溫度對高氮氣調中米象各蟲態致死時間和發育的影響
2020-07-07 02:00:26黃依林王殿軒朱毓斌唐培安
中國糧油學報 2020年6期
黃依林 王殿軒 朱毓斌 唐培安
(河南工業大學糧油食品學院;糧食儲藏與安全教育部工程研究中心;糧食儲運國家工程實驗室1,鄭州 450001)
(江蘇省現代糧食流通與安全協同創新中心2,南京 210023)
氮氣氣調殺蟲長期受到關注,國內外關于氮氣氣調的研究[1-6]和應用[7-10]多有報道。在我國利用制氮設備制氮氣調應用較多的是控溫氣調,相對于常溫條件下的氣調殺蟲效果報道,控制較低溫度下氣調致死害蟲的時間研究為數尚少。通常采用高濃度氮氣致死害蟲需要較長的時間[11, 12],在長期保持氮氣濃度的密閉過程中需要倉房具有較高的氣密性,或倉房氣密性不足時需補充氮氣氣體。不斷或多次補充氮氣需要更多的氣源,或運轉制氮設備消耗大量能量。在一個密閉殺蟲周期內如果目標害蟲仍有存活則為氣調殺蟲失敗,有效掌握氣調中的密閉時間以做到在一個處理過程中完全殺死害蟲尤其重要。
氣調殺蟲效果受諸多因素影響,環境氣密性、害蟲種類和蟲態、環境溫度和濕度、氣體濃度及保持時間等均會影響到處理效果[11,13-17]。同樣氣調過程中溫度不同致死害蟲的時間變化很大[18]。在常溫條件下保持98%以上的氮氣濃度也需要很長的密閉時間[19],環境溫度降低時氣調殺蟲效果相應降低。我國低溫或控溫儲糧已得到了較大規模應用[20],但在低溫儲糧過程中依然會有害蟲發生[21]。基于溫度低時會延遲氣調殺蟲效果[18,22],明確低溫對氣調殺蟲的影響程度,或同樣氣調濃度下低溫對隱蔽性害蟲致死時間的延遲程度很有必要,而此方面的研究尚少。米象Sitophilusoryzae(L.)是世界性分布的主要儲糧害蟲,是我國儲糧主要害蟲之一[23],其在我國分布范圍近年來呈擴大態勢[24],在儲糧害蟲綜合治理方面一直備受重視。米象成蟲和幼蟲均可嚴重蛀食原糧,其非成蟲蟲態卵、幼蟲和蛹均在糧粒內部生活[25],通常方法檢查其發生及處理后的死亡效果較為困難。害蟲不同蟲態對氣調處理的耐受力又存在顯著差異[14, 26],氮氣氣調對米象不同蟲態的效果差異程度缺乏研究。本文研究了98%氮氣濃度下不同溫度時米象卵、幼蟲、蛹和成蟲的致死時間,以期為氮氣氣調殺蟲的有效掌握密閉時間提供量化參考。
1 材料與方法
1.1 試蟲
實驗用米象采自中央儲備糧南寧直屬庫有限公司,后在河南工業大學儲藏物昆蟲研究室適宜條件下培養數代,參照FAO推薦方法測定其磷化氫抗性倍數為1.2倍。
1.2 氣調裝置
