超表達StCYS1對馬鈴薯生長發育及酶促褐變的影響
2020-06-29 12:09:02劉敏敏李廣存劉士揚董田田王慶國
農學學報 2020年6期
劉敏敏,李廣存,劉士揚,董田田,王慶國,劉 佩
(1山東農業大學食品科學與工程學院/山東省高校食品加工技術與質量控制重點實驗室,山東泰安 271018;2中國農業科學院蔬菜花卉研究所,北京 100081)
0 引言
蛋白酶抑制子(proteinase inhibitors,PIs)是生物體內發揮重要作用的多肽和蛋白質,廣泛存在于動植物和微生物中。半胱氨酸蛋白酶抑制子(cystatin,CYS)是PIs 家族的一個重要分支,通過抑制半胱氨酸蛋白酶(cysteine proteinase,CP)的活性參與多種生理生化過程[1]。相關研究表明,CYS 能夠調控植物的生長發育,如貯藏蛋白的沉積和動員、衰老器官中蛋白質的分解代謝、維管組織的分化、葉片衰老、種子萌發和病原菌誘導的細胞程序性死亡等,也參與植物對環境脅迫的應答[2-4]。
CYS 在作物生長發育中具有重要作用。過表達海棠MpCYS4基因的轉基因蘋果減緩了干旱脅迫下葉片凈光合速率的下降,并阻止了CP對葉片中蛋白質的降解,從而減緩了蘋果葉片的衰老[5]。異源表達水稻CYS 基因oryzacystatin-I或用人工合成的CYS E64 處理增加了擬南芥的側根密度[6]。CYS通常局限于植物發育的特定階段如果實成熟期表達,影響果實產量[7]。馬鈴薯中CYS的研究主要集中于對環境脅迫的應答和調控方面,目前關于CYS是否能直接對馬鈴薯的生長發育起調控作用,CYS對馬鈴薯植株的光合作用過程及根系發育影響的研究未見報道。
酶促褐變是鮮切馬鈴薯貯藏、加工過程中的難題。前人研究結果表明較高水平PIs的表達具有抑制馬鈴薯褐變的作用,外施PIs 類似物處理馬鈴薯漿液后褐變顯著減輕[8]。劉博文[9]研究表明,預切傷處理馬鈴薯導致Kunitz型PIs基因表達量上調,總蛋白酶活性下降,游離酪氨酸含量下降,褐變顯著減輕。但是,目前關于PIs抑制馬鈴薯褐變的調控途徑尚不清楚。
本文通過測定超表達CYS 基因StCYS1和野生型二倍體馬鈴薯植株的光合作用、根系發育情況及結薯量;以超表達株系和野生型株系的塊莖為材料,對馬鈴薯塊莖的褐變表型進行觀察,對PPO 活性、總抗氧化能力及游離氨基酸含量進行測定,旨在探究StCYS1基因對馬鈴薯生長發育的作用,揭示StCYS1對馬鈴薯褐變調控的生理機制,旨在為StCYS1調控馬鈴薯生長發育及酶促褐變方面提供分子基礎和理論依據。
1 材料與方法
1.1 材料
試驗所用的馬鈴薯(Solanum tuberosum)材料為二倍體‘MDS’,由中國農業科學院蔬菜花卉研究所李廣存提供。超表達StCYS1基因(NCBI 登錄號:No.MG793453.1)二倍體馬鈴薯株系OE1、OE2、OE3 是將StCYS1基因連接pBI121 超表達載體后,通過農桿菌AGL1+VirG介導轉化MDS所得,由本實驗室保存。
