云南臨滄大雪山自然保護區非飛行小型獸類物種多樣性調查
2020-07-23 10:23:48龔正達李華昌楊貴榮劉起勇
中國人獸共患病學報 2020年7期
關鍵詞:物種
馬 英,魯 亮,龔正達,李華昌,楊貴榮,劉起勇
哺乳動物是動物類群中分類學和物種多樣性研究非常深入的類群[1],近20年來分子系統學的研究依然揭示著該類群中存在大量被形態學研究忽視的種類[2]。小型獸類一般指兔形目的鼠兔屬、嚙齒目、勞亞食蟲目和翼手目等小型哺乳動物,其中翼手目以外的類群統稱為非飛行小型獸類(Nonvolant small mammals)。非飛行小型獸類在各種生態系統中發揮著重要的功能,如參與植物種子擴散、草場土壤更新等重要的生態環節[3-4]。在食物鏈中,小型獸類作為初級消費者,為上層的捕食動物提供了大量的食物,是食物鏈的重要組成部分[5]。因此,小型獸類的生物多樣性具有重要的生態學意義。同時,小型獸類在許多人獸共患病的保存和傳播過程中都具有重要意義,從而會影響生態系統中人群的健康[6]。相關研究表明,小型獸類傳播病原體與當地小型獸類的生物多樣性水平相關[7-10]。
目前,我國記錄的非飛行小型獸類的數量有350種[11]。通常在宿主動物多樣性和動物流行病學的調查取樣中,小型獸類種類的準確鑒定主要依賴專業的形態分類學專家。但現在大量的動物流行病學調查中往往沒有分類專家的實時參與,鑒定的準確性容易出現問題。而且,越來越多的研究發現形態學確定的小型獸類種類中存在很多形態差異很小的隱存種[12-13],這增加了鑒定的難度。DNA條形碼(DNA barcoding)技術是本世紀初發展起來的一項利用DNA序列進行物種鑒定的技術[14]。通過十多年的數據積累和各類群的研究,該技術被證明在鑒別許多動物類群中是可行的,包括各類群小型獸類中也是如此[15-20]。因此,我們可以利用DNA條形碼技術協助開展系統的小型獸類生物多樣性研究。
云南南部從上世紀80年代開始出現大范圍的家鼠鼠疫疫情,涉及到臨滄市的部分縣區[21]。為了掌握當地鼠疫宿主動物的構成,尤其是自然環境中各種非飛行小型獸類種類的構成,以便在鼠疫動物流行病學調查中能夠有明確的宿主動物分類學數據,能更好地分析各種野生鼠類在鼠疫動物間流行過程中的作用,我們開展了臨滄大雪山自然保護區及周邊地區中非飛行小型獸類的多樣性調查。為了能夠更好闡明當地非飛行小型獸類物種多樣性特征,我們在應用傳統形態學分類的基礎上,利用DNA條形碼技術對所獲標本進行了進一步鑒定。
1 材料與方法
