初生休眠解除狀態和干燥處理對水曲柳種子萌發的影響
2020-07-14 09:03:46崔程程
植物研究 2020年4期
盧 艷 閆 月 崔程程 張 鵬
(東北林業大學林學院,森林生態系統可持續經營教育部重點實驗室,哈爾濱 150040)
種子休眠是指具有正常生活力的種子在適宜條件下不能萌發的現象[1]。種子休眠依據休眠形成因素的不同,分為初生休眠和次生休眠[2]。休眠可以防止種子在惡劣的環境中萌發[3],這對種子保持其自身的繁衍能力極為有利,但種子休眠給農業生產帶來極大不便,播種前必須打破休眠。
水曲柳(FraxinusmandshuricaRupr.)是東北地區重要的珍貴用材樹種,是“東北三大硬闊”之一,具有重要的經濟、生態價值。水曲柳主要通過種子繁殖,由于其種子具有生理形態休眠,休眠程度較深,播種前需進行變溫層積處理打破初生休眠[4]。有研究表明,解除初生休眠的水曲柳種子在較高溫(20℃以上恒溫)條件下萌發會進入次生休眠(熱休眠)[5],生產中播種時如遇較高溫天氣,種子會進入熱休眠,嚴重影響育苗生產。
目前對水曲柳種子初生休眠[4,6~9]和次生休眠[10]的解除方法已有一定研究,但水曲柳種子初生休眠的解除狀態對種子次生休眠是否有影響尚不清楚。以往研究發現解除休眠的水曲柳種子經室溫干燥處理后發芽率和發芽時間與干燥前相比沒有明顯變化,這一研究結果表明已解除休眠的水曲柳種子可以進行再干燥處理[11]。解除休眠的水曲柳種子經干燥處理后可以隨時進行播種育苗,方便播種育苗工作的進行,但干燥處理對初生休眠解除狀態不同的水曲柳種子萌發的影響是否相同有待驗證。
本文以具有初生休眠的水曲柳種子為試驗材料,研究經不同裸層積和干燥處理的水曲柳種子在適宜溫度和較高溫條件下的萌發表現,擬解決以下問題:①確定初生休眠解除狀態不同的水曲柳種子在不同溫度下的萌發表現是否相同;②確定干燥處理對初生休眠解除狀態不同的水曲柳種子萌發的作用。通過上述研究,可以為水曲柳播種育苗工作中的種子處理提供技術參考,對農林業生產有一定的意義。
1 材料與方法
1.1 試驗材料
試驗用種采自吉林省露水河林業局成年水曲柳母樹上,種子進行4種裸層積處理(視為種子初生休眠解除狀態不同),①暖溫(18℃,下同)10周+低溫(5℃,下同)8周,記為W10+C8;②暖溫12周+低溫8周,記為W12+C8;③暖溫10周+低溫10周,記為W10+C10;④暖溫12周+低溫10周,記為W12+C10,以層積處理結束后的種子為試驗材料。
1.2 試驗方法
種子經層積處理結束后分為兩組,一組立即進行發芽試驗;另一組置于室溫(20~23℃)條件下自然陰干4 d(含水率為7%~8%),然后將干燥處理的種子用自來水浸泡48 h后進行發芽試驗。
發芽試驗:將種子用0.5% KMnO4溶液浸泡消毒30 min,隨后將種子上的KMnO4用自來水沖洗干凈。每種處理4次重復,25粒種子/重復,將種子置于培養皿(直徑9 cm)中,培養皿底部墊有一層濾紙,將培養皿放在無光照的人工氣候箱中,設置2種萌發溫度:①10℃(水曲柳種子的適宜萌發溫度);②25℃(誘導種子進入熱休眠的溫度)培養7 d后轉10℃。每天觀察記錄種子的萌發情況,以胚根突破種皮超過2 mm作為萌發標志。在種子置床后的第30天計算最終的發芽率、發芽指數、平均發芽時間。計算公式如下:
發芽率(GR)=發芽種子數/供試種子總數×100%
(1)
發芽指數(GI)=∑(ng/tg)
(2)
平均發芽時間(MGT)=∑(ng×tg)/∑ng
(3)
式中:tg為發芽時間(d),ng為與tg相對應的每天發芽種子數。
1.3 數據統計分析
采用Excel 2010(Microsoft)進行數據整理,用SPSS Statistics 20.0(IBM)進行層積處理與干燥處理對水曲柳種子萌發能力影響的方差分析,方差分析結果顯著時,采用Duncan法進行多重比較分析。其中發芽率為百分數,對其進行反正弦平方根轉換后再進行方差分析。
2 結果與分析
2.1 不同層積及干燥處理的水曲柳種子在適宜溫度下的萌發表現
