鹽度脅迫對黃條鰤消化生理和抗應激指標的影響
2020-06-27 13:13:50柳學周曹亞男劉永山徐永江
海洋科學 2020年6期
史 寶, 柳學周, 2, 曹亞男, 劉永山, 2, 徐永江, 姜 燕, 王 濱
鹽度脅迫對黃條鰤消化生理和抗應激指標的影響
史 寶1, 柳學周1, 2, 曹亞男3, 劉永山1, 2, 徐永江1, 姜 燕1, 王 濱1
(1. 青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室 農業農村部海洋漁業可持續發展重點實驗室 中國水產科學研究院黃海水產研究所, 山東 青島 266071; 2. 上海海洋大學 水產與生命學院, 上海 201306; 3. 煙臺市海洋經濟研究院, 山東 煙臺 264000)
為探討鹽度突變對黃條鰤()幼魚消化酶活力和抗應激指標的影響, 設計了采用自然海水養殖的對照組鹽度29(S29)和實驗組鹽度分別為35(S35)、15(S15)、10(S10)和5(S5), 對黃條鰤進行了120 h的急性脅迫實驗, 測定了各鹽度條件下消化酶活力、超氧化物歧化酶(SOD)活力及甲狀腺激素(T4)濃度的變化。結果顯示: 黃條鰤胃、腸、肝臟和幽門盲囊的脂肪酶活力, 6 h時實驗組與對照組差異不顯著(0.05), 12 h后呈現隨時間的增長而活力降低的現象, 且實驗組顯著低于對照組(0.05); 蛋白酶活力脅迫24 h后, 實驗組顯著低于對照組(0.05)。胃和肝臟蛋白酶活力脅迫6~12 h, S35顯著高于對照組(0.05)。胃和腸的淀粉酶活力脅迫后, 實驗組均顯著低于對照組(0.05); 肝臟淀粉酶活力脅迫6~12 h, S35均高于對照組, 24 h后顯著低于對照組(0.05)。S5的SOD活力隨著時間的增加而降低, 且差異顯著(0.05); S15和S35在120 h時SOD活力降低并接近對照組。各鹽度組血清中T4的濃度在6~96 h顯著高于對照組(0.05), 隨后降低并在120 h時趨于穩定。綜上所述, 鹽度脅迫對黃條鰤幼魚消化酶活力、SOD活力和T4濃度影響較大, 黃條鰤對鹽度變化有較強的調節能力, 相關生理指標變化可為黃條鰤養殖提供參考。
黃條鰤; 鹽度突變; 消化酶; 超氧化物歧化酶; 甲狀腺激素
魚類通過自身內部調節系統響應外界環境對其生理功能的影響, 其中包括環境因子對魚類生長相關性狀的調控。鹽度是一個重要的環境因子, 并且與魚類生長、發育、繁殖等生理活動密切相關[1-2]。鹽度通過影響魚類的滲透壓, 間接影響了魚類生長存活與攝食轉化、物質交換與能量流動等相關生理活動[3]。消化酶是由消化系統分泌的具有消化作用的酶類, 其活性可以在一定程度上反映魚類的消化吸收能力[4]。許多無機離子是消化酶的激活劑或抑制劑, 水生生物生活環境中的鹽度變化直接影響其中無機離子的濃度的變化, 進而影響消化酶活性[5]。黃鰭鯛()幼魚的蛋白酶、淀粉酶和脂肪酶活性在鹽度20~30時顯著高于鹽度5~15[6]。同一鹽度下飼養的點籃子魚()各消化器官的脂肪酶酶活性高低順序依次為: 腸道、胃、肝臟和幽門盲囊; 在鹽度5和10條件下的點籃子魚蛋白酶、淀粉酶和脂肪酶活性均顯著低于鹽度20和鹽度30~32組[7]。鹽度也是影響魚類免疫防御活動的主要環境因子, 外界環境鹽度的突然升高或降低都會使魚體產生大量的超氧陰離子自由基(O2–), 對魚體組織細胞等造成氧化損傷。超氧化物歧化酶(SOD)是魚體內抗氧防御系統的關鍵酶, 可通過清除由鹽度變化使魚體產生的過多O2–來保護魚體[8-9]。另外, 魚體內大部分生理反應都會有激素的參與。甲狀腺激素(T4)是一種內分泌激素, 對魚類適應鹽度起到積極的調節作用, 并能促進魚體生長, 是一種重要調節激素[10]。
