流行性感冒
2020-07-23 08:44:58張智芳王曉歡梁小潔林志龍嚴延生
中國人獸共患病學報 2020年7期
張智芳,王曉歡,梁小潔,林志龍,嚴延生
流行性感冒(influenza)簡稱流感,由流感病毒(influenza virus)引起急性呼吸道癥狀,主要侵犯機體呼吸道和肺部。早期癥狀包括發熱、寒戰、頭痛、全身酸痛、乏力、嗜睡,肺部和呼吸道表現咽喉痛、干咳及鼻塞等。流感病毒主要分4型,即甲型、乙型、丙型、丁型,一般由甲和乙型致病人類,丙(C)型只發生在少數幼兒中,散發且癥狀不典型,丁(D)型宿主為牛,對人類基本不致病[1]。流感病毒起源不詳,可能幾個世紀前就有該病毒引起的疾病存在。該病是典型的人獸共患病,病毒宿主為禽類、豬及人類,有季節性流行和大流行之分。1930年英國首先分離出流感病毒,1935年美國研制出第一株流感疫苗。該病具有傳染性,病毒在人類中長期循環存在。
1 病原學特征
1.1形態結構及分類 流感病毒屬于正黏病毒科(Orthomyxoviridae)流感病毒屬,為單股、負鏈分節段RNA病毒。根據病毒核蛋白(nucloprotein, NP)和膜蛋白(membrant protein, MP)抗原特性不同,人類致病性流感病毒分為甲、乙、丙,即A、B、C 3型,分別用 IAV、IBV及ICV簡稱。A、B型核膜包裹8個RNA節段,C型有7個節段(缺少編碼神經氨酸酶蛋白節段)。用冷凍電子顯微鏡觀察自雞胚培養的兩株IAV,約80%病毒粒子呈球形并有包膜,直徑約為80~120 nm,其余病毒粒子呈多形性結構[2]。
IAV至少編碼產生11 個蛋白。病毒表面可凝聚紅細胞并具有抗原性的血紅素(hemagglutinin, HA),可和宿主受體唾液酸結合的神經氨酸酶(neuraminidase,NA),負責離子通道的基質蛋白2(matrix2, M2),由這3種蛋白構成病毒的雙層包膜。HA約占包膜總量80%,NA約占17%,M2蛋白量很微(每個病毒粒子只有16~20個分子)。HA的形態和功能類似冠狀病毒的刺突蛋白,主要起型抗原性及侵入細胞作用;NA主要起釋放子代病毒并清理唾液酸的作用。脂二層包膜主要成分是宿主膜的膽固醇。包膜的下層是基質蛋白1(matrix 1,M1),其內表層圍著組成病毒8節段vRNAs為核心的遺傳物質。vRNAs由核蛋白(nucleoprotein,NP)和微量的核運輸蛋白(nuclear export protein, NEP)包繞著每一節段的RNA組成,在vRNAs的一端還有稱為“3P蛋白”RNA依賴的多聚蛋白酶(RdRp),RdRp復合物由聚合酶酸性蛋白酶(polymerase acidic protein,PA)、聚合酶堿性蛋白酶1(polymerase basic protein1,PB1)和聚合酶堿性蛋白酶2(polymerase basic protein2,PB2)組成;由NP和NEP蛋白結合每一節段病毒RNA及頂端的3P蛋白構成病毒核糖核蛋白復合物(viral ribonucleoprotein comples,vRNP); IAV的非結構蛋白(non-structural protein,NSP)編碼抗病毒反應蛋白拮抗劑;PB1片段還編碼促凋亡蛋白PB1-F2[3-4]。見圖1。
