高溫脅迫對小麥花藥活性氧代謝的影響
2020-07-31 04:14:40盧奕霏顧迎港侯澤豪張迎新方正武馬東方劉易科朱展望張改生王書平
麥類作物學(xué)報 2020年4期
盧奕霏,顧迎港,陳 威,王 娜,康 珍,侯澤豪,張迎新,方正武,馬東方,劉易科,朱展望,張改生,王書平,4
(1.長江大學(xué)農(nóng)學(xué)院/主要糧食作物產(chǎn)業(yè)化湖北省協(xié)同創(chuàng)新中心,湖北荊州 434025; 2.湖北省農(nóng)業(yè)科學(xué)院糧食作物研究所,湖北武漢 4300643; 3.西北農(nóng)林科技大學(xué)農(nóng)學(xué)院,陜西楊凌 712100; 4.湖北荃銀高科種業(yè)有限公司,湖北荊州 434025)
隨著大氣中CO2和其他溫室氣體排放量的逐年升高,地球表面平均溫度也在日益增高,溫室效應(yīng)給全球農(nóng)業(yè)生產(chǎn)帶來極大危害[1-2]。研究表明,氣溫升高1 ℃以上時,小麥產(chǎn)量開始快速下降,升高3 ℃時,產(chǎn)量降低20%左右[3-4]。在小麥對逆境響應(yīng)機(jī)制的研究中,有關(guān)干旱、鹽脅迫和低溫脅迫的研究較多,而關(guān)于高溫脅迫對小麥影響的研究仍不全面系統(tǒng)。因此,探索小麥高溫脅迫響應(yīng)機(jī)制,了解小麥耐熱性的分子遺傳基礎(chǔ)及機(jī)理進(jìn)而提高其的抗熱性尤為重要。
Driedonks等[5]研究表明,高溫脅迫會造成植物細(xì)胞內(nèi)的氧代謝失調(diào),增加細(xì)胞發(fā)生氧化損傷的風(fēng)險。鄭 飛等[6]研究表明,高溫脅迫能顯著提高冬小麥旗葉中丙二醛(MDA)的含量,加速葉片衰老。劉洪展等[7]研究了不同高溫脅迫對小麥葉片中活性氧(ROS)代謝的影響,結(jié)果表明,在輕度高溫脅迫下,超氧化物歧化酶(SOD)活性增強(qiáng),過氧化氫酶(CAT)活性稍微下降;在嚴(yán)重高溫脅迫條件下,SOD活性最終表現(xiàn)出減弱的趨勢,CAT活性則有所上升。郭洪雪等[8]對二葉期小麥葉片分別進(jìn)行1 h、12 h、24 h和36 h 高溫處理,發(fā)現(xiàn)SOD活性及MDA含量呈現(xiàn)先上升后下降的變化趨勢。目前為止,還未見有關(guān)高溫脅迫影響小麥花藥不同生育期ROS代謝的研究。
高溫脅迫會導(dǎo)致植物細(xì)胞的ROS代謝平衡失調(diào),而ROS代謝平衡失調(diào)是引起作物雄性不育的重要生理原因之一[9]。前人研究結(jié)果表明,短時高溫脅迫(42±1)℃使小麥花藥徹底敗育(敗育率100%)[10]。處于生殖生長階段的小麥對溫度最為敏感,當(dāng)外界環(huán)境溫度達(dá)到一定的閾值時,可引起小麥花藥氧化還原體系的紊亂,造成花藥的氧化脅迫。因此,本研究以小麥不同發(fā)育時期的花藥為材料,通過對高溫脅迫下小麥花藥細(xì)胞內(nèi)ROS積累進(jìn)行觀察,以及比較分析ROS的生成速率和抗氧化酶的活性,揭示高溫脅迫下ROS代謝的動態(tài)變化與小麥花藥敗育的關(guān)系,探索小麥花藥高溫脅迫響應(yīng)機(jī)制,為進(jìn)一步解析小麥耐熱性的分子遺傳基礎(chǔ)及機(jī)理,提高小麥的耐熱性提供依據(jù)。
1 材料與方法
1.1 材料處理
供試小麥品種為西農(nóng)1376。待花藥發(fā)育至單核早期,于人工氣候室中進(jìn)行高溫脅迫處理,設(shè)置溫度分別為(42±1)℃(高溫脅迫處理)和(25±1)℃(對照),濕度為(70±5)%,每天處理3 h,共3 d,此時花藥已發(fā)育至單核后期。在4 ℃條件下分別收集單核后期、二核期和三核期的花藥,取樣時期的確定按Wang等[11-12]的方法進(jìn)行。
1.2 ROS染色
采用二氨基聯(lián)苯胺(3,3′-diaminobenzidine-HCl, DAB)染色檢測法[13],將不同發(fā)育時期的小麥花藥置于DAB溶液(0.1 μg·mL-1DAB, 50 mmol·L-1三羥甲基氨基甲烷(Tris)-醋酸緩沖液,pH 5.0)中,真空滲透 15 min,25 °C黑暗條件下 150 r·min-1振蕩24 h。染色后的花藥經(jīng)蒸餾水漂洗后置于85%乙醇溶液中70 °C水浴15 min,待葉綠素完全褪去后置于乳酸∶苯酚∶水(體積比1∶1∶1)溶液中保存,在光學(xué)顯微鏡下觀察并拍照。
1.3 酶液制備
稱取花藥1 g,在液氮中迅速研磨成粉末狀,加入 10 mL 預(yù)冷的提取液(0.1 mol·L-1Tris-HCl、pH 7.8,0.5 mmol·L-1EDTA 和1% PVP),然后于4 ℃下 3 000 g離心 10 min,取上清液,再以13 000 g離心 15 min,上清液即為酶提取液,低溫保存供酶活性測定。ROS指標(biāo)和酶活性指標(biāo)均重復(fù)測定3次。
1.4 超氧陰離子自由基產(chǎn)生速率及過氧化氫(H2O2)和丙二醛(MDA)含量的測定