黃條鰤()屬鱸形目(Percifor-mes)、鲹科(arangidae)、鰤屬(), 廣泛分布于太平洋和大西洋沿岸, 是一種全球性分布的海洋中上層暖溫性遠洋洄游魚類, 并在日本、中國、新西蘭和澳大利亞等地進行了商業養殖[11-12]。黃條鰤生長速度快、個體大, 肉質鮮美, 含有豐富的EPA、DHA和鈣、磷、鐵等微量元素, 具有很高的經濟價值和食用價值,深受廣大消費者喜愛, 世界范圍內消費需求不斷增加, 是我國發展深海魚類養殖的優良品種[13-16]。國內外關于黃條鰤鹽度適應性機理研究鮮有報道[17-19], 本研究以黃條鰤幼魚為實驗材料進行鹽度突變實驗, 通過研究消化酶及SOD活力、T4濃度等指標的變化規律, 初步探討了黃條鰤對鹽度突變的適應性, 從而為深入認識黃條鰤對環境的應激和適應機制提供基礎資料, 并為養殖生產提供理論參考。
1 材料與方法
1.1 實驗材料
實驗在大連富谷水產有限公司進行, 實驗用魚為2013年在黃海北部捕獲的黃條鰤幼魚, 經網箱養殖和室內工廠化越冬, 促熟培育、成功產卵后人工培育的批量苗種, 體長為(20.02±2.07) cm, 體重為(115.50±6.67) g。苗種養殖期間自然海水的水溫為20~27 ℃、鹽度29, 每天投喂鮮雜魚3~4次, 投喂量為魚體重的3%~5%。實驗開始前所有暫養的實驗用魚停食24 h后再放入各鹽度組, 實驗期間不投餌。采用自來水(經24 h以上連續曝氣)并添加自然海水進行低鹽度調節; 高鹽度組的海水使用自然海水加海水晶進行人工配置。
1.2 實驗方法
1.2.1 實驗設計
參照本課題組黃條鰤鹽度突變的實驗方法[17], 設計當前實驗。各實驗組的鹽度調節: 低鹽度組采用自然海水(鹽度29)加自來水調節, 使用的自來水用1 000 L塑料桶作為自來水曝氣池, 連續充氣、曝氣24 h以上使用; 高鹽度實驗組的海水使用自然海水加海水晶進行人工配置后使用。實驗容器為1 000 L實驗桶, 每個鹽度組放入30條黃條鰤幼魚, 用納米充氣石持續充氣保證溶氧充足。設置實驗組鹽度為35(S35)、15(S15)、10(S10)和5(S5), 對照組為自然養殖海水鹽度29(S29), 實驗開始時, 將黃條鰤幼魚直接放入各鹽度組, 持續觀察魚的適應情況, 并按以下時間取各組魚的組織樣品, 取樣時間點為6 h、12 h、24 h、48 h、72 h、96 h、120 h。每次隨機從不同鹽度組中撈取3條魚取樣。每天晚上18: 00換等鹽度海水一次, 每次換水量為50%。實驗期間各實驗組的水質指標控制在水溫23~24℃、pH 7.9~8.1、DO 6~7 mg/L、NH4+-N 0.1~0.3 mg/L。升、降鹽度采取逐漸換水的方式, 鹽度計算公式如下:
當水溫高于17.5 ℃,(‰)= 1 305(– 1) + (– 17.5) × 0.3
式中,代表鹽度;代表比重;代表溫度。
1.2.2 樣品采集與保存
每次在取樣時間點隨機從不同鹽度的實驗桶中撈取3條魚取樣, 用MS222麻醉劑將黃條鰤幼魚麻醉,用無菌1 mL 注射器快速從尾部取血, 置于1.5 mL無菌離心管中, 在冰水混合物中靜置30 min 后, 4℃、5 000 r/min離心10 min, 取上清即為血清。分離出的血清用來檢測SOD和T4。用液氮保存胃、腸、肝臟和幽門盲囊, 用于消化酶活力的測定, 所測消化酶包括脂肪酶、淀粉酶、蛋白酶。
1.2.3 樣品生化指標測定
生化指標的測定參照本課題組黃條鰤鹽度漸變研究中的測定方法[18]。
采用脂肪酶測試盒(A054, 南京建成生物工程研究所)測定了黃條鰤幼魚胃、腸、肝臟和幽門盲囊的脂肪酶活性, 活力單位定義為: 在37℃條件下, 每毫升酶液在反應體系中與底物反應1 min, 每消耗1 mmol底物為1個酶活力單位, 即脂肪酶活力單位為(U/gprot)。
采用蛋白酶測試盒(A080, 南京建成生物工程研究所)測定了黃條鰤幼魚胃、腸、肝臟和幽門盲囊的蛋白酶活性, 其活力單位定義: 每毫升組織液37℃每分鐘分解生成1 g氨基酸相當于1個酶活力單位, 即蛋白酶活力單位為(U/mL)。
采用碘-淀粉比色法(C016, 南京建成生物工程研究所)測定了黃條鰤幼魚胃、腸、肝臟和幽門盲囊的淀粉酶活性, 其活力單位定義: 組織中每毫克蛋白 37℃, 與底物作用30 min, 水解10 mg淀粉定義為1個淀粉酶活力單位, 即淀粉酶活力單位為(U/mgprot)。