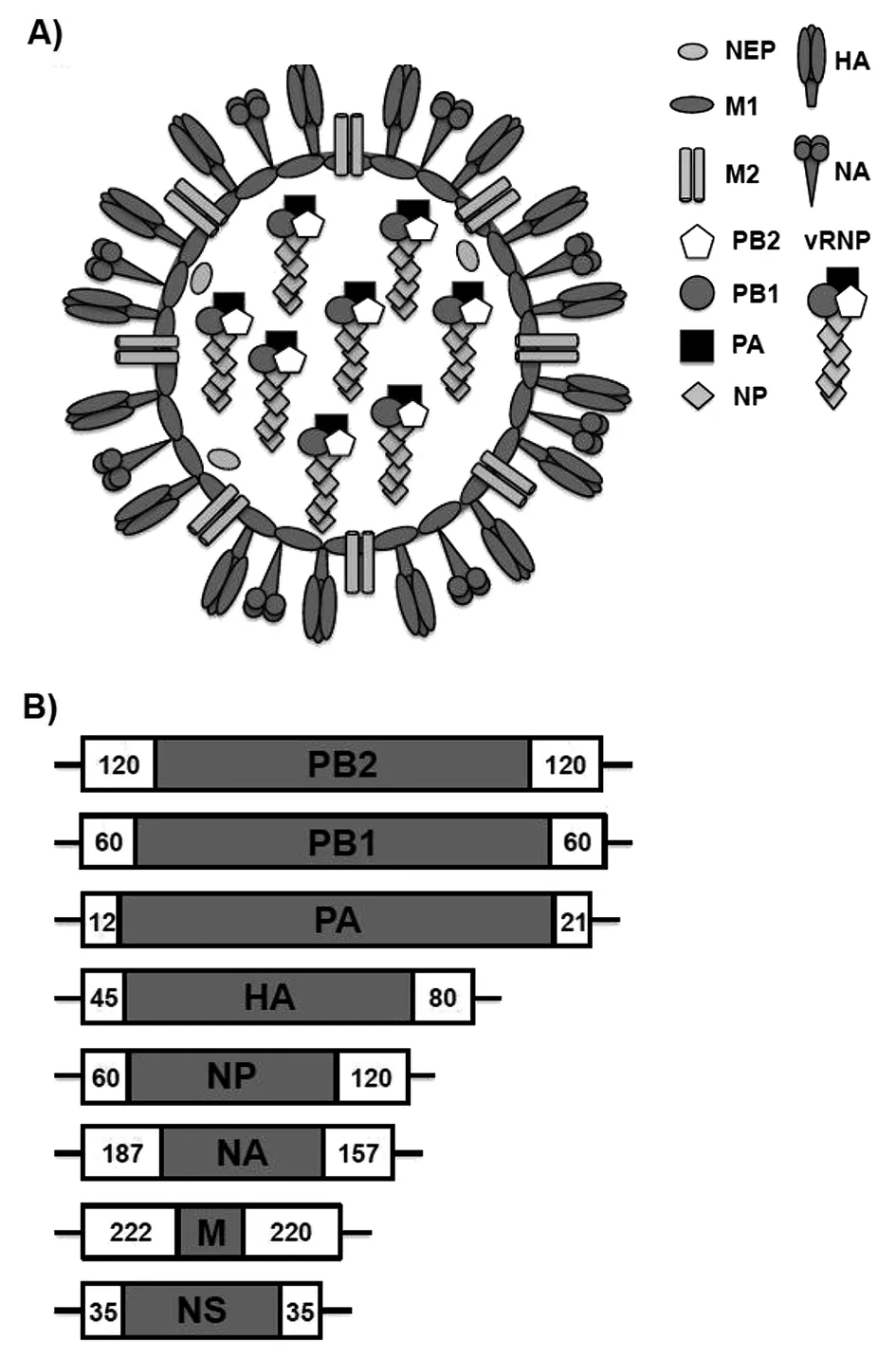
A) IAV的電腦模式圖。 HA:血凝素;NA:神經氨酸酶;NEP:核移動蛋白;M1:基質蛋白1;M2:基質蛋白2;PB2:聚合酶堿性蛋白酶2;PB1:聚合酶堿性蛋白酶1;PA:聚合酶酸性蛋白酶;NP:核蛋白;vRNP:病毒核糖核蛋白復合物。B) IAV 8節段單股、負鏈vRNA基因組的基因組成及其每一節段vRNA編碼的主要蛋白。如下:PB2, PB1, PA, HA, NP, NA, M, and NS。白框表示每個vRNA選擇性包裝入病毒粒子的信號,而數字則表示每個vRNA節段含有包裝信號的核苷酸長度,黑線代表病毒基因組復制和轉錄時可被病毒RdRp識別的3′~5′非編碼區(NCRs)。圖1 甲型流感病毒(IAV)病毒和基因組結構(引自參考文獻[4])Fig.1 Influenza A virus (IAV) virion structure and genome organization
迄今發現IAV有18個HA亞型(H1~H18),11個NA亞型(N1~N11),由這些血清型組成各種流感病毒株[5-6]。一般稱血清型 H1~H3株為季節性流行株,其他組合型大多為禽流感株。2012~2013年分別從蝙蝠中測得核苷酸序列H17N10和H18N11,但病毒株至今仍未分離到。IBV只有山形(Yamagata)和維多利亞(Victoria)兩型[7]。
流感病毒按以下公式命名分類:型別/宿主/分離地點/毒株序號(指采樣時標本號)/分離年代(血凝素和神經氨酸酶亞型)[8]。
1.2IAV的生命循環 IAV的感染和產生子代周而復始的過程即是的IAV生命循環(見圖2)。首先IAV的HA與宿主唾液酸的結合起始感染過程,因此,從感染來說,其宿主受體是非常重要的。在人類主要位于上呼吸道的α2,6鍵結合半乳糖的唾液酸,禽類則主要在于下呼吸道的α2,3鍵結合半乳糖的唾液酸[9],而在豬上呼吸道既有α2,6鍵結合半乳糖的唾液酸、也有α2,3鍵結合半乳糖的唾液酸,因此把豬稱為流感病毒的混合器。
IAV HA的酸化(pH5~6)裂解出了HA1和HA2,其次HA2與宿主上皮細胞(巨噬細胞)內小體膜融合,M2進一步酸化促使M1釋放vRNP進入宿主細胞質中,通過病毒NP,vRNPs被轉運到病毒復制和轉錄的細胞核,進行vRNP的復制與轉錄。