1.1研究區域概況 臨滄大雪山自然保護區位于臨滄市臨翔區和云縣交界處,東靠瀾滄江,地理坐標:東經99°40′34″~100°22′01″,北緯23°17′52′~24°57′28″之間。南坡位于臨翔區境內,北坡為云縣地域。最高峰海拔3 429 m,因冬季有積雪,被當地稱為大雪山。整個保護區處于橫斷山區怒山山脈南段的高山縱谷地帶,最高峰與瀾滄江的相對高程2 600 m左右,形成了豐富的自然景觀,植被的垂直分布非常明顯。1 800~2 000 m范圍內為季風常綠闊葉林帶、2 000~2 600 m之間為中山濕性常綠闊葉林帶、2 600~3 000 m之間為針闊葉混交林帶、3 000~3 400 m之間為蒼山冷杉林帶和亞高山灌叢草甸帶[22]。
1.2 方 法
1.2.1調查時間和范圍 調查工作從2008年11月22日開始,至12月6日結束。在海拔1 200~3 200 m之間按照每200 m海拔進行一次調查的原則進行了14個點的調查。每天布放360個左右的鼠夾,第2 d上午檢查捕獲情況。調查點及其經緯度和海拔高度見表1。
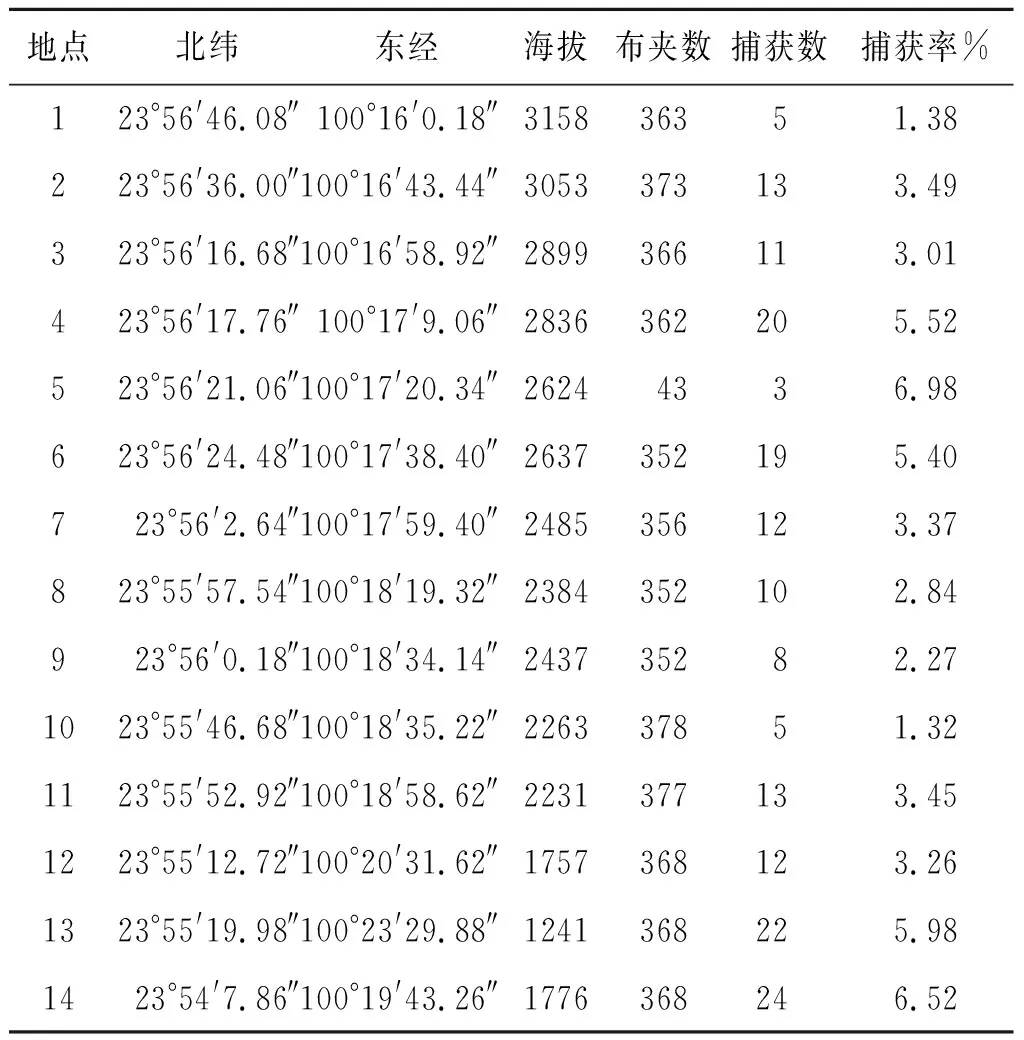
表1 采集點經緯度和海拔以及布夾和捕獲情況Tab.1 Location and altitude of trap sites, trap number and number of captures for each locations
1.2.2非飛行小型獸類種類鑒定 現場捕獲的非飛行小型獸類標本首先根據外部形態特征進行初步的分類。然后選擇皮張和頭骨沒有破損、發育成熟及形態有代表性的個體制作成假剝制標本,再根據頭骨形態和牙齒形態進行較為準確的鑒定。嚙齒目標本形態鑒定的依據是《中國嚙齒類》[23],其他小型獸類的形態鑒定參考《中國獸類野外手冊》[24]。
1.2.3 DNA條形碼鑒定
1.2.3.1組織樣品的保存 現場采集標本取肌肉組織用95%酒精浸泡保存,帶回實驗室后低溫冷凍保存。提取肌肉組織的總DNA時,使用QIAGEN DNeasy Kit(凱杰生物技術有限公司,上海),根據試劑盒的使用方法提取DNA,并冷凍保存備用。
1.2.3.2線粒體基因組細胞色素氧化酶亞基I(COI)基因片段的PCR擴增 在25 mL反應體系中,10×Buffer 2.5 μL,上下游引物各0.5 μL,dNTPs 0.5 μL,DNA模板1 μL,LATaqDNA聚合酶(寶生物工程有限公司,大連) 0.75 U。擴增條件為:預變性94 ℃ 5 min,變性94 ℃ 30 s,退火55 ℃ 40 s,延伸72 ℃ 1 min,共35個循環。其中引物首先采用BatL5310和R6036R[25],在不能擴增的情況下,選擇雞尾酒引物VF1LFt1和VR1LRt1[26]進行擴增。擴增產物直接送測序公司進行雙向測序。
