豬MyoG基因外顯子1的多態性分析
2020-07-13 13:39:16孫奴奴
運城學院學報 2020年3期
關鍵詞:實驗
孫奴奴,劉 瑾
(運城學院 生命科學系,山西 運城 044000)
我國畜牧業中養豬業占主導地位,全國豬肉產量不斷地增加,在我國肉類總生產量中占了很大的比例。消費者對豬肉肉質的要求提高,不僅對豬肉的風味、嫩度有要求,對豬肉的營養價值也有更高的要求。國內外對于如何改善豬肉肉質做過大量研究,其中發現可以通過對肌肉生長的調節,改善豬肉品質。而MyoG基因控制肌細胞的增殖與分化,可以增加肌肉的生長速度。肌細胞生成素(MyoG)是生肌調節因子家族中的一員[1],在生肌調節因子家族中只有MyoG基因可以在所有骨骼肌細胞系中表達,它在胚胎發育中控制中胚層干細胞分化定向形成肌細胞,以促進肌纖維的形成,從而控制肌肉的形成[2]。MyoG基因不僅能對自身基因的表達進行調節,還可以與生肌因子家族中MyoD、Myf-5、Myf-6等基因相互作用,對彼此基因的表達進行調節,調節肌肉中肌球蛋白輕鏈基因、肌肉肌酸激酶、肌鈣蛋白等特異基因的表達。所以肌肉的形成、肉質及產量都受到MyoG基因遺傳變異的影響,該基因的遺傳變異直接導致肌肉的變異[3]。
國內外相關研究表明MyoG基因調節肌細胞分化成為肌纖維,與肉質的生成有一定的聯系。劉梅等用PCR-RFLP技術對申農Ι號豬的MyoG基因的兩個多態性位點(3’端和第二內含子內)進行分析,研究發現AA基因型和MN基因型分別為兩個位點的優勢基因型[4]。在滇南小耳豬MyoG基因多態性分析中發現2041,2047這兩個位點的多態性對背膘厚、pH24有顯著影響,對瘦肉率有極顯著影響;1655,1688這兩個位點多態性對瘦肉率和pH24有顯著影響,對背膘厚有極顯著影響[5]。程廣龍等[6]對長淮豬、大淮豬、杜淮豬和淮豬的MyoG基因進行研究,結果顯示胴體質量、瘦肉率、眼肌面積等性狀與股二頭肌的肌纖直徑分別呈顯著正相關。高勤學等[7]對申農I號豬MyoG基因進行多態性分析,發現具有NN基因型的豬初生重顯著高于其他基因型豬,并且具有不同基因型的豬,它的半腱肌和半膜肌的肌纖維密度差異顯著,其中NN基因型肌纖維密度高于NM和MM基因型。TePas等[8]用PCR-RFLP技術對MyoG基因的多態性進行了檢測,結果顯示有3個突變位點:其中,一個是在實驗中研究的所有豬種共同特有的,一個是杜洛克豬和長白豬特有的,另一個是梅山豬特有的。實驗結果表明豬初生重、肌纖維數與這三個變異位點有一定的關聯。Huges等人的研究中發現,MyoD和MyoG基因在肌纖維中的表達不同,MyoD基因在快肌纖維中的表達量比較高,而MyoG基因在慢肌纖維中的表達量比較高。因此,MyoG基因與肉質的風味和嫩度有關聯[9]。另外在其他物種的MyoG基因也有類似研究,雞MyoG基因可以調節肌纖維的形成,對邊雞的生長有影響[10]。對牦牛的MyoG基因進行多態性分析,發現有4個突變位點與牦牛的體高有顯著影響[11]。對6個綿羊群體進行多態性分析,發現含有三種基因型,MyoG基因對羊肉的水分和色澤有影響,在水分的表達中BB>AB>AA;在色澤的表達中AB>BB>AA[12]。