由進入細胞核的病毒RdRp操控了細胞的解旋酶、復制酶等,首先形成發夾狀RNA,合成正鏈RNA,再轉合成具有IAV特征互補的負鏈RNA;其次轉錄IAV mRNA,這種轉錄需要有引物引導,病毒 RdRp識別PB2活化了宿主細胞核的5′端帽酶,截取或偷竊(cap-snatching)了12~14個核苷酸作為引物[3],而3′端聚腺苷酸尾則由IAV 自帶的RdRp序列作為引物轉錄mRNA,原則上這種轉錄機制可以轉錄單個PB2、HA、NA和NA蛋白和多個的PB1、PA、M和NS蛋白。在細胞核中復制的IAV遺傳物質和轉錄生成的蛋白由NP及NEP在β肌動微纖毛的協同配合下移出細胞核,其中HA、NA和M2被轉運到細胞的內質網中進行脂或糖基化修飾,最后由信號引導至細胞膜極性化頂端,等待包裝好的vRNPs從極化的細胞膜頂端出芽生殖(圖 2),按信號裝配成功的子代病毒粒子具有感染性可以再感染周邊細胞,完成了IAV的第一代生命循環。而在復制、轉錄等過程不能按照裝配信號要求形成正確的8節段的vRNPs的子代病毒粒子或缺失某蛋白的子代病毒粒子則不能再感染周邊細胞[10]。
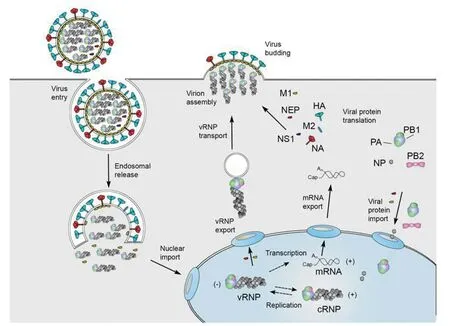
注:圓型為病毒粒子,方框示細胞內環境,方框內藍色橢圓型部分為細胞核;各符號代表見圖1。圖2 甲型流感病毒(IAV)生命循環圖(引自參考文獻[10])Fig.2 Influenza A virus life cycle
2 流行特征及過程
流感一般分為流感大流行、季節性流感及禽流感。流感大流行一般難以預測,往往涉及禽或豬類流感病毒節段基因的重排。季節性流感指引起常年人類的流感,其抗原性的變化多為病毒的遺傳物質發生“漂移”引起;其流感病毒亞型為H1~H3。據WHO報告,全球每年因流感發病300~500萬人,嚴重感染者有約29~60萬例死亡,是全球一大公共衛生的高負擔疾病。禽流感有高、中、低致病性之分(致禽類死亡程度而分),高致病性禽流感一般為H5N1。2013年3月始,我國曾一度發生低致病性禽流感H7N9在活禽市場引發人感染事件,多年來感染病例合計1 537例;2017年2月份在廣東發現的H7N9發生2個位點的變異而可能成為高致病性禽流感株后[11-12],同年9月在農業部的協調下,約有85%的禽類用含H5和H7的二價獸用疫苗免疫接種,此后終止了H7N9禽流感病毒的變異及感染人。我國每年禽類養殖在50億羽以上,由于禽流感的流行可引起人員的死亡以及因疫情需要撲滅禽類,引起巨大的經濟損失[12]。其他禽流感病毒亞型一般為低致病性,偶爾引起死亡病例發生。
2.1傳染源、傳播途徑和易感人群 引起季節性流感發生的傳染源為人類循環散發的流感患者,其本質是HA或NA病毒抗原發生位點突變的結果。豬和一些禽類在禽流感傳播中可作為傳染源。傳播途徑一般是經呼吸道途徑,由飛沫或密切接觸傳播;從某種程度來說,全人群為流感易感人群。但按WHO分類,需接種流感疫苗的易感人群分為5類[13]:①孕婦;② 6個月~5歲兒童;③65歲以上老者;④ 患除單純高血壓外的基礎疾病及免疫低下者;⑤ 醫務工作者。