1.2 試驗設計
試驗于2018年8—12月在山東農業大學食品科學與工程學院的植物組織培養室內進行。組培室內晝/夜溫度為21~23℃/21~23℃,16 h光照/8 h黑暗,相對濕度為45%~55%。馬鈴薯野生型株系‘MDS’(WT)和超表達StCYS1株系OE1~OE3 試管苗在MS 培養基中生長1個月后,分別選取長勢相近的各株系試管苗30株栽植于盆(規格為A350;直徑31.2 cm;高19.0 cm)中,營養土:蛭石=2:1,每盆一株。待植株生長60 天后,觀察其生長狀態,取樣測定光合指標及根系發育情況。待植株生長120天后,采收各株系馬鈴薯,用于統計產量并測定馬鈴薯塊莖褐變情況、PPO活性、總抗氧化能力(FRAP抑制率,ABTS、DPPH自由基清除率)和游離氨基酸含量。
1.3 主要儀器
GXZ 光照培養箱,寧波江南儀器廠;熒光定量PCR 儀,美國Bio-Rad 公司;CIRAS-3 便攜式光合儀,英國PP System公司;Win RHIZO植物根系分析儀,石家莊泛勝科技有限公司;紫外分光光度計T6 新世紀,北京普析通用儀器有限責任公司;TGL16高速冷凍離心機,長沙英泰有限公司;DW-86L 388A超低溫冰箱,海爾集團;超聲波振蕩器,上海生析超聲儀器有限公司;液氮研磨儀,德國IKA 公司;高效液相色譜儀,日本島津LC-20AT;C18液相色譜柱,Thermo Fisher公司。
1.4 試驗方法
1.4.1StCYS1基因表達量的測定 取WT 和OE1~OE3株系葉片及塊莖測定StCYS1表達量。采用OminiPlant RNA Kit(DNase I)全能型植物RNA提取試劑盒(康為世紀)提取樣品RNA,cDNA 利用反轉錄試劑盒(HiFiScript cDNA Synthesis Kit,康為世紀)獲得。qRT-PCR采用UItraSYBR Mixture試劑盒(康為世紀),操作參照說明書進行,所用引物為:正向引物F:5′-TCGTAAGTCGGAGTCGGATTATGGT-3′,反向引物R:5′-TGGAACTTCCTGGTGTCACCAAGTC-3′。Actin基因參考Hemavathi 等[10]。反應程序為:95℃預變性10 min,然后執行40 個循環反應(95℃變性15 s,60℃退火和延伸1 min),最后65~95℃進行溶解曲線分析。所有實時熒光定量PCR反應均設置3次生物學重復和3次技術重復,所得數據采用2-ΔΔCt進行計算。
1.4.2 凈光合速率測定 盆栽苗生長60天后,選取在頂葉下完全展開的第3 片復葉,于上午9:00—11:00 用CIRAS-3便攜式光合儀測定系統(PPSystems,英國)測定凈光合速率(Pn)、胞間二氧化碳濃度(Ci)、蒸騰速率(Tr)和氣孔導度(Gs),各株系分別選取長勢一致的12株苗子測定,取其平均值。