本實驗選用晉汾白豬和新山西黑豬作為實驗動物,采用PCR-SSCP的方法檢測MyoG基因多態性并判定基因型,利用統計分析軟件計算出基因頻率和基因型頻率,分析其基因型在兩豬種中的分布狀況,為研究MyoG基因對晉汾白豬、新山西黑豬肉質影響提供一些材料,為人工選育優良豬種提供依據,對如何生產出優質豬肉有重要指導意義。
1. 材料與方法
1.1 實驗動物
本實驗選用山西省運城市新龍豐畜牧有限公司提供的40頭晉汾白豬和40頭新山西黑豬,分別采集兩個豬種的豬耳組織1.0g,放入裝有75%無水乙醇的離心管中并標號,放入冰盒中帶回實驗室,在-20℃冰箱保存。
1.2 主要試劑與儀器設備
PCR mix,瓊脂糖,溴酚藍,冰醋酸,EB,甘油,DNA Marker,丙烯酰胺,過硫酸銨,亞甲叉雙丙烯酰胺,硼酸,Tris base TEMED等(購自北京華越洋生物工程有限公司);引物合成(上海生物工程技術服務有限公司);PCT-200PCR儀(BIO-RAD公司);DYCZ-28D雙板夾芯式垂直電泳儀,DYY-6C恒溫恒壓電泳儀(北京六一生物科技有限公司);Tanon-3500數碼凝膠圖像分析系統(武漢愛斯佩科學儀器有限公司);TGL-16M高速臺式冷凍離心機(湖南湘儀實驗室儀器開發有限公司)。
1.3 試驗方法
1.3.1 基因組DNA提取及引物合成
使用酚/氯仿/異戊醇/的方法提取豬基因組DNA;根據Genebank發表的豬MyoG基因序列(U14331)并參考郭云雁[13]等人的實驗進行設計引物,并由上海生工生物工程技術服務有限公司合成。
1.3.2 PCR及SSCP
以DNA為模板進行PCR擴增,10微升反應體系為:DNA(0.5微升),上游引物(0.2微升),下游引物(0.2微升),2×Taq PCR Master Mix(5微升),雙蒸水(4.1微升);PCR反應程序為:預變性95℃,5min,94℃變性30s,50℃退火30s,72℃延伸30s,35個循環,72℃后延伸5min;PCR產物取其中的2微升用1.5%的瓊脂糖凝膠電泳檢測,然后將檢測成功的PCR產物,取4微升與6微升6×Loading Buffer混勻,在98℃條件下變性10 min,迅速冰鎮10 min后上樣,在12%的聚丙烯酰胺凝膠中400 V,預電泳10 min,然后將電壓調為200 V,電泳16 h,然后銀染20min,顯影10~20min;最后利用凝膠成像系統拍照并判別基因型。
1.3.3 數據的統計處理
基因型結果利用PopGen32軟件進行群體遺傳學分析,并利用SAS8.1統計軟件進行卡方檢驗。