使用超氧化物歧化酶(SOD)試劑盒(A001-1, 南京建成生物工程研究所)測定了黃條鰤幼魚血清總SOD含量, 檢測每毫升反應液中SOD抑制率達50%時所對應的SOD量為1個酶活力單位, 即SOD活力單位為(U/mL)。使用Fish T4 ELISA Kit試劑盒(CK-E90004F)測定了黃條鰤血清甲狀腺激素(T4)的含量, T4濃度單位為(ng/mL)。
1.3 統計分析
使用SPSS 17.0軟件的單因素方差分析(One-way ANOVA)和Duncan’s多重比較分析。0.05為差異顯著。作圖數據均以平均值±標準差(Mean±SD)表示。
2 結果
2.1 脂肪酶活力變化
黃條鰤幼魚胃脂肪酶的活力在各實驗組內均隨處理時間的增加而降低, 在6 h之內各實驗組之間無顯著差異(0.05), 隨后持續降低, 96 h之后各實驗組均趨于穩定; 且在整個處理過程中所有實驗鹽度組胃脂肪酶活力均低于對照組(圖1)。
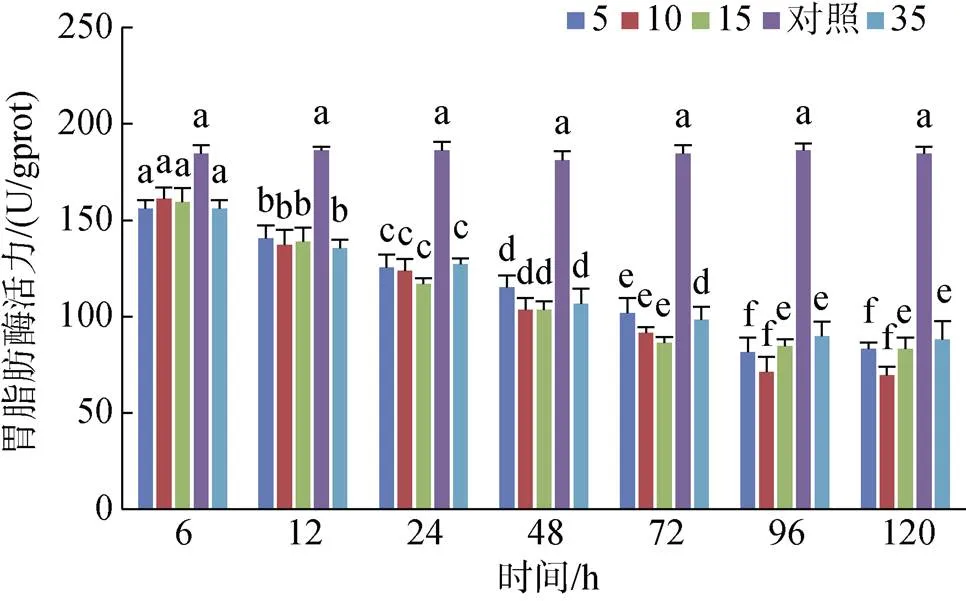
圖1 不同鹽度對胃脂肪酶活力的影響
注: 不同字母表示不同鹽度下胃脂肪酶活力有顯著性差異(0.05)
腸脂肪酶在S10組的活力在6 h時超過對照組鹽度的活力; 隨后各實驗組腸脂肪酶活力持續降低, 均低于對照組; 隨著時間的延長, S5、S15和S35組腸脂肪酶活力整體上呈顯著降低趨勢 (0.05), S5組在48 h之后趨于穩定, S15組在72 h之后趨于穩定, S35組腸脂肪酶活力96 h后趨于穩定(圖2)。
肝臟脂肪酶活力在整個處理過程中各實驗組鹽度下均低于對照組活力, 且隨著時間的增加活力均呈現降低的趨勢, 96 h之后各實驗組肝臟脂肪酶活力趨于穩定(圖3)。

圖2 不同鹽度對腸脂肪酶活力的影響
注: 不同字母表示不同鹽度下腸脂肪酶活力有顯著性差異(0.05)

圖3 不同鹽度對肝臟脂肪酶活力的影響
注: 不同字母表示不同鹽度下肝臟脂肪酶活力有顯著性差異(0.05)
幽門盲囊脂肪酶活力在各實驗組鹽度下均隨時間增加均呈現降低的趨勢, 且均低于對照組; S5、S10和S15組幽門盲囊脂肪酶活力在96 h時趨于穩定, 而S35組則持續降低(圖4)。

圖4 不同鹽度對幽門盲囊脂肪酶活力的影響
注: 不同字母表示不同鹽度下幽門盲囊脂肪酶活力有顯著性差異(0.05)
2.2 蛋白酶活力變化
黃條鰤幼魚胃蛋白酶的活力隨著鹽度的增加而增加。S35組活力在6~12 h時顯著高于對照組(0.05), 之后顯著降低且低于對照組。S5、S10和S15組在整個處理過程中均未超過對照組的活力; 96 h后4個實驗鹽度組的胃蛋白酶活力趨于穩定, 各實驗組與對照組有顯著性差異(0.05)(圖5)。
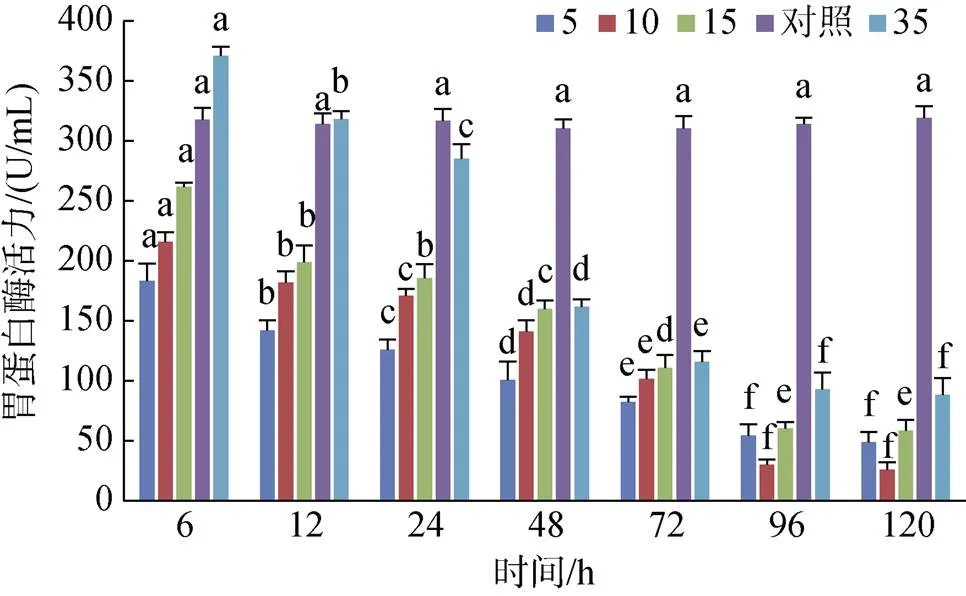
圖5 不同鹽度對胃蛋白酶活力的影響
注: 不同字母表示不同鹽度下胃蛋白酶活力有顯著性差異(0.05)
腸蛋白酶的活力在6 h時, S35組略低于對照組, 其他鹽度組腸蛋白酶活力都處于較低水平, 顯著低于對照組的活力(0.05), 且均隨著時間的增長而降低, 72 h后4個實驗組腸蛋白酶活力均趨于穩定(圖6)。

圖6 不同鹽度對腸蛋白酶活力的影響
注: 不同字母表示不同鹽度下腸蛋白酶活力有顯著性差異(0.05)
肝臟蛋白酶活力在6 h和12 h時S35組活力均高于對照組; 6~48 h各實驗鹽度組活力持續降低, 與對照組有顯著性差異(0.05), 96 h之后趨于穩定(圖7)。

圖7 不同鹽度對肝臟蛋白酶活力的影響
注:不同字母表示不同鹽度下肝臟蛋白酶活力有顯著性差異(0.05)
S15和S35組幽門盲囊蛋白酶活力在6 h時高于對照組, 之后各實驗鹽度組幽門盲囊蛋白酶活力隨著時間的增長而降低, S5組72 h時幽門盲囊蛋白酶活力趨于穩定, 而S10、S15和S35組則在96 h后趨于穩定, 均與對照組有顯著性差異(0.05)(圖8)。

圖8 不同鹽度對幽門盲囊蛋白酶活力的影響
注: 不同字母表示不同鹽度下幽門盲囊蛋白酶活力有顯著性差異(0.05)
2.3 淀粉酶活力變化