萌發溫度為10℃時,層積處理、干燥處理以及兩者的交互作用對水曲柳種子發芽率的影響均極顯著(P<0.01)(如表1所示)。未經干燥處理的種子,層積時間越長種子發芽率越高,W12+C10處理種子發芽率顯著高于其他層積處理,其他層積處理之間種子發芽率差異不顯著(如表2所示)。干燥處理的種子,干燥前后種子發芽率變化受種子低溫層積時間影響,低溫層積時間較短(8周)的種子經干燥處理后發芽率沒有明顯變化,低溫層積時間較長(10周)的種子經干燥處理后發芽率顯著下降(如表2所示)。
表1 不同溫度下層積與干燥處理組合對水曲柳種子萌發能力影響的方差分析
Table 1 Variance analysis of the effect of the combination of stratification and drying treatment on the germination ability ofF.mandshuricaseeds at different temperatures

變異來源Sourceofvariation10℃25℃培養7d后轉10℃Cultivatedat25℃for7dandthentransferedto10℃發芽率Germinationrate發芽指數Germinationindex發芽時間Germinationtime發芽率Germinationrate發芽指數Germinationindex發芽時間Germinationtime層積處理Stratification0.0070.0210.0220.0140.0510.399干燥處理Dryingtreatment<0.001<0.001<0.001<0.0010.0010.055層積處理?干燥處理Stratification?Dryingtreatment<0.0010.0010.3670.0090.0060.057R20.8120.8420.6790.8220.6260.429
表2 不同層積及干燥處理的水曲柳種子在10℃下的萌發情況
Table 2 Germination ofF.mandshuricaseeds under different stratification and drying treatments at 10℃

干燥處理Dryingtreatment層積處理Stratification發芽率Germinationrate(%)發芽指數Germinationindex發芽時間Germinationtime(d)否NoW10+C888.00±3.65b3.61±0.30b7.08±0.53W12+C889.00±1.92b4.00±0.28b6.22±0.30W10+C1089.00±4.73b3.55±0.24b7.06±0.37W12+C1099.00±1.00a5.01±0.13a5.34±0.12是YesW10+C890.00±1.15b2.75±0.17c9.13±0.76W12+C892.00±1.63b2.91±0.18c8.49±0.58W10+C1076.00±1.63c2.62±0.09c8.00±0.36W12+C1068.00±1.63c2.41±0.12c7.80±0.31
注:表中數值為平均值±標準誤差。同列不同字母表示采用Duncan法進行多重比較差異顯著(P<0.05),下同。
Note:The figures in the table are mean±standard error.The different small letters in a column indicate significant difference at 0.05 level,the same as below.
層積處理對水曲柳種子發芽指數的影響顯著(P<0.05),干燥處理對水曲柳種子發芽指數的影響極顯著(P<0.01),層積處理與干燥處理的交互作用對水曲柳種子發芽指數的影響極顯著(P<0.01)(見表1)。未經干燥處理的種子,層積時間越長種子發芽指數越高,W12+C10處理種子發芽指數顯著高于其他層積處理,其他層積處理之間種子發芽指數差異不顯著(見表2)。干燥處理的種子,干燥前后種子發芽指數變化不受種子低溫層積時間的影響,各處理間種子發芽指數差異不顯著,但干燥處理種子的發芽指數顯著降低(見表2)。