1.4.3 根系形態指標的測定 使用專業版Win RHIZO根系分析儀,對生長60 天馬鈴薯苗根系長度、根系體積和根面積等主要根系參數進行分析。
1.4.4 馬鈴薯產量統計WT 和OE1~OE3 株系采收后的馬鈴薯清洗、擦干后稱重,統計產量。
1.4.5 馬鈴薯褐變感官評價 將WT 和OE1-OE3 株系馬鈴薯的塊莖用液氮研磨儀打粉后,分別稱取2 g 于50 mL離心管中,迅速加入10 mL去離子水,置于常溫下,分別在0、1、2、3 h觀察褐變情況并拍照。
1.4.6 PPO 活性測定 粗酶液提取及PPO 活性的測定參考等Zhou 等[11]的方法并有所改動。取1 mL 10 mmol/L 的鄰苯二酚溶液和1.5 mL 磷酸緩沖溶液,混勻后迅速加入0.75 mL 粗酶液,于420 nm 測定吸光值(每30 s測定一次,測定3 min)。每分鐘內每克樣品吸光值變化0.01為一個酶活力單位[U/(g·min)]。
1.4.7 總抗氧化能力測定ABTS自由基清除率的測定參考Re 等[12]的方法;DPPH 自由基清除率的測定參考Seijo-Rodríguez等[13]的方法;鐵還原能力FRAP抑制率的測定參考羅冬蘭等[14]的方法。
1.4.8 游離氨基酸含量測定 馬鈴薯游離氨基酸的提取參考Kumar 等[15]的方法,采用高效液相色譜法測定16種游離氨基酸(天冬氨酸、酪氨酸、脯氨酸、亮氨酸、甘氨酸、組氨酸、蘇氨酸、纈氨酸、蛋氨酸、異亮氨酸、苯丙氨酸、谷氨酸、絲氨酸、精氨酸、丙氨酸和賴氨酸),方法參考徐鑫等[16]的研究。
1.5 數據分析
采用SPSS 17.0 軟件對數據進行數據統計分析及差異顯著性的檢驗。采用Excel 2010統計分析軟件進行基礎數據整理分析與作圖。所有試驗均重復3次。
2 結果與分析
2.1 超表達和野生型株系StCYS1基因表達量
馬鈴薯植株栽植前,取WT和OE1~OE3葉片進行qRT-PCR 鑒定,超表達株系OE1、OE2 和OE3 葉片StCYS1基因表達量分別是野生型WT 的4.91、6.20 和3.92 倍(圖1A),差異顯著。馬鈴薯結薯后,取WT 和OE1~OE3馬鈴薯塊莖進行qRT-PCR鑒定,超表達株系OE1、OE2 和OE3 塊莖StCYS1基因表達量分別是野生型WT的5.21、5.67和4.17倍(圖1B),差異顯著。測定結果表明,超表達株系StCYS1在馬鈴薯葉片和塊莖中均有過量表達。
2.2 超表達StCYS1對馬鈴薯光合特性的影響
光合作用是影響植物生長的關鍵因素,不僅影響馬鈴薯植株的生長發育,而且直接影響塊莖產量和品質。對野生型和轉基因株系馬鈴薯植株光合參數進行測定,結果表明超表達株系OE1、OE2和OE3的凈光合速率為12.51、13.36、12.03 μmol CO2/(m2·s),比WT 提高了16.31%、24.24%、11.90%,各株系間差異達顯著水平。超表達株系OE1、OE2 和OE3 的胞間CO2濃度比WT 提高了8.00%、11.74%、8.63%,各株系與WT 相比差異顯著。與WT相比,OE1、OE2和OE3各株系的蒸騰速率、氣孔導度測定結果趨勢一致(圖2),分別是WT 的1.29、1.32、1.33 倍和1.23、1.25、1.25 倍,超表達各株系均顯著高于野生型(P<0.05)。研究結果表明超表達StCYS1顯著提高了馬鈴薯植株的光合能力。