寨卡病毒感染可導致嚴重的神經系統并發癥,開發疫苗保護易感人群至關重要。最近兩種DNA疫苗(VRC5288質粒和VRC5283質粒)已進入臨床I期試驗,通過對最后一次接種4周后的血樣分析發現,60%~89%接種VRC5288的受試者及77%~100%接種VRC5283的受試者產生了中和抗體應答,并且該疫苗在試驗中具有良好的耐受性及安全性,該試驗鼓舞了DNA疫苗的研究,并加快了預防性疫苗的實際應用[41]。
2. 結果與分析
2.1 PCR產物瓊脂糖凝膠電泳結果
利用晉汾白豬、新山西黑豬的DNA和MyoG引物進行PCR擴增,然后在1.5%的瓊脂糖凝膠上點樣,對PCR產物進行檢測,結果如圖1所示。
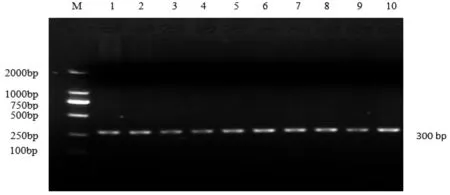
圖1 PCR產物檢測結果(注:M為DL20000 maker,1~10為擴增結果)
如圖1所示,MyoG基因PCR擴增產物長度為300 bp,條帶明亮清晰,沒有雜帶,可進行后續實驗。
2.2 PCR-SSCP結果
用12%聚丙烯酰胺凝膠電泳對晉汾白豬和新山西黑豬的PCR產物進行SSCP檢測,結果如圖2和圖3所示。
如圖2所示,電泳條帶比較清晰,MyoG基因在晉汾白豬中存在多態性,基因型為:1~7和11為AB型,8、10為AA型,9為BB型。
如圖3所示,電泳條帶比較清晰,MyoG基因在新山西黑豬中存在多態性,基因型為:3、4、6、9、11為AB型,5、7、10為AA型,1、2、8為BB型。

圖2 晉汾白豬PCR-SSCP結果(注:1~7、11為AB型,8、10為AA型,9為BB型)

圖3 新山西黑豬PCR-SSCP結果(注:1、2、8為BB型,3、4、6、9、11為AB型,5、7、10為AA型)
2.3 群體遺傳學分析
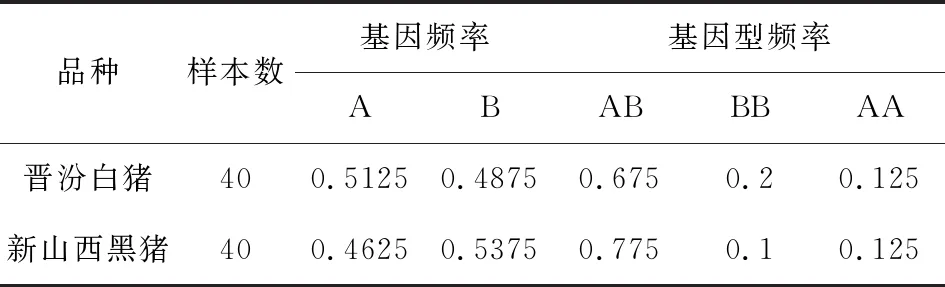
表1 MyoG基因在兩個豬種的基因頻率和基因型頻率
根據表1結果顯示,晉汾白豬中有三種基因型,其中AB有31個,BB有4個,AA有5個;新山西黑豬中有三種基因型,其中AB有27個,BB有8個,AA有5個;在晉汾白豬中A等位基因為優勢等位基因,AA基因型為優勢基因型。說明AA基因型對于晉汾白豬的進化具有促進作用,在人工選育中具有AA基因型的豬更具有經濟效益。新山西黑豬中B等位基因為優勢等位基因,BB基因型為優勢基因型。說明在新山西黑豬中具有BB基因型的豬種在進化的時候更具有優勢,在人工選育中具有BB基因型的豬更具有經濟效益。
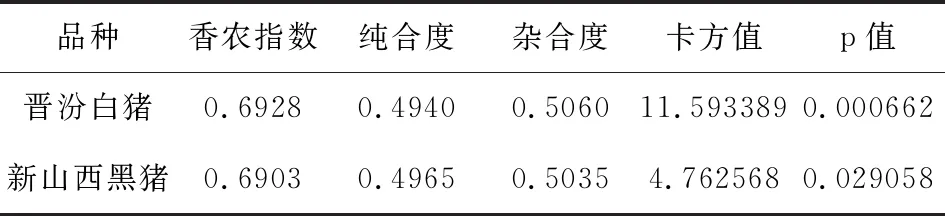
表2 晉汾白豬、新山西黑豬遺傳分析結果
根據表2結果顯示,MyoG基因在晉汾白豬中卡方值為11.593389,p值為0.000662;MyoG基因在新山西黑豬中卡方值為4.762568,p值為0.029058,P值均小于0.05,所以在晉汾白豬、新山西黑豬中MyoG基因的多態性位點均不符合Hardy-Weinberg平衡。晉汾白豬的香農指數(0.6928)大于新山西黑豬的香農指數(0.6903),說明晉汾白豬的遺傳多態性高于新山西黑豬。晉汾白豬的遺傳純合度(0.4940)小于新山西黑豬的遺傳純合度(0.4965),說明新山西黑豬更能穩定遺傳。