2.2時空分布 流感一般全年可以發生,但病例多寡有季節性分布。所謂季節性流感,在全球有南北半球的流行。在我國以長江為界,南北流行也很明顯,北方約發生在每年的 11月至次年的2月份,即在冬季流行;南方如福建省、廣東省,一年有兩次流行,一次在夏季,約7~8月份,另一次在冬季。
甲型流感一般3~4年一次小流行,10~15年一次大流行,這種周期性流行與亞型病毒變異及人群免疫水平有關。乙、丙型周期性不明顯。流感在全球各地均可發生,但不同地區間因氣候因素、人群密集程度、交往頻度、傳染源數量、人群免疫情況、病毒抗原變異及防控策略措施不同其發病率出現較大差別。一般而言流感流行是先城市后農村,先平原地區后山區,沒交通線路蔓延。
2.3流行狀況 持續不斷的流感流行,主要是由于流感病毒HA和NA的抗原易變性,抗原變異幅度大小直接影響流感流行的強度和規模。小幅度的變異即抗原漂移(drift),屬于量變,其結果多引起中小型流行;而抗原轉換(shift)即發生了較大幅度的抗原變異,形成新亞型,是質變,其結果常引起世界性大流行。
1918年大流感的死亡率高達3%~5%,其他3次全球流感大流行的發病率與死亡率也不能與1918年的比較。而作為季節性流感來說,其死亡率一般在0.003%~0.005%之間。
2.3.11918年流感大流行 1918年的流感大流行始于1918年的春天,一個不同尋常的特征是大多數的死亡病例發生在15至34歲的健康成年人中,要知道,這些身體健康的年輕人相對老幼來說擁有更強大的免疫力和自我修復力。而這場瘟疫在全球范圍內流行,根據保守的統計數字,它至少感染了5億人,死亡人數為2 500~5 000萬人,而因為很多偏遠地區和戰亂無法統計,有學者認為,死亡人數超過一億,而當時世界的全部人口也只有17億而已。
Taubenberger等[13]從1918年一死者的福爾馬林固定、石蠟包埋肺部組織中用RT-PCR擴增到4個節段9個基因的DNA片段,并進行測序分析,證實當初流行的IAV為H1N1[13]。2005年,Taubenberger等[13]又從埋葬于阿拉斯加凍土帶下1918年因流感死亡的一女尸肺中獲得標本,將其復活為具有活性的H1N1病毒,作為對照,取一經修飾暫稱為德州的IAV,該H1N1病毒感染肺細胞株、小鼠與德州IAV進行比較,感染1 d后從肺細胞株中釋放的病毒50倍高于德州IAV,感染2 d后小鼠失重13%,感染4 d后該病毒在小鼠肺部中的量高于德州IAV,6 d后感染該復活的H1N1小鼠全部死亡。當時認為該病毒可制成最有毒力的生物戰劑[14]。
2.3.21957年H2N2大流行 1957年2月,我國西南貴州省首先檢測出該流感的流行,4月份蔓延至中國香港,導致發生250 000個病例;此后5月,流行至日本和新加坡。該流感最終造成全球100~400萬死亡,是中華人民共和國成立后流行最嚴重的流感。