采用長為67 cm、寬為52 cm、高為41 cm的食品級PP塑料密封箱作為氣調室,用3 mm 厚的有機玻璃板封蓋。箱體一側密封連接直徑100 mm、長200 mm 圓柱管制成的取蟲緩沖倉,倉的兩頭分別以螺旋蓋密封,可以通過另一側開孔安裝的密封操作手套拿取氣調室中的蟲籠等,以盡量減小氣調過程中從氣調室取樣給氣體濃度造成的影響。密封操作手套安裝孔直徑150 mm,密封連接橡膠手套。氣調室兩側分別設進氣口與閥門、出氣口與閥門,分別通過導管連接氣泵和固定式氮氣檢測儀,檢測儀型號為MOT500 — LM(N2),測量范圍0%~100%,精度0.1%。氣調室可連接鋼瓶裝氮氣氣源,以及相應的減壓閥。氮氣檢測儀可即時檢測室內氮氣濃度,并可促進氣體分布均勻。處理中充入99.999%的氮氣,通過檢測和補充氣體維持目標濃度在98%。氣調室內用過飽和食鹽水調節相對濕度在(75±5)%,并設溫濕度檢測計以觀察,見圖1。氣調室使用狀態下的氣密性正壓 500 Pa的壓力半衰期為120 s。

圖1氣調裝置設置示意圖
1.3 試蟲準備與處理
成蟲的準備:采用直徑10 mm、長70 mm的兩端通透玻璃管用作試蟲蟲籠,放入50頭羽化14 d的成蟲和2 g水分含量為14%的潔凈小麥,兩端用紗布和橡皮筋封口。
卵的準備:適宜條件下用800 g潔凈小麥感染2 000頭羽化2周的米象成蟲,24 h后移出成蟲得染蟲卵小麥,又24 h后將感染蟲卵小麥用分樣器分成5 g的160等份,按需要數量分別放入前述蟲籠中進行測試。
幼蟲的準備:取分樣后的小麥之一份,每日解剖檢查小麥粒內害蟲發育情況,至籽粒內出現幼蟲后,再2 d后將前述同期小麥按需要數量分別放入前述蟲籠中進行測試。
蛹的準備:如幼蟲解剖糧粒檢查,當糧粒內有化蛹后,將含2日齡蛹的小麥進行氣調處理。方法同上。
以上每個處理設3個重復,并設空白對照。將前述氣調室分別置于18、23和28 ℃的溫度下,放入試蟲籠,控制其相對濕度與密封條件,1 d后調節其中氮氣濃度至98%。之后按計劃于不同時間取出蟲籠,培養檢查存活或死亡結果。成蟲蟲籠置于溫度(30±1) ℃、濕度(75±5)% RH環境中,2周后檢查死亡率,含卵、幼蟲和蛹的蟲籠置于相同的溫濕度條件下培養,至羽化時每日觀察羽化出的成蟲數量,直至連續3 d無新增羽化成蟲為止,非成熟蟲態的樣本均對小麥進行剖粒檢查,其中出現未羽化成蟲、存活的蛹和幼蟲計為活蟲,計算未達到100%死亡樣本中存活個體發育至羽化出成蟲的時間。對照(非氣調)樣品做同樣檢查和處理。
1.4 數據處理方法
使用IBM SPSS Statistics 20和Microsoft Excel 2016軟件對實驗數據進行處理。
2 結果與分析
2.1 不同溫度下米象各蟲態在不同時間的死亡率
圖2顯示,不同溫度下米象卵、幼蟲、蛹和成蟲均表現為溫度低時害蟲死亡率增加緩慢,達到100%死亡率的時間越長。18 ℃時的完全致死時間與23和28 ℃時相比,卵達到100%死亡的時間相應延遲4 d和10 d、幼蟲延遲了4 d和8 d、蛹延遲了6 d和12 d、成蟲延遲了6 d和10 d,溫度越低,延遲時間越長。從完全致死時間相比,不同溫度下米象不同發育階段對高氮缺氧環境耐受力的排列順序均為:蛹>卵>幼蟲>成蟲,即在米象對氮氣氣調的耐受力上成蟲最易于被殺死,其次為卵和幼蟲,蛹最難被殺死。




圖2不同溫度時98%氮氣中米象各蟲態不同時間死亡率
(A:卵;B幼蟲; C:蛹;D:成蟲)