4個實驗組黃條鰤幼魚胃淀粉酶活力在整個處理過程中均未超過對照組的活力, 且整體隨時間的增加呈降低的趨勢, S10組胃淀粉酶活力隨著時間的增加上下波動; S15組胃淀粉酶活力持續降低, S5和S35組則在96 h后趨于穩定(圖9)。
腸淀粉酶活力在實驗組中也未超過對照組的活力, 總體隨時間增加呈下降的趨勢, S5和S35組腸淀粉酶活力持續降低, 而S10和S15組則在96 h后趨于穩定(圖10)。
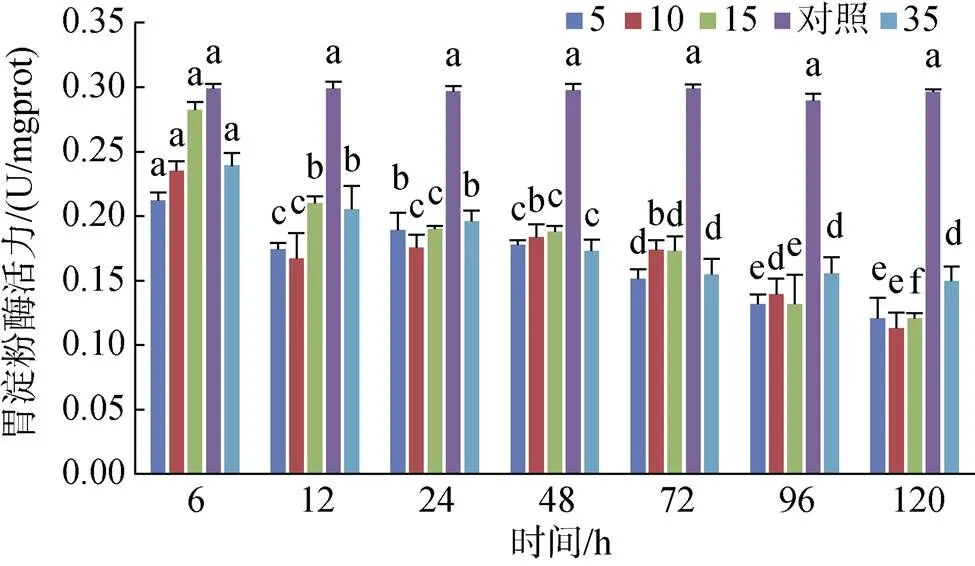
圖9 不同鹽度對胃淀粉酶活力的影響
注: 不同字母表示不同鹽度下胃淀粉酶活力有顯著性差異(0.05)

圖10 不同鹽度對腸淀粉酶活力的影響
注: 不同字母表示不同鹽度下腸淀粉酶活力有顯著性差異(0.05)
4個實驗組肝臟淀粉酶活力在6 h和12 h時活力均高于對照組, 且S35組明顯高于其他組(0.05); 4個實驗鹽度組肝臟淀粉酶活力24 h之后則均低于對照組活力, 且各實驗組鹽度下肝臟淀粉酶活力持續降低, 各組間差異顯著(0.05)(圖11)。

圖11 不同鹽度對肝臟淀粉酶活力的影響
注: 不同字母表示不同鹽度下肝臟淀粉酶活力有顯著性差異(0.05)
幽門盲囊淀粉酶活力在S5、S10和S15組均低于對照組, 且隨時間增長呈降低趨勢, 其中鹽度10組在72 h時趨于穩定; S35組在6 h和12 h時活力顯著高于對照組(0.05), 24 h和48 h基本和對照組持平, 隨后降低且低于對照組, 96 h后趨于穩定(圖12)。
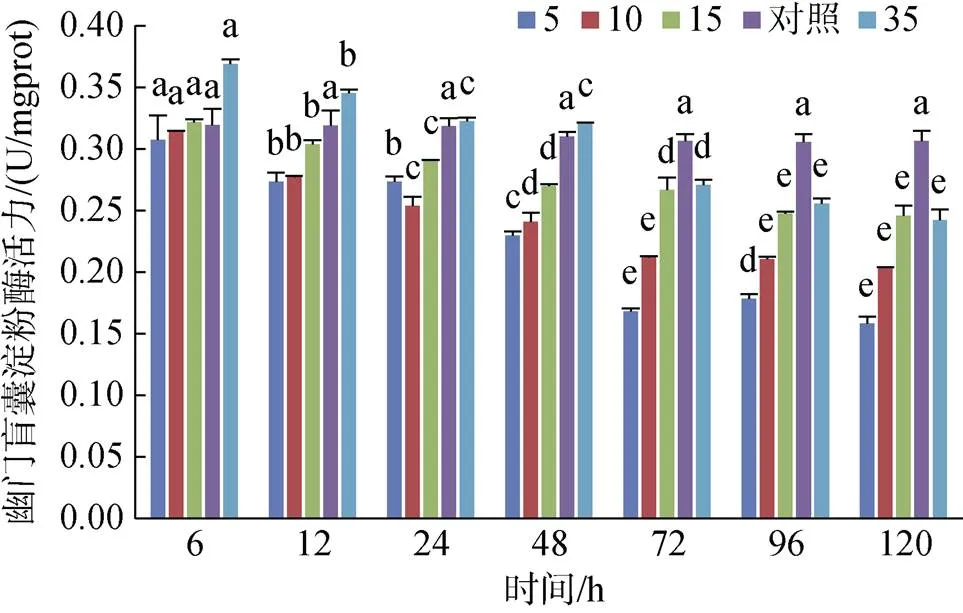
圖12 不同鹽度對幽門盲囊淀粉酶活力的影響
注: 不同字母表示不同鹽度下幽門盲囊淀粉酶活力有顯著性差異(0.05)
2.4 超氧化物歧化酶活力變化
S5組隨著時間的增加, 黃條鰤幼魚血清SOD活力逐漸降低, 120 h時顯著低于其他鹽度組(0.05); S10組SOD活力隨著時間的增加逐漸升高, 48~96 h之間無顯著性差異(0.05); S15組SOD活力在96 h之前一直升高, 且各組間差異顯著(0.05), 120 h時略有降低, 與96 h時SOD活力有顯著性差異(0.05); S35組SOD活力在6 h和12 h顯著低于其他鹽度組(0.05), 24 h之后顯著升高(0.05), 48 h之后逐漸穩定, 48~96 h組間無顯著性差異(0.05), 120 h時SOD活力降低, 接近對照組SOD活力(圖13)。

圖13 不同鹽度對血清超氧化物歧化酶活力的影響
注: 不同字母表示不同鹽度下血清超氧化物歧化酶活力有顯著性差異(0.05)
2.5 甲狀腺激素濃度變化
黃條鰤在不同鹽度海水中, 經過6~120 h后其血清中甲狀腺激素(T4)濃度變化情況。96 h之前各實驗組黃條鰤幼魚血清T4濃度顯著高于對照組(0.05), 隨著時間的增加濃度逐漸降低。S5組T4濃度在6 h和12 h間無顯著性差異(0.05), 120 h急劇下降, 與96 h時T4濃度有顯著性差異(0.05)。S10組T4濃度持續降低, 在12 h時降低明顯, 之后降低幅度變小, 96 h后保持穩定。S15和S35組T4濃度均隨時間的增加而降低, 96 h后趨于穩定。各實驗組T4濃度均在96 h時降低到較低水平, 隨后趨近對照組濃度(圖14)。

圖14 不同鹽度對血清中甲狀腺素濃度的影響
注; 不同字母表示不同鹽度下血清甲狀腺激素濃度有顯著性差異(0.05)
3 討論
3.1 鹽度突變下黃條消化酶活力的變化規律
消化酶是影響營養吸收轉化的關鍵酶類, 鹽度對魚類消化酶活性有重要的影響[20]。水體的鹽度通過影響魚類消化酶活性來影響對餌料的消化和吸收, 最終影響魚類的生長發育[21]。有研究表明, 鹽度在一定范圍內的變化導致魚體消化道內消化酶活力的變化, 可大致歸結為3類: 第一類是激活消化酶的作用[22], 第二類是抑制消化酶的作用[23], 第三類是對消化酶作用沒有明顯影響[24]。張龍崗等[25]對高體革鯻()的研究結果表明在0~13鹽度下隨著鹽度的升高脂肪酶活力受到的抑制逐漸降低, 在鹽度超過13后高體革鯻脂肪酶活力開始上升, 脂肪酶活力又受到激活。但是在本研究中黃條鰤幼魚消化吸收相關的各組織脂肪酶活力并未出現上升情況, 這可能是由于魚的種類以及鹽度設置的不同導致消化系統脂肪酶活力變化的不同。在本研究中黃條鰤幼魚胃、腸、肝臟和幽門盲囊的脂肪酶活力在6 h之內與對照組無顯著差異(0.05), 12 h之后隨著處理時間的增長脂肪酶活力降低, 顯著低于對照組, 推測可能是由于鹽度變化導致了pH等變化, 超出了脂肪酶的適應范圍, 進而受到抑制。