1957年下半年第二波流感再度肆虐我國各省,雖然采取了關閉公共設施如學校、電影院等措施,但缺醫少藥造成不少的病例死亡[12]。同年,這一事件促使國家成立了流感中心(Chinese National Influenza Center,CNIC)進行流感的監控。
由于流感對人類的威脅,因此1947年WHO開展了全球流感項目,1952年成立了全球流感監測網絡(the Global Influenza Surveillance Network,GISN),1957年的流感大流行即成為GISN的首個監測項目[12]。
2.3.31968年流感大流行 1968年7月大流行的流感株由H3N2引發。首先從香港突發病例,約15%的香港人群發病。疫情擴散蔓延至印度及澳大利亞北部城市。此后,1968-1969 冬疫情流行至北半球國家,全球約有(100~400)萬死于該流感流行。我國雖處于“文化大革命”期間,大多數公共衛生資料沒有收集。但由廣東、四川、上海、北京、哈爾濱和青島等仍有流感樣病例(influenza-like illness,ILI)報告給CNIC,統計分析認為1968 年的年度報告的ILI 仍是1968-1992年報告的最高年份[12]。
2.3.42009年 H1N1pdm09流感大流行 2009年4月份墨西哥和美國等報告了甲型流感的流行,美國首先檢測確定了該流感是由甲型H1N1流感病毒引起。 H1N1病毒的全基因組核苷酸測序表明,該病毒是由北美豬系的H1N1和歐亞豬系H1N1流感病毒的基因組重組而成,因此稱為“豬源性甲型流感病毒”,以H1N1pdm09稱之,區別現季節性H1N1 IAV。
2009年4月25日,根據《國際衛生條例》的規定,世衛組織(WHO)總干事宣布2009年 H1N1疫情為國際關注的突發公共衛生事件。4月27日,WHO將流感大流行疫情警戒級別從3級提高到4級;4月29日,WHO又把流感大流行的警戒級別從4級提高到5級,表明大流行迫在眉睫,6月11日,WHO將全球流感大流行警戒級別再升至6級。這是WHO 40年來第一次把傳染病警戒級別升至最高級別。本次疫情相對多只引起輕癥發生,但也蔓延到214個國家和地區,導致近20萬人死亡,歷時一年,2010年4月10日宣布本次大流行結束。
這次大流行美國估計發生大約6 080萬例感染(95%可信限為4330萬~8930萬例),27.4萬例住院治療(195 086~402 719),死亡12 469例(8 868~18 306)。我國報告12.7多萬例病例,住院治療4 859例,死亡病例800多例。
本次大流感促進了我國進一步加強流感監測網的投入和建設,監測實驗室增至411個,哨點醫院增至556個[12]。
2.4禽流感在我國的發生 1997年及2005年分別在香港及我國內地發生高致病性禽流感的流行;2013年始迄今,我國流行H7N9禽流感,2017年2月份我國監測發現H7N9基因組發生2個位點的突變,有可能導致高致病性禽流感的流行。2017年9月我國對禽類進行了H5和H7二價獸用流感疫苗的接種,此后至今僅5個人病例的發生,合計病例1 537例,病死率高達近40%。其他亞型禽流感病例也引起個例死亡病例發生,但很快就被撲滅。禽流感的發生,引起很大的經濟和人員死亡的損失,以致引起我國主要城市“白條雞鴨”上市的改革[12]。