H2O2含量按Patter-son等[15]的方法測定,并略有改動,取5%硫酸鈦0.1 mL和17 mmol·L-1氨溶液0.2 mL 加入酶液中,用丙酮沖洗5次,靜置沉淀后用5 mL 2 mmol·L-1硫酸銨溶解,在410 nm處測其吸光值,含量以 nmol·g-1FW表示。
MDA含量按Wang等[16]修改的方法測定,以nmol·mg-1protein 表示。即1 mL 0.6%TBA(硫代巴比妥酸)與1 mL酶液混合后100 ℃水浴加熱30 min,迅速冰浴降溫,二次離心后記錄 532 nm、600 nm、450 nm 處的吸光值。
1.5 酶活性的測定
SOD 活性的測定按王愛國等[17]的方法進(jìn)行,并略有改動,利用抑制氮藍(lán)四唑在熒光下的還原作用,將每3 mL反應(yīng)液(1.3 μmol·L-1核黃素、13 mmol·L-1甲硫氨酸、63 μmol·L-1NBT、0.05 mol·L-1pH 7.0磷酸緩沖液)中加入適量酶液后于4 000 lx熒光燈下光照15 min,以緩沖液作空白,在560 nm下測其吸光值。以抑制NBT 光化還原作用50 %的酶量為1個酶活性單位。
POD 活性按Wang等[11]的方法進(jìn)行測定。CAT 活性參照Ba等[18]的方法測定。
1.6 統(tǒng)計分析
試驗設(shè)置三個生物學(xué)重復(fù),每個重復(fù)進(jìn)行三次測量。采用Microsoft Excel 2010進(jìn)行數(shù)據(jù)整理,采用IBM SPSS Statistics 19軟件進(jìn)行差異顯著性分析。
2 結(jié)果與分析
2.1 高溫脅迫對小麥花藥細(xì)胞ROS的影響
正常發(fā)育的小麥花藥經(jīng)高溫脅迫后用DAB染色檢測ROS的積累情況,結(jié)果(圖1)發(fā)現(xiàn),正常發(fā)育的小麥花藥從單核后期至三核期DAB染色程度均較弱,染色斑點較少(圖1A~圖1C)。而高溫脅迫后的花藥從單核后期即可檢測到明顯的DAB染色斑點(圖1D),在花藥二核期時DAB染色強(qiáng)度最強(qiáng),并與對照的差異最明顯(圖1E),且在各發(fā)育時期花藥的DAB染色程度(圖1D~圖1F)均高于對照組。這表明高溫脅迫造成了花藥內(nèi)ROS代謝的紊亂,使花藥始終處于氧化脅迫的狀態(tài),進(jìn)而影響花藥的正常發(fā)育。

A~C:對照組; D~F:高溫脅迫組;A、D:單核后期;B、E:二核期;C、F:三核期。比例尺:0.5 mm。
2.2 高溫脅迫對小麥花藥中產(chǎn)生速率、H2O2和MDA含量的影響
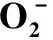
表1 花藥經(jīng)高溫脅迫后不同發(fā)育期的產(chǎn)生速率、和MDA含量
植物組織中積累的ROS會導(dǎo)致脂膜過氧化,產(chǎn)生MDA,其濃度是判斷脂質(zhì)過氧化程度和膜系統(tǒng)傷害大小的主要指標(biāo)[11-12]。MDA含量在對照組花藥的整個發(fā)育過程中均比較穩(wěn)定,而高溫脅迫后的花藥MDA含量則呈現(xiàn)先升高后下降的趨勢,且在二核期時達(dá)到最大值,為2.95 nmol·g-1,是對照組的1.59倍,差異顯著。此外,在單核后期和三核期時,MDA 的含量與對照相比差異顯著,分別為對照的1.09倍和1.34倍。這說明高溫脅迫對花藥細(xì)胞膜造成了持續(xù)的 損傷。
2.3 高溫脅迫對小麥花藥中SOD、POD 和CAT 活性的影響
由表2可知,在正常發(fā)育的小麥花藥中,POD活性從單核后期到二核期比較穩(wěn)定,分別為46.14和46.50 U·g-1FW·min-1,發(fā)育至三核期則急劇上升,達(dá)到86.66 U·g-1FW ·min-1。而高溫脅迫后花藥的POD活性呈現(xiàn)先升高再降低的變化趨勢,且始終顯著低于相同發(fā)育時期的對照組,在單核后期、二核期和三核期分別比對照組降低了61.38%、50.62%和88.37%。這說明在高溫脅迫的初期POD活性的增強(qiáng)清除了花藥內(nèi)過量積累的部分ROS,但隨著后期ROS的不斷積累,進(jìn)而抑制了POD的活性。

表2 花藥經(jīng)高溫脅迫后不同發(fā)育期的SOD、POD和CAT酶活性
此外,隨著小麥花藥的不斷發(fā)育,CAT活性在高溫脅迫和對照中均呈持續(xù)下降趨勢,在不同發(fā)育時期間,高溫脅迫的花藥下降速率顯著高于對照,其中對照的下降幅度為27.04%(單核后期至二核期)和10.90%(二核期期至三核期);而高溫脅迫后的花藥下降幅度為44.07%(單核后期至二核期)和66.52%(二核期期至三核期)(表2)。在相同發(fā)育時期,高溫脅迫后的花藥的CAT活性始終顯著低于相同發(fā)育時期的對照組,在單核后期、二核期和三核期分別比對照下降了 51.06%、62.48%和85.90%。
3 討 論