2.3 超表達StCYS1對馬鈴薯根系發育的影響
馬鈴薯根系從土壤中吸收水分和營養物質,一方面支撐地上部分的生長,另一方面,根圍繞匍匐莖生長,對馬鈴薯地下塊莖的發育也起著重要作用[17]。組培苗移栽前,WT株系根分生數較OE1-OE3株系少(圖3)。移栽入定植盆60天后測定根系相關參數發現,超表達株系OE1~OE3須根系比野生型WT的根系發達,具體表現為總根長、總表面積和總體積顯著高于野生型(P<0.05)。尤以OE2株系根發育最好,總根長、總表面積、平均直徑和總體積分別為1221.05 cm、292.25 cm2、0.87 mm 和6.87 cm3,是WT 的2.42、1.87、1.34 和3.15倍(表1),差異顯著。由此可知,超表達StCYS1顯著促進了馬鈴薯植株根系的發育。
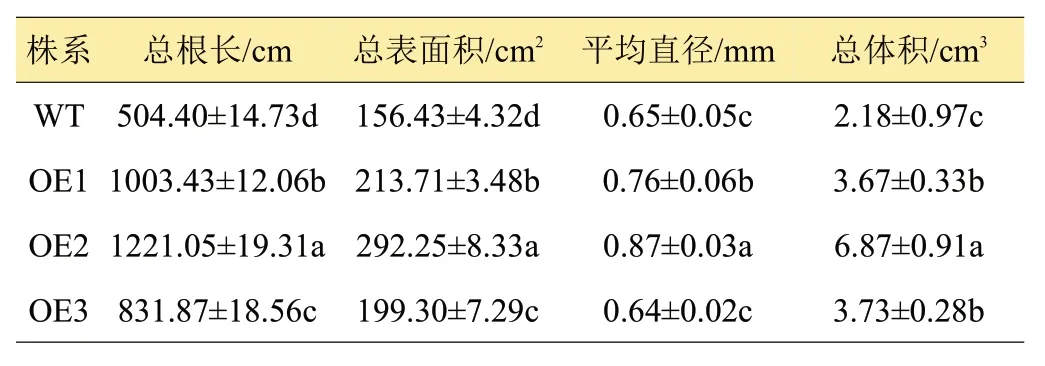
表1 不同馬鈴薯株系根系相關參數
2.4 超表達StCYS1對馬鈴薯產量的影響
馬鈴薯植株的結薯量的統計結果(表2)表明,不同株系馬鈴薯塊莖的產量差異較大,超表達株系OE1~OE3 馬鈴薯單株薯重和單株結薯數量為14.25、19.63、10.93 g 和23.33、17.86、10.33 個,均顯著高于WT(P<0.05);而且OE1~OE3 株系最大單薯重量大于WT,尤以OE2株系的單薯重量最大,達11.48 g。由此可知,超表達StCYS1顯著提高了馬鈴薯塊莖產量。
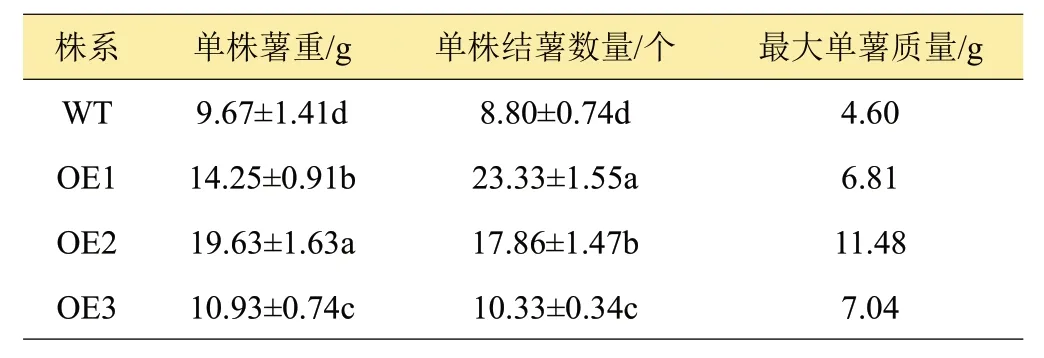
表2 不同株系馬鈴薯產量
2.5 超表達StCYS1對馬鈴薯酶促褐變的影響
OE1~OE3 和WT 株系結薯后,挑選大小相當、無破損的馬鈴薯塊莖去皮、液氮研磨成漿,觀察其漿液褐變情況。如圖4 所示,隨時間變化,WT 漿液褐變程度逐漸加重,OE1~OE3 漿液褐變程度均顯著低于WT,且3 個株系中,StCYS1表達量較高的2 個株系OE1、OE2褐變程度一直低于OE3。結果表明超表達StCYS1可顯著抑制馬鈴薯漿液的褐變。在此基礎上,通過測定WT 和超表達株系塊莖PPO 活性、抗氧化能力及游離氨基酸含量,探討超表達StCYS1對馬鈴薯塊莖褐變的影響機制。
2.6 超表達StCYS1對馬鈴薯PPO活性的影響
多酚氧化酶(PPO)是酶促褐變過程中黑色素合成途徑中的關鍵酶。研究表明通過基因沉默或者敲除手段來抑制馬鈴薯PPO 基因表達,以及通過物理、化學方法抑制PPO 活性,均能抑制塊莖鮮切褐變[18]。由圖5可知,超表達株系OE1~OE3的PPO活性均高于野生型WT。其中,OE1 和OE2 酶活性分別是WT 的1.41和1.36 倍,顯著高于WT(P<0.05),OE3 酶活性是WT的1.08 倍,但無顯著性差異(圖5)。超表達StCYS1的馬鈴薯塊莖漿液褐變程度降低,但PPO 活性卻增高了,結果表明PPO可能并不是影響超表達StCYS1馬鈴薯塊莖褐變的關鍵因素。