3 重癥流感的病理機制
重癥流感的引發肺炎病理機制有2種,季節性流感或流感大流行大多為隨后感染化膿性細菌引起肺炎的發生[15-16],但也有少數是由于病毒的大量復制引發“細胞因子風暴”使肺部積血、充水、纖維化而致重癥;H5N1、H7N9禽流感病毒和引發個別死亡的低致病性禽流感病毒也是由于肺部“細胞因子風暴”而致重癥的[17]。
4 診療及實驗室檢測
4.1流感治療 流感致病性與感染的毒株型別侵染有關,臨床表現也不盡相同。個體免疫狀況與感染也有密切關系,出現隱性感染或顯性感染。一般感染流感后的潛伏期很短,為1~4 d,平均2 d。起病急,伴有發熱(38 ℃或以上),全身中毒癥狀明顯,表現為頭疼、肌疼和嚴重不適,上呼吸道癥狀明顯;嬰幼兒有高熱驚厥及拒奶等現象[18]。
流感的藥物治療一般抑制流感病毒的3個靶點,一是離子通道M2抑制劑,以金剛烷胺或金剛乙胺治療,本世紀已發現該位點全部發生突變,現已不用該抑制劑治療。二是神經氨酸酶(NA)抑制劑, 目前用磷酸奧司他韋(達菲)在流感發病48 h之內用該藥治療有效,該藥2000年初被美國FDA批準用于13歲以上人群的治療[19],但流感患者常超過時間就診,臨床醫生發現在加倍使用奧司他韋情況下對此類還有作用;扎那米韋(zanamivir),因藥效不穩定,我國未批準使用。三是RNA聚合酶抑制劑,該藥有法匹拉韋和巴洛沙韋(Baloxavir),前者已在日本上市,美國也批準使用,但其使用范圍主要為輕癥;后者仍處于研究階段。用流感恢復期血漿治療也有用于重癥流感治療,但需要注意其中和抗體效價及安全性。季節性流感的治療還配用抗生素(一般對老年人及幼童),以預防細菌隨后侵入感染引起的大葉性肺炎。除此以外,一般以支持療法或臥床休息為主。
4.2實驗室檢測 一般三大常規檢測更要注意外周血象檢測結果。白細胞計數范圍廣,沒有發現明顯的由細菌續發感染引發的肺炎癥狀時,看不出白細胞增高情況;中性粒細胞和淋巴細胞是白細胞的重要組成部分,特別是中性粒細胞占比達50%~70%,是對抗入侵細菌最重要的防衛細胞,特別是急性化膿性細菌入侵時,會引起中性粒細胞的增高;重癥患者的淋巴細胞數量降低;另一重要指標是C反應蛋白(CRP),在感染、炎癥等應急狀態下,血漿中CRP會迅速增高[19]。
實驗室直接檢測流感病毒是特異性檢測方法,現最精確的做法是用咽拭子采取口咽部標本提取RNA作qRT-PCR檢測。因分離病毒較耗時,已較少用于疾病的早期診斷,但在病毒抗原變異監測中仍發揮重要作用。血清學檢測一般用于感染或疫苗接種后評估流感病毒特異性抗體水平高低,常用的方法有血凝抑制試驗(HIA)、中和試驗和酶聯免疫試驗(ELISA)等,用于早期診斷,與qRT-PCR方法相比,其敏感性和特異性均較低,已基本不采用。
