基于耐鹽指數STTI的柳枝稷F2代群體耐鹽性分析
2020-06-19 08:53:50劉一明孔丹宇劉小慧張訓忠趙炳宇劉國道
熱帶作物學報 2020年5期
劉一明 孔丹宇 劉小慧 張訓忠 趙炳宇 劉國道
摘? 要:