sm,核型類型為ⅡA,可以得出具有隨體的濱麥染色體以中部著絲粒染色體為主,有少數(shù)的近中部著絲粒染色體,這與前人報(bào)道的賴草屬大多數(shù)物種的核型相似。綜合分析,賴草屬不同物種的染色體核型既有相似之處,也有各自的特征差異,這種差異可以作為物種基礎(chǔ)分類的細(xì)胞學(xué)依據(jù)。多態(tài)性分子標(biāo)記在華山新麥草、百薩偃麥草、二倍體長(zhǎng)穗偃麥草和擬鵝觀草與濱麥擴(kuò)增出相同帶型的不同比例初步表明,華山新麥草與濱麥的親緣關(guān)系較其他三個(gè)供體種更近。
3.2 普通小麥7128標(biāo)準(zhǔn)核型及其與濱麥高世代衍生系的染色體組成
本研究利用兩種寡核苷酸探針pSc119.2和pTa-535代替耗時(shí)費(fèi)力的重復(fù)序列pAsl和pHvG38,更加經(jīng)濟(jì)、高效和準(zhǔn)確地繪制出普通小麥7182中21對(duì)染色體的標(biāo)準(zhǔn)FISH核型圖。通過雙色FISH-GISH技術(shù)鑒定出兩種類型的普通小麥-濱麥高世代衍生系M47(2n=56=42T.a+14L.m)和M39(2n=56=44T.a+12L.m),均可作為創(chuàng)制新的普通小麥-濱麥種質(zhì)資源的中間橋梁材料。本研究為后續(xù)普通小麥-濱麥種質(zhì)資源的創(chuàng)制和鑒定提供了基礎(chǔ)理論依據(jù)和技術(shù) 方法。