5 防治對策
目前尚無有效的控制流行性感冒流行的措施,為一世界性問題,一旦出現大流行,各國都難幸免。防治對策主要以降低發病率和死亡率為目的,以反復疫苗接種和各國協調一致的疫情監測和控制為重點。
5.1策略的實行 該策略包括3個部分,簡介、概論和策略的實行。其中概論包括了4個應對內容措施。一是強調對公共衛生難以解決的問題需進行研究和創新;二是對全球流感的監測、溯源和資料利用;三是擴展季節性流感的預防控制和保護易感人群;四是為使全球更安全,要切實加強流感大流行的準備和應對。在策略的實行里也包括兩大內容,一是概述2002年以來的主要對流感防控(包括對SARS、MERS等呼吸道病毒性疾病)成效;二是在2019-2030年流感應對策略中可能遇到的預防、控制等出現的問題和對策。客觀地說,2002年以來的流感防控策略是取得了很大的成效,全球從1952年開始的全球流感系統的監測協作從最初的幾個發展到目前110個國家的150個研究機構的參與;期間平息了SARS疫情;2005年制定了可操作的《全球衛生條例》(International Health Regulation,IHR);發現并控制了MERS的流行;開展了對醫院SARI病例的報告和診斷,促進全球各地在流感(或呼吸道病毒性疾病)的防控、診斷和應對(包括防護衣帽、消毒物質的儲備等)。
要是沒有這樣的儲備,應對2019年末發生的新冠肺炎的流行可能情況更糟。
5.2WHO注重流感疫苗對流感的應對 WHO在2019-2030年全球流感應對策略中,特別強調采用疫苗保護易感人群的作用。流感疫苗每年需要通過全球100多家流感實驗室報送當年流感流行的亞型,并經WHO的5個參比實驗室經抗原測定,每年2次例會推薦企業生產用的疫苗株。對于多數疾病而言,目前主要使用五類疫苗,包括滅活苗、減毒苗、活病毒載體苗、基因重組苗和DNA或mRNA苗。但對于流感疫苗來說,主要是滅活苗(雞胚苗)、活病毒載體苗(MDCK等細胞苗)兩類。前者至少占全部苗的90%以上,又可分為主要含病毒顆粒的甲醛滅活苗、裂解苗和亞單位苗3種;甲醛滅活苗基本上保存了完整的病毒顆粒,其預防免疫效果也最為突出,但副作用稍大些;裂解苗系由完整的病毒顆粒經裂解后去除病毒RNA等,暴露出各病毒組成成分,免疫效果好,副作用也小,目前主要是這類疫苗在使用;亞單位苗,主要是血凝素HA成分,免疫效果小于裂解苗,但副作用較之裂解苗也小,這種苗大多用于兒童和65歲以上老人群中。活病毒載體苗主要是表達HA蛋白苗或添加有細胞免疫和/或體液免疫作用的佐劑(如MF59等)苗;流感疫苗目前主要是這4種。但因為抗原“漂移”的緣故,其保護作用一般只能達到60%,有的年份因預測不準或其他因素使選擇的疫苗株不能匹配,還無法達到這樣的保護效果,這也就是為什么流感疫苗需要年年注射接種預防的主要原因。
利益沖突:無