2.2 不同溫度時米象卵的LT50值和LT99值
表1顯示,在98%氮氣濃度下溫度18 ℃時致死米象卵的半數致死時間LT50值為9 d,分別比23 ℃和28 ℃時延遲了3 d和6 d,即在實驗溫度范圍內溫度每降低5 ℃,其LT50值對應延長了3 d。高氮氣濃度氣調下,溫度降低后隱藏于小麥粒內的米象蟲卵半數致死時間顯著延遲。溫度降低使得蟲卵呼吸需求變小[27],高氮低氧環境下對其呼吸抑制影響也變小,從而使半數致死時間延長。在18、23 ℃和28 ℃溫度下,高氮氣調致死米象卵的LT99值相應為24、19 d和11 d,溫度降低后代表完全致死蟲卵時間的LT99值延遲顯著,這同樣與低溫下卵生命代謝水平降低,呼吸抑制影響小因而耐力相對較大有關。比較同溫度下的LT99與LT50值,發現18、23 ℃和28 ℃溫度下LT99值與LT50值的差值分別達15、13 d和8 d,同一溫度下LT99值與LT50值的差異與試蟲卵群體中少數個體有極強耐受力有關,不同溫度下LT99值與LT50值的差異也與蟲卵的生命呼吸代謝水平有關。

表1 不同溫度下98%氮氣致死米象卵的LT50值和LT99值
2.3 不同溫度時米象幼蟲的LT50值和LT99值
從表2可見, 不同溫度下98%氮氣氣調致死米象幼蟲的LT50值和LT99值差異顯著。溫度18 ℃時致死米象幼蟲的LT50值長達8 d,分別比23 ℃和28 ℃時延遲了3 d和5 d。在降溫幅度相同的情況下,致死幼蟲的LT50值與卵的LT50值相同或相近。推測處于取食期的幼蟲活動代謝需氧程度比卵大一些,但在高氮環境中降低溫度對其LT50值的影響表現與卵相似,這可能是由于米象幼蟲為無足式,其活動強度較小而需氧量與卵相近。溫度18 ℃時氣調對幼蟲的LT99值為21 d,比23 ℃和28 ℃時相應延長了5 d和11 d,溫度降低同樣大幅度降低了氣調致死幼蟲的LT99值。比較LT99值和LT50值可以看出,同一溫度下的LT99值明顯大于相應溫度下的LT50值,且這種現象在幼蟲期比卵期更為明顯,原因也在于群體中少數耐力極強的幼蟲個體會導致LT99值顯著增大。

表2 不同溫度下98%氮氣致死米象幼蟲的LT50值和LT99值
2.4 不同溫度時米象蛹的LT50值和LT99值
表3顯示,98%氮氣濃度下18 ℃時致死米象蛹的LT50值達11 d,分別比23 ℃和28 ℃時延遲了5 d和7 d。與卵和幼蟲的結果相比,23 ℃和28 ℃時致死蛹的LT50值與卵和幼蟲的LT50值相近,但溫度18 ℃時對蛹的LT50值明顯增大。顯示出高氮低氧環境中溫度降低至18 ℃后,米象蛹對低氧的耐受力明顯增加,這可能與蛹低溫下呼吸強度和生理代謝率更低[28],溫度降低對蛹的致死效果更慢有關。同樣溫度下,18、23 ℃和28 ℃時致死蛹的LT99值分別比相應的LT50值延長了15、15 d和8 d,呈現出溫度越低氣調致死蛹的LT99值越大,其原因同樣是溫度低時蛹呼吸需求低,少數耐力極強個體導致LT99值更多延長。

表3 不同溫度下98%氮氣致死米象蛹的LT50值和LT99值