1.2.3.3DNA條形碼的序列分析 首先用Chromos軟件觀察測序峰圖質量,如果峰圖質量很差不能準確判斷堿基,則重新進行擴增和測序。拼接后的測序結果用Clustal X[27]軟件進行多重序列比對,并進行適當的手工調整。同時將測定的序列在NCBI上運行BLAST程序進行序列同源性比較,以確保所獲得的序列是目標序列,排除擴增得到假基因序列的可能。根據BLAST結果,將相似度大于98%的序列作為參考序列用于系統樹構建及小型獸類的鑒定。對于GenBank中沒有近緣序列的個體,首先依據形態特征定種;對于難以用形態特征確定種類的食蟲類小型獸類,將頭骨照片和序列提交給中國科學院昆明動物所的何鍇博士進行鑒定。所有小型獸類中文名和學名參照“中國哺乳動物多樣性編目(第2版)”[11]。最后運用Mega6[28]軟件基于Kimura-2-parameter (K2P)[29]模型計算各物種的遺傳距離。采用鄰接法構建全部COI基因序列的NJ系統樹,對NJ樹進行內部分支檢驗與1 000次Bootstrap檢驗分析,來確定各支系的BS置信度。
1.2.3.4利用DNA條形碼進行樣品鑒定 使用DNA條形碼進行標本鑒定遵從2個原則, 一是序列是否與已知物種的序列構成單系[30],二是遺傳差異在一定的范圍之內。真獸亞綱哺乳動物COI基因的條形碼區域種間平均遺傳距離約為7.39%(K2P距離)[31],因此對于用于DNA條形碼的COI基因,本研究選用8%作為界定不同物種的標準。
當測得序列與參考序列構成單系,且單系的BS值大于等于0.95時認為二者為同一物種,當測得序列與參考序列不構成單系時,則認為并非同一物種。若遺傳距離中等(85%~92%),或測得序列與GenBank中幾個不同物種序列相似度相同時,認為該標本的分子鑒定只能精確到屬,但不能確定到種。
2 結 果
2.1小型獸類捕獲情況 在各種生境中,共捕獲各種非飛行小型獸類177只。其中,通過形態學鑒定確定其中131只屬于嚙齒目、勞亞食蟲目的17個物種,無法明確到種的有嚙齒目白腹鼠屬和勞亞食蟲目個體計46只(見表2)。制作的憑證標本中,白腹鼠屬的標本暫時保存在中科院動物所標本館,其他標本保存在臨滄市疾病預防控制中心。