大馬哈魚()的研究結果表明適當增加鹽度能夠促進胃蛋白酶活性, 推測其原因是由于氯離子是蛋白酶的激活劑[22]。與該研究類似, 在本研究中高鹽度S35組時黃條鰤幼魚6 h之前胃、肝臟和幽門盲囊的蛋白酶活力顯著高于其他鹽度組, 但隨時間增長蛋白酶活力顯著降低; 推測可能是由于Na+、K+、Cl–等離子濃度升高, 導致對黃條鰤幼魚消化系統蛋白酶的激活作用下降。在點籃子魚研究中發現, 鹽度5和鹽度10條件下第24 d和第48 d時點籃子魚蛋白酶活力均顯著低于鹽度30~32(對照)組[7]。本研究中, 低鹽度S5、S10和S15實驗組黃條鰤蛋白酶活力也均顯著低于S29(對照)組, 可能是因為低鹽度脅迫下黃條鰤無法通過消化酶活性的調節進而維持正常的消化功能。
在花鰻鱺()和太平洋雙色鰻鱺()的研究中顯示胃、腸淀粉酶活力隨著鹽度增加而下降[4]。在本研究中黃條鰤幼魚胃、腸、肝臟和幽門盲囊淀粉酶活力在S35組隨著時間的延長均明顯下降, 實驗結束120 h時顯著低于對照組淀粉酶活力, 說明較高鹽度對黃條鰤幼魚淀粉酶活力具有一定的抑制作用, 筆者認為, 這可能與鹽度變化產生的O2–清除不及時影響了淀粉酶的活性有關。黃條鰤幼魚消化器官中蛋白酶、脂肪酶和淀粉酶活力在120 h時最高的均為對照組, 不同鹽度海水對黃條鰤幼魚各消化器官中消化酶活力均有顯著性影響, 但各鹽度組酶活性在96 h后基本趨于穩定, 說明黃條鰤幼魚已經慢慢適應環境, 具有較強的調節能力。
3.2 鹽度突變下黃條SOD活力的變化規律
SOD活力是魚體內自由基的代謝情況的重要指標, 反映了魚體的應激能力[26]。環境鹽度的變化可使魚體產生大量的超氧陰離子自由基(O2–), SOD能夠將超氧陰離子自由基(O2–)轉化成H2O2, 接著由過氧化氫酶將產生的H2O2進一步處理轉變為H2O和O2, 保護機體細胞內環境穩定[27]; SOD活力變化可以準確反映魚體內自由基的代謝情況, 對判斷魚的健康狀況具有重要意義[28]。在鹽度突變的過程中, 許氏平鲉()幼魚血液中SOD活力在略低的5和15鹽度條件下被激活以消除機體中過多的O2–自由基, 從而保護機體內細胞免受其傷害[29]。在本研究中S10組黃條鰤幼魚血清SOD活力持續升高, 且顯著高于對照組; S15組SOD活力24 h后均顯著高于對照組, 僅在120 h時略有降低; 說明黃條鰤受到低鹽度環境脅迫時, 魚體進行滲透調節, 代謝加快, 體內產生大量的活性氧自由基, SOD活力升高以清除產生的活性氧自由基。本研究中S5組SOD活力顯著低于其他鹽度組, 且持續降低, 可能是由于鹽度5對黃條鰤幼魚來說屬于重度的低鹽度脅迫, 產生的大量自由基在體內積累并對實驗魚造成氧化損傷, 導致魚體內自由基代謝紊亂, 其SOD活力被抑制, 不能進行正常的抗氧化調節。
3.3 鹽度突變下黃條T4濃度的變化規律
在魚體中, 甲狀腺激素具有多效性, 它與皮質醇和生長激素有類似的作用, 均參與調控魚類的生長、發育、新陳代謝和滲透調節[30]。水體鹽度的突變會導致金頭鯛()血漿中T4濃度的變化, 引發甲狀腺機能亢進, 進而激活腦垂體、鰓和腎等組織器官參與調節[10], 因此T4濃度變化可以反應魚體對外界環境變化的應激能力。該研究顯示金頭鯛在低鹽度5和15組, 血漿中T4濃度顯著高于鹽度40組[10]。本研究中在低鹽度組黃條鰤幼魚血清T4濃度也顯著高于對照組, 其濃度的增加表明T4參與了滲透調節, 以適應鹽度的劇烈變化, 有助于增加黃條鰤對低鹽度變化的適應性。對淡水龜殼攀鱸()的研究發現在高鹽度20組血漿中T4濃度顯著升高, 在該鹽度條件下養殖3周后, 血漿中T4濃度恢復到對照組水平[31]。本研究結果與之相似, 6~24 h高鹽度S35組黃條鰤幼魚血清T4濃度顯著高于對照組, 說明黃條鰤在鹽度提升后, 通過增加T4含量進而增強其對高鹽度海水的耐受性, 96 h之后逐漸穩定接近對照組, 說明黃條鰤對激素的調控作用依懶性降低, 因此分泌的T4減少。