2.5 不同溫度時米象成蟲的LT50值和LT99值
表4為98%氮氣中不同溫度下致死米象成蟲的LT50和LT99值。18 ℃溫度時致死米象成蟲的LT50值為7 d,分別比23 ℃和28℃時致死成蟲的LT50值延遲了3 d和4 d。低溫延遲LT50值的程度在23 ℃時與幼蟲和卵的相近而明顯小于蛹,在28 ℃時成蟲的LT50值則明顯小于幼蟲、卵和蛹的LT50值。溫度18 ℃時的LT99值為17 d,分別比23 ℃和28 ℃時延長了6 d和10 d,低溫導致對成蟲的LT99值顯著增大。同樣,在同一溫度下都表現出了對成蟲的LT99值均顯著小于對卵、幼蟲和蛹的LT99值,這都與成蟲取食和運動活動更為旺盛,代謝需氧量更多,在高氮低氧環境中更易于被致死有關。
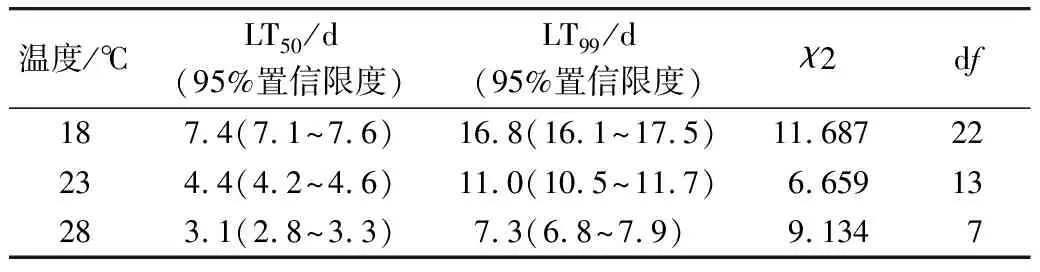
表4 不同溫度下98%氮氣致死米象成蟲的LT50值和LT99值
2.6 米象非成蟲態發育為成蟲的時間
表5顯示,非氣調環境中從蛹羽化出成蟲的時間在23 ℃和28 ℃時均為3 d,18 ℃時接近4 d。在98%氮氣中7 d后再脫離氣調環境,23 ℃和28 ℃時蛹發育至羽化為成蟲時間分別為7 d和8 d,18 ℃時則達10 d。僅氣調7 d的時間再回到正常氣體環境中,23 ℃和28 ℃下蛹至羽化出成蟲的時間延遲了3~4 d,18 ℃下則相應延遲了7 ~ 8 d,低溫18 ℃下導致短期氣調環境中害蟲發育延遲異常明顯。在98%氮氣中殘存蛹發育為成蟲的時間在18、23和28 ℃下分別為27、20 d和14 d,氣調時間延長而未到完全致死害蟲時間,其存活的蛹發育時間延遲更加顯著。98%的氮氣殺蟲濃度下未致死的蛹可發育羽化出成蟲,溫度降低蛹的發育歷期更加延遲。
非氣調環境中28 ℃溫度下從幼蟲發育至羽化出成蟲需要14 d,之后溫度每降低5 ℃幼蟲發育至成蟲的歷期相應延遲2 ~ 3 d,溫度對幼蟲發育影響顯著。98%氮氣氣調7 d后在溫度28 ℃時幼蟲發育至成蟲延長至32 d,溫度降至23 ℃和18 ℃幼蟲發育至成蟲相應延遲2 d和4 d,高氮殺蟲濃度延遲幼蟲發育期明顯,溫度降低幼蟲在短期氣調后發育期延遲也同樣顯著。長期98%氮氣氣調未至完全死亡,其存活的幼蟲在溫度28 ℃時發育至羽化出成蟲時間延長到了39 d,23 ℃和18 ℃相應延遲了3 d和8 d。同樣長期氣調殺蟲濃度下存活幼蟲發育至羽化為成蟲的時間更加延遲,且溫度降低對該發育期的延遲更加明顯。
米象在正常氣體中28 ℃溫度下從卵發育至羽化出成蟲經歷了25 d,之后溫度每降低5 ℃卵發育至成蟲的歷期也相應延遲了3 d以上,這種延遲包括了低溫對卵期的延長,以及對幼蟲和蛹發育期的延長。98%氮氣氣調7 d后卵在28 ℃溫度發育至成蟲需要46 d,比正常氣體環境中相應延遲了21 d,23 ℃和18 ℃溫度下相比較28 ℃分別延遲了2 d和5 d。98%氮氣濃度中完全致死時間前殘存的卵在28 ℃時發育至羽化出成蟲經歷了48 d,23 ℃和18 ℃時相應的歷期也分別延遲了6 d和11 d,溫度對卵至成蟲發育歷期的影響程度與幼蟲期和蛹期相似。