2.4 基因型分布差異分析
利用SAS8.1分析軟件,對MyoG基因在晉汾白豬、新山西黑豬中基因型分布進行檢驗,結果如表3所示。
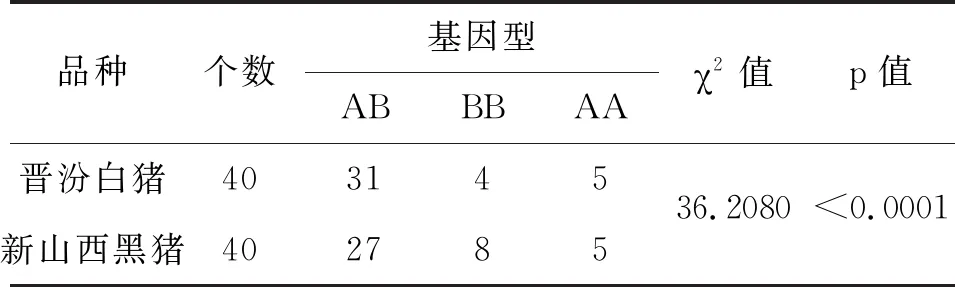
表3 兩個豬種中MyoG基因型分布差異性檢驗
根據表3結果顯示,χ2值為36.2080,p<0.0001。說明在晉汾白豬和新山西黑豬中MyoG基因型分布差異極顯著。
3. 討論與結論
本實驗選用了40頭晉汾白豬和40頭新山西黑豬作為樣本,采用PCR-SSCP的技術對MyoG基因進行多態性分析。結果顯示MyoG基因在晉汾白豬中有三種基因型:AB、BB、AA,A基因為優勢等位基因,在新山西黑豬中有三種基因型:AB、BB、AA,B基因為優勢等位基因。MyoG基因在晉汾白豬中和新山西黑豬中的p值均小于0.05,表明MyoG基因的多態性位點在晉汾白豬、新山西黑豬中都不符合Hardy-Weinberg平衡。對比郭云雁[13]等人的實驗,以180頭大白豬和170頭北京黑豬作為實驗動物,在北京黑豬與大白豬中A基因均為優勢等位基因,在北京黑豬中基因型AB出現的頻率最高,基因型BB出現的頻率最低;在大白豬種中純合基因型AA的頻率最高,而BB基因型的頻率最低,并且MyoG基因在這兩個豬種中都處于Hardy-Weinberg平衡。在王立辛[14]等的實驗中選用255頭大白豬、390頭長白豬、155頭杜洛克豬,三個豬種共800頭作為實驗動物,結果顯示在這三個豬種中均存在三種基因型:AB、BB、AA,其中AB出現的頻率最高,并且B基因均為優勢等位基因。群體遺傳學分析結果顯示MyoG基因多態性位點在大白豬、長白豬和杜洛克豬中均處于Hardy-Weinberg平衡。
本實驗與郭云雁、程篤學和王立辛、蘇玉虹等的研究結果相比,基因頻率、基因型頻率以及Hardy-Weinberg平衡不一致,原因可能有:本實驗選用晉汾白豬和新山西黑豬為樣本,而郭云雁等人選用了大白豬和北京黑豬,王立辛、蘇玉虹等的實驗中選用大白豬、長白豬、杜洛克豬作為試驗動物,實驗中選用的豬種不同,生活的環境、飼養方式、人工選育方法不同,其遺傳性能也會有差異;選用的豬品種經過人工選育時人為擇優選育也有可能會影響基因型頻率,從而影響Hardy-Weinberg平衡。對比郭云雁等人和王立辛、蘇玉虹等人的實驗,本實驗選取的樣品數量太少,本實驗選取晉汾白豬和新山西黑豬樣本各40個,雖符合統計學樣本選取的最低要求,但是誤差比較大,容易出現特異性。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