[1] 楊板, 趙文, 魏杰, 等. 鹽度對透明溞存活、生長和繁殖的影響[J]. 水產科學, 2019, 38(3): 361-367.Yang Ban, Zhao Wen, Wei Jie, et al. Effects of salinity on survival, growth and reproduction in water fleas(Crastacea: Cladocera)[J]. Fisheries Science, 2019, 38(3): 361-367.
[2] Lein I, Tveite B, Gjerde B, et al. Effects of salinity on yolk sac larvae of Atlantic halibut (L)[J]. Aquaculture, 1997, 156: 291-303.
[3] De Silva S, Perera P. Studies on the young grey mullet,L.: I. Effects of salinity on food intake, growth and food conversion[J]. Aquaculture, 1976, 7(4): 327-338.
[4] 羅鳴鐘, 關瑞章, 靳恒. 鹽度對花鰻鱺和太平洋雙色鰻鱺幼鰻生長性能及消化酶活力的影響[J]. 水生生物學報, 2015, 39(4): 653-660. Luo Mingzhong, Guan Ruizhang, Jin Heng. Effects of the salinity on the growth performance and digestive enzyme activities ofelver andelver[J]. Acta Hydrobiologica Sinica, 2015, 39(4): 653-660.
[5] 沈永龍, 戈賢平, 黃金田, 等. 鹽度對瘤背石磺()消化酶活性的影響[J]. 動物營養學報, 2012, 24 (9): 1839-1846. Shen Yonglong, Ge Xianping, Huang Jintian, et al. Effects of salinity on digestive enzyme activities of[J]. Chinese Journal Animal Nutrition, 2012, 24(9): 1839-1846.
[6] 李希國, 李加兒, 區又君. 鹽度對黃鰭鯛幼魚消化酶活性的影響及消化酶活性的晝夜變化[J]. 海洋水產研究, 2006, 27(1): 40-45.Li Xiguo, Li Jiaer, Ou Youjun. Effects of salinity on digestive enzyme activity and diurnal variation of digestive enzyme activity of young yellowfin black porgy[J]. Progress in Fishery Sciences, 2006, 27(1): 40-45.
[7] 羅集光, 章龍珍, 莊平, 等. 鹽度對點籃子魚消化酶活性的影響[J]. 海洋漁業, 2011, 33(1): 33-38.Luo Jiguang, Zhang Longzhen, Zhuang Ping, et al. Effects of salinity on digestive enzyme activity in[J]. Marine Fisheries, 2011, 33(1): 33-38.