2.7 超表達StCYS1對馬鈴薯總抗氧化能力的影響
研究表明,PIs 的積累能夠提高抗氧化能力[19],而在馬鈴薯鮮切褐變控制的研究中發現,塊莖抗氧化能力,如DPPH自由基清除率、FRAP和ABTS自由基清除率等的提高,往往會抑制鮮切褐變[20],因此測定了WT和超表達株系的抗氧化能力。如圖6所示,超表達株系的FRAP抑制率(鐵還原能力)、DPPH自由基清除率和ABTS自由基清除率均顯著高于野生型WT,不同指標的測定結果具有一致性,三者互為印證,即StCYS1基因的超表達顯著提高了馬鈴薯的抗氧化能力。
2.8 超表達StCYS1對馬鈴薯游離氨基酸含量的影響
前人[21]研究發現,果蔬褐變過程中,PPO催化酚類底物氧化而生成鄰醌,游離氨基酸可與醌類物質反應,促進褐變過程或作為螯合劑抑制褐變。而在馬鈴薯鮮切褐變中,游離酪氨酸是褐變不可或缺的底物[22]。經過測定,發現OE1~OE3株系與WT株系存在11種差異氨基酸,其中OE1~OE3株系中游離天冬氨酸、蘇氨酸、脯氨酸、亮氨酸共4 種游離氨基酸含量顯著高于WT,游離酪氨酸、甘氨酸、組氨酸、纈氨酸、甲硫氨酸、異亮氨酸和苯丙氨酸共7 種氨基酸含量顯著低于WT。褐變底物酪氨酸含量的降低,與StCYS1基因超表達降低馬鈴薯鮮切褐變密切相關。而脯氨酸含量的升高,可使得馬鈴薯塊莖抗氧化能力增加,從而抑制鮮切馬鈴薯褐變。
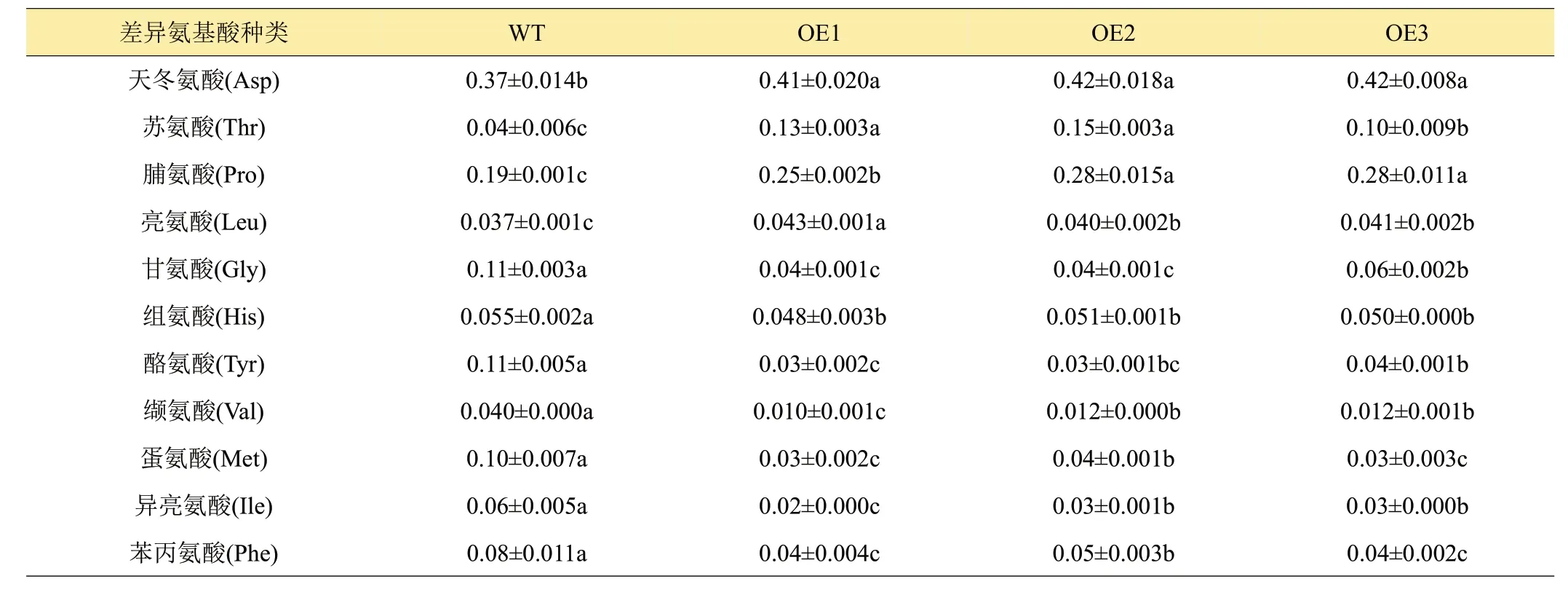
表3 不同株系馬鈴薯差異游離氨基酸含量 g/kg
3 討論
3.1 超表達StCYS1 提高了植株光合能力,促進了根系發育進而提高了馬鈴薯的產量
前人研究發現薯類產量與光合作用的強弱密切相關。Song 等[23]研究合理調整甘薯的種植距離和種植密度能夠提高甘薯的凈光合速率,達到增產的目的。光合作用是決定馬鈴薯產量的最重要的初級代謝過程,干旱脅迫下光合能力較強的馬鈴薯減產較少[24]。本研究發現,StCYS1超表達株系的光合能力提高,產量較野生型顯著增加,這與前人的研究結論一致。植物根系的發育構型與其利用土壤養分、水等資源的能力密切相關,影響植株地上部的生長和果實產量[25]。適度干旱脅迫能誘導馬鈴薯根長和根表面積增加,塊莖產量提高[26]。本研究發現,StCYS1超表達株系的總根長和總表面積與野生型相比顯著增加;光合作用的提高使植株的有機物積累增多,這可能是馬鈴薯產量提高的重要原因。然而,超表達StCYS1是否進一步影響了馬鈴薯塊莖的品質,如淀粉、VC和酚酸含量等,還需要進一步深入研究。
3.2 超表達StCYS1 通過提高馬鈴薯塊莖抗氧化能力、降低游離酪氨酸含量來抑制褐變