層積處理與干燥處理的交互作用對水曲柳種子發芽時間的影響不顯著(P>0.05),但層積處理對水曲柳種子發芽時間的影響顯著(P<0.05),干燥處理對水曲柳種子發芽時間的影響極顯著(P<0.01)(見表1)。干燥處理極顯著地延長了種子發芽時間,無論種子是否經過干燥處理,層積時間越長的種子發芽時間越短(見表2)。
2.2 不同層積及干燥處理的水曲柳種子經較高溫培養7 d后轉入適宜溫度下的萌發表現
萌發條件為25℃培養7 d后轉10℃時,層積處理對水曲柳種子發芽率的影響顯著(P<0.05),干燥處理對水曲柳種子發芽率的影響極顯著(P<0.01),層積處理與干燥處理的交互作用對水曲柳種子發芽率的影響極顯著(P<0.01)(見表1)。未經干燥處理的種子,層積時間最長(22周)的種子發芽率最高,其他層積處理之間種子發芽率差異不顯著(見表3)。干燥處理的種子,干燥前后種子發芽率變化受種子低溫層積時間影響,低溫層積時間較短(8周)的種子干燥處理后發芽率有明顯下降,平均下降約45%,而低溫層積時間較長(10周)的種子干燥處理后發芽率急劇下降,平均下降約73%(見表3)。
層積處理對水曲柳種子發芽指數的影響不顯著(P>0.05),干燥處理對水曲柳種子發芽指數的影響極顯著(P<0.01),層積處理與干燥處理的交互作用對水曲柳種子發芽指數的影響極顯著(P<0.01)(見表1)。未經干燥處理的種子,層積時間越長種子發芽指數越高,W12+C10處理種子發芽指數顯著高于其他層積處理,其他層積處理之間種子發芽指數差異不顯著(見表3)。干燥處理的種子,干燥前后種子發芽指數變化不受種子低溫層積時間的影響,各處理間種子發芽指數差異不顯著。除W12+C10處理外,其他層積處理種子的發芽指數不受干燥處理的影響,但W12+C10處理種子發芽指數受干燥處理影響較大(見表3)。
層積處理、干燥處理以及兩者的交互作用對水曲柳種子發芽時間的影響均不顯著(P>0.05)(見表1)。
表3 不同層積及干燥處理的水曲柳種子經較高溫培養7 d后轉入適宜溫度下的萌發情況
Table 3 Germination ofF.mandshuricaseeds with different stratification and drying treatments at appropriate temperature after 7 d of higher temperature culture
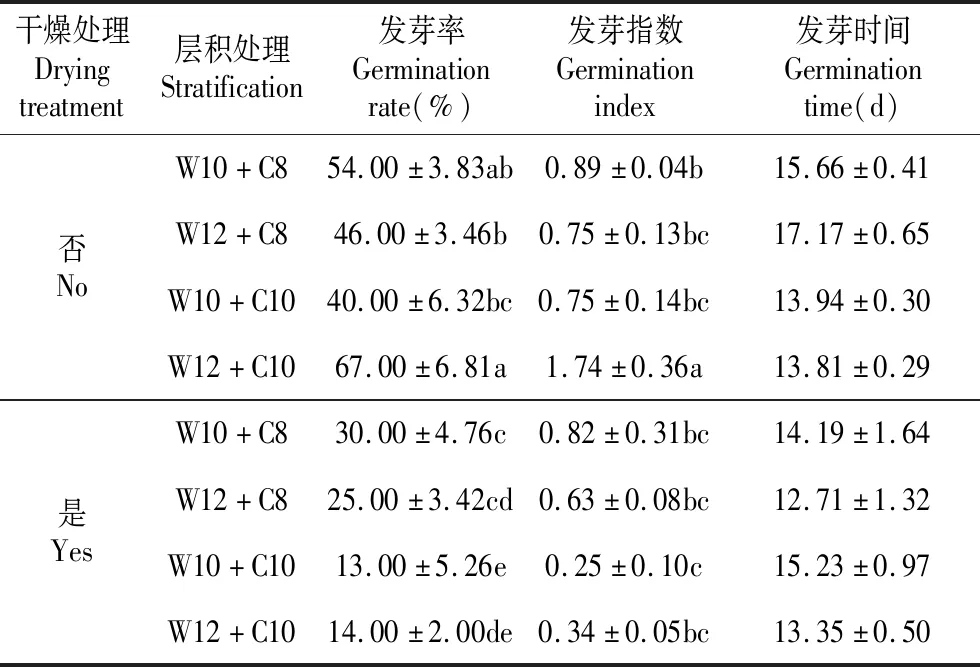
干燥處理Dryingtreatment層積處理Stratification發芽率Germinationrate(%)發芽指數Germinationindex發芽時間Germinationtime(d)否NoW10+C854.00±3.83ab0.89±0.04b15.66±0.41W12+C846.00±3.46b0.75±0.13bc17.17±0.65W10+C1040.00±6.32bc0.75±0.14bc13.94±0.30W12+C1067.00±6.81a1.74±0.36a13.81±0.29是YesW10+C830.00±4.76c0.82±0.31bc14.19±1.64W12+C825.00±3.42cd0.63±0.08bc12.71±1.32W10+C1013.00±5.26e0.25±0.10c15.23±0.97W12+C1014.00±2.00de0.34±0.05bc13.35±0.50
3 討論