[8] Sinha A K, Elgawad H A, Zinta G, et al. Nutritional status as the key modulator of antioxidant responses induced by high environmental ammonia and salinity stress in European Sea Bass ()[J]. PLoS ONE, 2015, 10(8): e0135091.
[9] Sui Y M, Huang X Z, Kong H, et al. Physiological responses to salinity increase in blood parrotfish (♀♂)[J]. Springer---Plus, 2016, 1246.
[10] Ruiz-Jarabo I, Klaren P H M, Louro B, et al. Characteri-za-tion of the peripheral thyroid system ofacclimated to different ambient salinities[J]. Comparative Biochemistry and Physiology, Part A, 2017, 203: 24-31.
[11] Chai X L, Li X X, Lu R M, et al. Karyotype analysis of the yellowtail kingfish(Perci-formes: Carangidae) from South Australia[J]. Aquculture Research, 2009, 40(15): 1735-1741.
[12] 劉靜, 陳詠霞, 馬琳. 黃渤海魚類圖志[M]. 北京: 科學出版社, 2015: 172.Liu Jing, Cheng Yongxia, Ma Lin. Fishes of the Bohai Sea and Yellow Sea[M]. Beijing: Science Press, 2015: 172
[13] 史寶, 柳學周, 劉永山, 等. 黃條鰤線粒體全基因組測定及結構特征分析[J]. 中國水產科學, 2019, 26(3): 405-415. Shi Bao, Liu Xuezhou, Liu Yongshan, et al. Complete sequence and gene organization of the mitochondrial genome of[J]. Journal of Fishery Sciences of China, 2019, 26(3): 405-415.
[14] 柳學周, 徐永江, 李榮, 等. 黃條鰤()肌肉營養組成分析與評價[J]. 漁業科學進展, 2017, 38(1): 128-135.Liu Xuezhou, Xu Yongjiang, Li Rong, et al. Analysis and evaluation of nutritional composition of the muscle of yellowtail kingfish ()[J]. Progress in Fishery Sciences, 2017, 38(1): 128-135.
[15] 史寶, 劉永山, 柳學周, 等. 黃條鰤()染色體核型分析[J]. 漁業科學進展, 2017, 38(1): 136-141.Shi Bao, Liu Yongshan, Liu Xuezhou, et al. Study on the karyotype of yellowtail kingfish ()[J]. Progress in Fishery Sciences, 2017, 38(1): 136-141.
[16] 韓羽嘉, 田甲申, 李多慧, 等. 黃條鰤不同組織碳穩定同位素的轉化率與分餾[J]. 水產學雜志, 2017, 30(6): 34-40.Han Yujia, Tian Jiashen, Li Duohui, et al. Conservation rate and fractionation of carbon stable isotope from different tissues in yellow tail[J]. Chinese Journal of Fisheries, 2017, 30(6): 34-40.
[17] 柳學周, 史寶, 劉永山, 等. 鹽度突變對黃條鰤幼魚滲透調節功能的影響[J]. 大連海洋大學學報, 2019, 34(6): 767-775. Liu Xuezhou, Shi Bao, Liu Yongshan, et al. Effects of sharp changes in salinity on osmotic regulation func-tion in juvenile yellowtail kingfish[J]. Journal of Dalian Ocean University, 2019, 34(6): 767-775.
[18] 史寶, 劉永山, 柳學周, 等. 鹽度漸變對黃條鰤消化酶和超氧化物歧化酶活力及甲狀腺激素的影響[J]. 中國海洋大學學報(自然科學版), 2020, 50(1): 48-56.Shi Bao, Liu Yongshan, Liu Xuezhou, et al. Effects of gradual salinity on the digestive enzyme activity, super-oxide dismutase activity and thyroid hormone content of yellowtail kingfish ()[J]. Periodical of Ocean University of China, 2020, 50(1): 48-56.
[19] 史寶, 劉永山, 柳學周, 等. 鹽度漸變過程對黃條鰤()幼魚滲透調節的影響[J]. 海岸工程, 2019, 38(1): 63-70.Shi Bao, Liu Yongshan, Liu Xuezhou, et al. Effects of gradual salinity change on osmotic regulation of juvenile yellowtail kingfish ()[J]. Coastal Engineering, 2019, 38(1): 63-70.
[20] Gheisvandi N, Hajimoradloo A, Ghorbani R, et al. The effects of gradual or abrupt changes in salinity on digestive enzymes activity of Caspian kutum(Kamensky, 1901) larvae[J]. Journal of Applied Ichthyology, 2015, 31(6): 1107-1112.
[21] 田宏杰, 莊平, 高露姣. 生態因子對魚類消化酶活力影響的研究進展[J]. 海洋漁業, 2006, 28(2): 158-162.Tian Hongjie, Zhuang Ping, Gao Lujiao. Advances on the studies of the effect of ecological factors on activities of digestive enzymes of fish[J]. Marine Fisheries, 2006, 28(2): 158-162.
[22] Sánchez-Chiang L, Cisternas E, Ponce O. Partial purification of pepsins from adult and juvenile salmon fish. Effect of NaCl on proteolytic activities[J]. Comparative Biochemistry and Physiology Part B, 1987, 87(4): 793-797.
[23] 林浩然. 魚類生理學[M]. 廣州: 廣東高等教育出版社, 2011: 213-219. Lin Haoran. Fish Physiology[M]. Guangzhou: Guang-dong Higher Education Press, 2011, 213-219.
[24] Fang L S, Chiou S F. Effect of salinity on the activities of digestive protease from the Tilapia fish,in different culture environments[J]. Compara-tive Biochemistry and Physiology, Part A, 1989, 93: 439- 443.