2.2測序結果和系統樹的構建 由于中國鼩猬捕獲數量較多,所以只擴增了其中4個樣品的COI基因。通過兩套引物的擴增,從152份樣品中共成功擴增并測序了121份樣品(表2),擴增和測序成功率為79.6%。擴增片段的有效長度在650 bp左右,沒有發現片段的插入或缺失,證明擴增的片段確實為線粒體基因組的COI基因片段。所得序列都提交GenBank共享,各種對應的注冊號見表2。通過構建系統樹,發現這些序列形成了25個有效的分支,多個個體構成的分支都有100%的BS支持度(圖1)。其中,嚙齒目的鼠類共17種,分屬鼠科、田鼠科和松鼠科;勞亞食蟲目中的猬科2種,鼩鼱科6種。
根據系統樹的分支,可以發現臨滄大雪山地區白腹鼠屬的種類應該有6種,而不是根據形態特征鑒定的2~3種,說明白腹鼠屬中存在形態學的隱存種。此外,勞亞食蟲目的大量標本無法用形態特征予以準確鑒別,通過系統樹可以確定存在6個種。
3 討 論
3.1鼠科的隱存種問題 嚙齒目鼠科是分類學研究比較深入的一個小型獸類類群,但即使這樣,還有許多種類的分類地位存在爭議,鑒定特征不明確,同時一些類群中存在隱種。在臨滄大雪山地區的嚙齒動物中,白腹鼠屬的分類問題顯得尤其突出。在白腹鼠屬的傳統形態分類中,被毛中是否有刺毛是分類的重要特征[23,32]。但在臨滄大雪山高海拔生境中捕獲的白腹鼠標本往往都沒有刺毛,因此,區分安氏白腹鼠和北社鼠有一定的難度。同時由于地處中國和東南亞交界處,原本記錄在東南亞的白腹鼠(N.niviventer)在該地區也有分布[13,33],因此更增加了形態分類的難度。
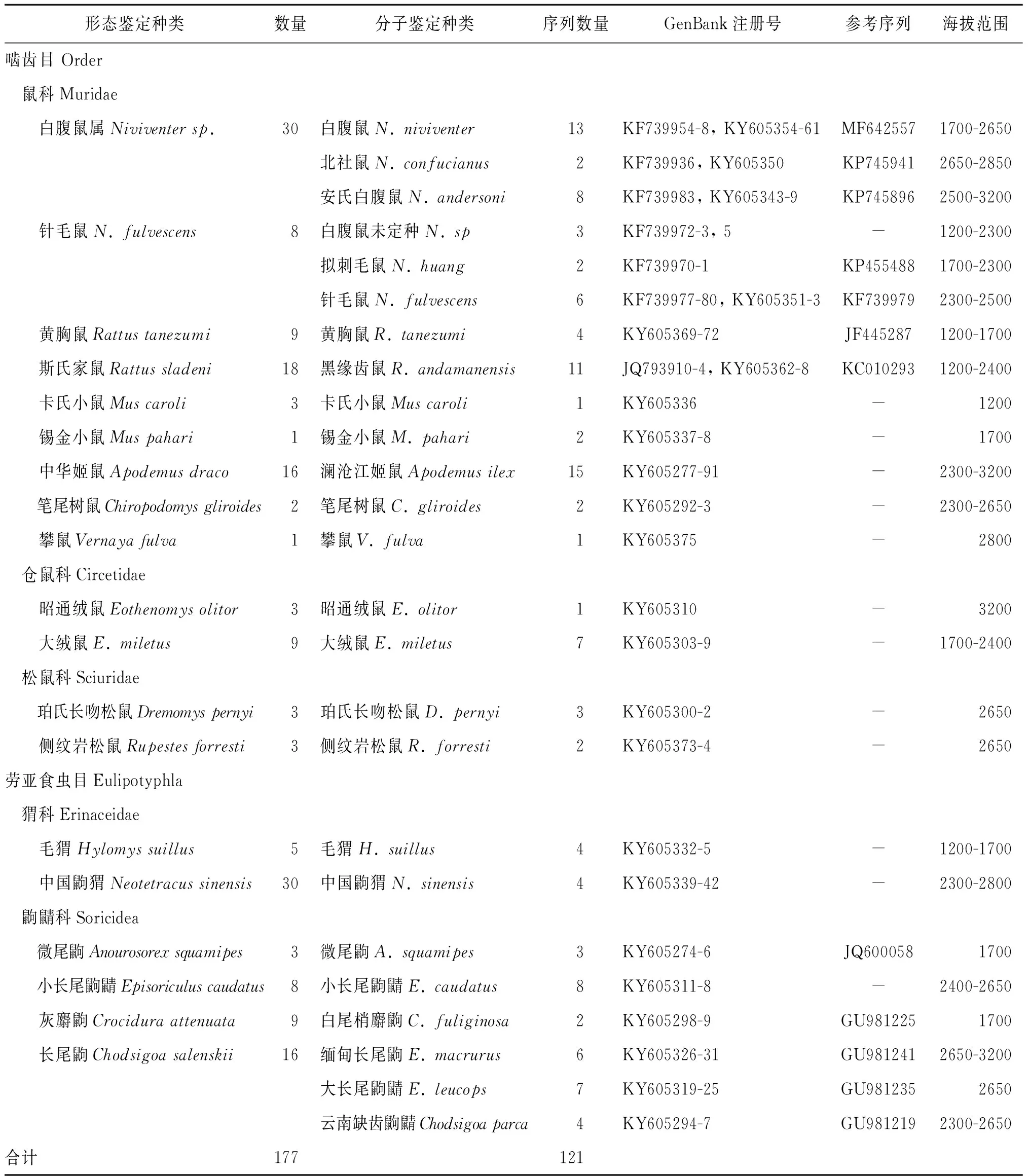
表2 捕獲標本的形態分類、分子鑒定結果和數量及相關信息Tab.2 Information of morphologic classification, identification with DNA barcoding, number and other related data

括號中為作為參考序列的GenBank注冊號,每個物種分支節點的自舉檢驗(BS)值標注在節點上方。圖1 基于COI 基因片段的臨滄大雪山小型獸類Neighbor-Joining系統樹Fig. The Neighbor-Joining phylogenetic tree of small mammals of Lincang mountain based on data of COI gene fragment.