本研究發現,萌發溫度為適宜溫度或較高溫條件下,干燥處理對水曲柳種子發芽率、發芽指數的影響均極顯著,表明在不同溫度下種子的萌發會受到干燥處理的影響。萌發溫度為適宜溫度或較高溫條件下,不經干燥處理的種子中,層積時間最長(22周)的種子發芽率、發芽指數最高,發芽時間最短,表明層積時間越長種子的萌發能力就越強,種子初生休眠的解除程度越充分。種子的休眠與萌發受植物激素含量調控,其中脫落酸(ABA)具有誘導、維持種子休眠和抑制種子萌發的作用[11~12],而赤霉素(GA)具有解除種子休眠的作用,并且通過拮抗ABA促進種子萌發[3,13]。有研究表明水曲柳種子休眠與種子中存在長鏈脂肪酸(亞油酸、油酸)等天然發芽抑制物有關[14],今后的研究中可以通過測定種子初生休眠的解除過程中激素、抑制物含量變化來闡釋種子初生休眠的解除程度與萌發之間的關系。經過干燥處理,層積時間越長的種子發芽率下降越明顯,低溫層積時間較長(10周)的種子發芽率下降幅度大于低溫層積時間較短(8周)的種子。由此,可以回答本研究提出的第一個問題,初生休眠解除狀態不同的水曲柳種子在不同溫度下的萌發表現具有相似的規律,種子的萌發都會受到干燥處理的影響。不經干燥處理的種子解除休眠越充分,其萌發能力就越強,但層積處理后的種子若經過干燥處理,則解除休眠越充分(尤其是低溫時間越長)種子萌發能力下降越多。
本研究發現,萌發溫度為適宜溫度時,干燥處理前后,種子發芽率平均下降約11%,發芽指數平均下降約34%;萌發溫度為較高溫時,干燥處理前后,種子發芽率平均下降約60%,發芽指數平均下降約50%;萌發溫度為適宜溫度或較高溫條件下,層積時間最長的種子發芽率、發芽指數下降幅度最大。由此可以回答本研究提出的第二個問題,干燥處理會降低水曲柳種子的萌發能力,尤其是較高溫條件下的萌發能力,初生休眠解除越充分的種子萌發受干燥處理影響越大。而以往研究發現解除休眠的水曲柳種子經室溫干燥處理后發芽率和發芽時間與干燥前相比沒有明顯變化,該試驗材料為20℃層積3個月后轉入5℃層積3個月的水曲柳種子[15],本研究與之產生不同結果的原因可能是初生休眠解除狀態不同的水曲柳種子的萌發能力對干燥處理的響應不同。
有研究表明擬南芥種子次生休眠的誘導受種子初生休眠狀態的影響[16],本研究發現,未干燥的種子經過較高溫培養后,各層積處理的種子萌發表現不同,表明初生休眠解除狀態不同的水曲柳種子的萌發能力對熱休眠的響應不同,即水曲柳種子熱休眠的誘導同樣受種子初生休眠狀態的影響。同時,本研究發現,萌發溫度為較高溫條件下,層積處理顯著影響水曲柳種子的發芽率,但對發芽指數沒有顯著影響;而干燥處理極顯著地影響種子的發芽率和發芽指數,表明水曲柳種子次生休眠(熱休眠)的誘導受種子初生休眠解除狀態的影響較小,受干燥處理影響較大。
種子脫水干燥是一個復雜的生理生化過程,有研究表明低溫或快速脫水會引起水曲柳種子萌發能力下降,其原因是細胞膜透性和丙二醛含量增加,種子中可溶性蛋白含量和胚中抗氧化酶活性降低,從而引起了種子萌發能力下降[17]。還有研究表明許多種子脫水耐性與植物抗氧化系統(抗氧化酶、非酶類抗氧化物質)[18~20]、可溶性糖(蔗糖、棉子糖、水蘇糖、寡糖等)[21~23]、蛋白質(發育晚期豐富蛋白、熱休克蛋白、油脂性蛋白等)[23~25]、分子調控網絡[26]等有關,還有研究發現綠豆種子脫水耐性與程序性細胞死亡(PCD)有關[27]。初生休眠解除狀態不同的水曲柳種子的萌發能力對干燥處理響應的生理機制可以從以上幾個方面進行深入研究。有研究表明適宜濃度的氯化鈣、ABA和蔗糖預處理能夠提高水曲柳種子低溫脫水后的萌發能力[28],在今后的研究中可以與本研究結果相結合,以期探索出最佳的種子脫水干燥處理方法。
綜上所述,初生休眠解除狀態不同的水曲柳種子在不同溫度下的萌發表現具有相似的規律,種子的萌發都會受到干燥處理的影響。不經干燥處理的種子解除休眠越充分,其萌發能力就越強,但層積處理后的種子若經過干燥處理,則解除休眠越充分(尤其是低溫時間越長),種子萌發能力下降越多。水曲柳種子次生休眠(熱休眠)的誘導受種子初生休眠解除狀態的影響較小,但受干燥處理影響較大。干燥處理會降低水曲柳種子的萌發能力,尤其是較高溫條件下的萌發能力,初生休眠解除越充分的種子萌發受干燥處理影響越大。生產中如需對解除休眠的種子干燥貯藏,選擇低溫層積時間較短的處理為好,若從節省層積時間的角度考慮,建議選擇暖溫10周+低溫8周的層積方法。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