[25] 張龍崗, 安麗, 孫棟, 等. 鹽度脅迫對高體革鯻幼魚消化酶活力的影響[J]. 水產學雜志, 2011, 24(3): 21-24. Zhang Longgang, An Li, Sun Dong, et al. Effects of salinity on digestive enzyme activities of juvenile Jade Perch[J]. Chinese Journal of Fisheries, 2011, 24(3): 21-24.
[26] 楊靜雯, 周演根, 黃銘, 等. 鹽度對虹鱒和硬頭鱒幼魚消化酶和抗氧化酶活性的比較研究[J]. 中國海洋大學學報(自然科學版), 2019, 49(3): 119-128.Yang Jingwen, Zhou Yangen, Huang Ming, et al. Comparative studies on digestive and antioxidant enzyme activities between juvenile Rainbow () and Steelhead Trout ()[J]. Periodical of Ocean University of China, 2019, 49(3): 119-128.
[27] 王妤, 莊平, 章龍珍, 等. 鹽度對點籃子魚的存活、生長及抗氧化防御系統的影響[J]. 水產學報, 2011, 135(1) : 66-73.Wang Yu, Zhuang Ping, Zhang Longzhen, et al. Effects of salinity on survival, growth and antioxidant defense system of[J]. Journal of Fisheries of China, 2011, 35(1): 66-73.
[28] Martínez R M, Morales A E, Sanz A. Antioxidant defenses in fish: biotic and abiotic factors[J]. Reviews in Fish Biology and Fisheries, 2005, 15(1-2): 75-88.
[29] 王曉杰, 張秀梅, 李文濤. 鹽度脅迫對許氏平鲉血液免疫酶活力的影響[J]. 海洋水產研究, 2005, 26 (6): 17-21. Wang Xiaojie, Zhang Xiumei, Li Wentao. Effect of salinity stress on the non-specific immuno-enzymetic activity of[J]. Marine Fisheries Research, 2005, 26(6): 17-21.
[30] Peyghan R, Enayati A, Sabzevarizadeh M. Effect of salinity level on TSH and thyroid hormones ofgrass carp,[J]. Veterinary Research Forum, 2013, 4 (3): 175-178.
[31] Rejitha V, Peter V S, Subhash M C. Short-term salinity acclimation demands thyroid hormone action in the climbing perchBloch[J]. Journal of Endocrinology and Reproduction, 2009, 13(2): 63-72.
Effects of salinity stress on the digestive physiology and anti-stress index of Yellowtail Kingfish ()
SHI Bao1, LIU Xue-zhou1, 2, CAO Ya-nan3, LIU Yong-shan1, 2, XU Yong-jiang1, JIANG Yan1, WANG Bin1
(1. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China; 2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China; 3.Yantai marine economic research institute, Yantai 264000, China)
In this study, we assessed the influence of salinity mutation on the digestive enzyme activity and antistress index of juvenile yellowtail kingfish()Five salinity levels (29, 35, 15, 10, and 5, abbreviated as S29, S35, S15, S10, and S5, respectively) were designed. Here, S29 is natural seawater and is regarded as the control group.The effects of abrupt changes in salinityon the digestive enzymes, superoxide dismutase (SOD), and thyroid hormone (T4) of juvenile yellowtail kingfishwere measured without salinity acclimation after transfer to S29, S35, S15, S10, and S5 during the 120-h breeding experiment. The results showed that the lipase activity in the gastric, intestinal, hepatic, and pyloric follicles in the experimental group were not significantly different from those corresponding to S29 in 6 h (>0.05). The lipase activity decreased with time after 12 h for S35, S15, S10, S5, and it was significantly lower than that for S29 (<0.05). The protease activity of the gastric, intestinal, hepatic, and pyloric follicles in the experimental groupwas significantly lower than that in S29 after 24 h (<0.05). The protease activity in stomach and liver in S35 group was greater than that in S29 group in the time range of 6~12 h (<0.05). The amylase activity in the stomach and intestine in the experimental group was significantly lower than that in S29 group (<0.05). However, the amylase activity in the liver was higher than that for S29 in the time period 6~12 h, and then, it significantly decreased after 24 h (<0.05). The SOD activity significantly decreased in S5 as time elapsed (<0.05). The SOD activity in S15 and S35 began to decrease and approached values of the activity of S29 at 120 h. The concentration of T4 in the serum of each experimental group was significantly higher than that in S29 group for the period 6~96 h (<0.05). Then, the concentration of T4 decreased with time, reaching a stable lower level at 120 h that was close to the concentration of S29. These results demonstrate that the salinity stress significantly influences the digestive enzymes, SOD, and T4 of yellowtail kingfish. Thus, it can be concluded that the yellowtail kingfish has a strong capacity for salinity adaptation. These findings also provide a basis for the aquaculture of yellowtail kingfish.
; salinity mutation; digestive enzymes; superoxide dismutase; thyroid hormone
Dec. 19, 2019
S917
A
1000-3096(2020)06-0064-09
10.11759/hykx20191219001
2019-12-19;
2020-02-25
國家重點研發計劃項目(2018YFD0901204, 2019YFD0900503);青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室開放課題(2017-3A01); 中國水產科學研究院院級基本科研業務費-農業部海洋漁業可持續發展重點實驗室開放課題資助(2019HY-XKQ01); 國家自然科學基金項目(31772829); 國家海水魚產業技術體系專項(CARS-47).
[National Key Research and Development Program, No. 2018YFD0901204, No. 2019YFD0900503; The Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), No.2017-3A01; Central Public- interest Scientific Institution Basal Research, CAFS & Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, No. 2019HY-XKQ01; The National Natural Science Foundation of China, No. 31772829; the China Agriculture Research System, No. CARS-47]
史寶(1979-), 男, 副研究員, 主要從事魚類繁育理論及增養殖技術研究, E-mail: shibao@ysfri.ac.cn; 柳學周,通信作者, E-mail: liuxz@ysfri.ac.cn.
(本文編輯: 趙衛紅)