總體上,98%高氮缺氧環境中米象隱蔽蟲態在接近死亡前如果脫離氣調環境,許多個體可以存活,并且可發育羽化出成蟲。此過程中溫度的降低可顯著延遲害蟲的發育歷期。

表5 非氣調和氣調不同時間的非成蟲態米象
3 討論與結論
氣調殺蟲中溫度影響明顯,害蟲致死時間隨溫度降低而延長[22]。在高濃度CO2(90%~95%)和低氧(0.7%~2.1%)環境中,34 ℃溫度下經過10 d米象和玉米象成蟲可完全死亡,22 ℃溫度下則要經過26 d的時間方可完全死亡[25]。關于氣調中溫度較低時對成蟲的殺蟲效果也有所降低[18, 22, 28, 29],氣調中低溫對隱蔽性蟲態的致死效果與影響報道少見。本研究說明98%氮氣環境中米象不同蟲態間的致死時間差異顯著,成蟲對此環境的耐受力最小,氣調中為了保障完全致死害蟲以致成功,應充分考慮處理過程對耐受力最強蟲態的完全致死效果。鞘翅目害蟲如谷象、米象、煙草甲和藥材甲等的蛹期對氣調的忍耐性最強,其次是卵和幼蟲,多表現為成蟲對氣調最敏感[9,12,14]。本研究表明,在18、23 ℃和28 ℃各個溫度條件下,米象對氣調的忍耐力強弱均大體可排序為:蛹>卵>幼蟲>成蟲,實際氣調殺蟲效果評價中應更加關注對米象蛹的致死效果。
適宜溫區內溫度升高害蟲發育加快,呼吸需求大,氣調殺蟲環境下也易于死亡。在實際儲糧中低溫環境中也會有害蟲發生,此時因呼吸代謝相對小,氣調致死害蟲的時間會相應延遲,但具體延遲時間的程度報道很少,尤其氣調對隱蔽蟲態的延遲程度更是關系完全殺蟲的效果。本研究結果提示,在98%的氮氣殺蟲濃度中實現完全殺蟲,對于米象(以對蛹的LT99值為參考)溫度23 ℃時要比28℃延長10 d以上,18 ℃時要比23 ℃時延長4 d以上,18 ℃時要比28 ℃延長14 d以上。實際生產環境中,糧堆溫度常為不均勻分布,從保證完全殺蟲效果考慮,應根據環境中最低溫度或最低生蟲部位的溫度保持氣調密閉時間。
在 25 ℃和70%R H的條件下采用98%~100%的氮氣濃度進行氣調處理,19 d才可完全抑制玉米象的卵、蛹和幼蟲,11 d可以完全致死谷蠹成蟲,但要抑制其非成蟲蟲態則需要延長至20 d[12]。溫度28 ℃下2%穩定氧氣濃度條件下90 d對米象種群抑制率為99. 94%[17],說明缺氧殺蟲環境中米象種群中有些個體可以長時間存活。缺氧條件下長時間種群抑制不能達100%的原因與米象隱蔽蟲態耐氣調性較大有關,更與氣調過程中存活個體發育繁殖造成種群世代重疊,后代隱蔽蟲態耐力表現有關。本研究表明,98%高氮缺氧氣調時間不足時,米象隱蔽蟲態在未被殺死的情況下脫離氣調環境可以存活并發育至成蟲羽化,此過程中溫度的降低可以延遲氣調環境下害蟲的發育歷期。基于此結果,在氣調殺蟲中對于保持殺蟲氣體濃度的時間可以參考正常氣體環境下的昆蟲發育參數,但更要在相應發育歷期參數基礎上延長密閉氣調時間,環境溫度更低時延長時間的幅度更大。