在我國現有白腹鼠屬描述的種類中,被毛顏色鮮艷呈棕紅色調,背部硬刺毛發達的種類只有針毛鼠一種[23,34],但通過DNA條形碼技術和分子系統學研究,發現具備上述特征的白腹鼠屬的種類不少于3種[13]。在臨滄大雪山的生態環境中,這3種都存在,只是在海拔分布上存在一定的差異。這種現象說明,白腹鼠屬中存在明顯的隱存種現象,該屬的物種多樣性被遠遠低估了。
3.2不同生境中小型獸類的物種多樣性變化 臨滄大雪山不同海拔的植被類型大體可以分為季風常綠闊葉林帶(1 800~2 000 m)、中山濕性常綠闊葉林帶(2 000~2 600 m)、針闊葉混交林帶(2 600~3 000 m)、蒼山冷杉林帶(3 000~3 400 m)和亞高山灌叢草甸帶,其中2 600 m以下為北亞熱帶,2 600 m以上為南溫帶[22]。在2 000 m以下地勢較緩的區域,往往是農耕區;2 000~2 400 m為退耕還林區域,多為經濟林;2 400 m以上則成為保護區管轄區域,人類活動和干擾較少。
在臨滄大雪山3 000 m以上的冷杉林帶和亞高山灌叢中,捕獲率為2.45%,主要為瀾滄江姬鼠、昭通絨鼠、安氏白腹鼠和大長尾鼩等4種。在2 600~3 000 m之間的針闊葉混交林生境中,捕獲率為4.63%,小型獸類種類按捕獲數量排序有中國鼩猬、瀾滄江姬鼠、北社鼠、安氏白腹鼠、側紋巖松鼠、珀氏長吻松鼠和多種鼩鼱類等13種,包括筆尾樹鼠和攀鼠等稀有種類。在2 000~2 600 m的常綠闊葉林生境中,捕獲率為2.64%,包括中國鼩猬、北社鼠、針毛鼠、白腹鼠未定種、大絨鼠、黑緣齒鼠、瀾滄江姬鼠和鼩鼱類等11種。在2 000 m以下的生境中,農田較多,捕獲率達到5.25%,捕獲小型獸類包括黑緣齒鼠、黃胸鼠、灰麝鼩、毛猬、短尾鼩、大絨鼠、白腹鼠未定種和擬刺毛鼠等11種,其中與人類活動關系密切的小型獸類有兩種大鼠屬種類。可以看出,海拔2 000~3 000 m的生境中,小型獸類的物種多樣性是最高的。該結果與其他山地多樣性調查的結果一致,說明在山地環境中,中海拔生境是物種多樣性最為豐富的生境。
3.3不同小型獸類種類的海拔分布 根據不同小型獸類種類的采集記錄,可以發現這些小型獸類在臨滄大雪山的海拔分布范圍基本與其在我國的緯度分布基本是對應的。以白腹鼠屬的種類為例[13],安氏白腹鼠(N.andersoni)分布在我國青藏高原東緣一直到秦嶺的高海拔地區,相應的,在臨滄大雪山該種也分布在較高海拔(2 500~3 200 m)。北社鼠(N.confucianus)在青藏高原東南緣沿大巴山、秦嶺、太行山一直分布到華北地區[16],在臨滄大雪山則分布在安氏白腹鼠分布范圍的下緣。針毛鼠(N.fulvescens)基本分布于我國華南、西南南部地區,而在臨滄大雪山則分布在2 300~2 500 m之間的生境中,白腹鼠(N.niviventer)的分布一般認為在東南亞地區,在臨滄大雪山則分布于較低海拔的生境中(1 700~2 650 m)。臨滄大雪山捕獲的鼩鼱科種類也都是東洋界物種[35]。這些種類在大雪山的分布基本也在2 650 m海拔以下地區,和白腹鼠屬東洋界種類的海拔分布有相似之處。
(致謝:感謝臨滄市疾病預防控制中心的其他工作人員在現場調查中的工作,感謝臨滄大雪山自然保護區的兩位護林員在現場調查中予以的熱心幫助。感謝中國科學院昆明動物所的何鍇博士在食蟲類小型獸類的分子鑒定給予的幫助。)
利益沖突:無
本文引用格式:馬英,魯亮,龔正達,等.云南臨滄大雪山自然保護區非飛行小型獸類物種多樣性調查[J].中國人獸共患病學報,2020,36(7):572-577,582. DOI:10.3969/j.issn.1002-2694.2020.00.051
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