植物組織的抗氧化能力對保護細胞膜的完整性起重要作用。馬鈴薯受到鮮切傷害時,細胞膜的完整性被破壞,打破了酚-酚酶的區域分布,PPO 和酚類底物接觸并發生反應,引起鮮切馬鈴薯的褐變[27]。脯氨酸既能夠清除活性氧,減輕細胞的氧化損傷,也能作為重要的滲透調節物質,在細胞膜受損后維持細胞的滲透壓,對保護細胞膜完整性起重要作用。超表達StCYS1馬鈴薯塊莖的脯氨酸含量升高,起到了減輕細胞膜氧化損傷、維護細胞膜穩定性的作用,進而減輕了馬鈴薯的褐變。范昊安等[20]研究發現,蘋果梨酵素通過發酵提高了自身抗氧化活性,有效抑制了酶促褐變。抗氧化能力的強弱也是評價馬鈴薯抗褐變能力的重要指標。本研究結果證明了超表達StCYS1顯著提高了二倍體馬鈴薯的總抗氧化能力,推測StCYS1超表達抑制褐變與增強馬鈴薯的抗氧化酶系統有關。試驗后續需研究SOD、CAT等抗氧化酶的活性及其基因表達的變化,深入探究抗氧化系統與酶促褐變的關系。
在以往的研究中,氨基酸主要作為非酶褐變如美拉德反應的底物、抗壞血酸褐變系統的誘導因子等影響著農產品加工過程中的褐變[28,29]。雖然已有研究證明游離氨基酸尤其是酪氨酸是馬鈴薯酶促褐變的主要底物,但是關于氨基酸參與酶促褐變的機理研究較少。蛋白酶的大量表達會導致蛋白質的水解[15],進而產生大量的游離酪氨酸[30]。超表達StCYS1可能是通過抑制蛋白酶的活性使蛋白質水解減少,游離酪氨酸含量降低,進而抑制了褐變。因此,超表達StCYS1能否影響總蛋白酶活性及其作用的靶蛋白酶是StCYS1抑制馬鈴薯酶促褐變機制的研究方向。
3.3 超表達StCYS1提高了PPO活性,間接促進了馬鈴薯的生長發育
前人研究表明PPO除了在生物體酶促褐變、體內色素合成等過程中起關鍵的作用外,PPO 催化酚類底物需要的氧氣和NADH由光合系統提供,在植物光合作用、生物合成、抵御外界脅迫等生理過程中具有重要作用[31]。姜翠翠等[32]研究發現,?樹的生長發育過程中,PPO 基因在幼嫩組織中有較高的表達量。推測PPO 活性可能與超表達StCYS1馬鈴薯植株的光合作用有關,進而影響了馬鈴薯的生長發育。超表達StCYS1提高了馬鈴薯株系塊莖的抗氧化能力,抑制蛋白酶的水解,減少了褐變底物游離酪氨酸,PPO活性的提高沒有加重馬鈴薯的褐變。因此,還需進一步探究超表達StCYS1馬鈴薯的生長發育與酶促褐變過程中主要作用因子間的相互關系,測定PPO 基因的表達,以更好地揭示StCYS1對馬鈴薯生長發育和酶促褐變的作用機理。
4 結論
本研究結果表明,超表達StCYS1馬鈴薯植株的凈光合速率、根系發育程度和塊莖產量都顯著高于野生型植株,說明StCYS1的超表達促進了馬鈴薯植株的生長發育。此外,與野生型相比,超表達StCYS1馬鈴薯塊莖褐變程度輕,游離脯氨酸含量高,總抗氧化能力強;酪氨酸含量低,PPO活性高,說明StCYS1超表達主要通過增強馬鈴薯塊莖抗氧化能力和降低酪氨酸含量來減輕褐變,PPO不是主要影響因素。
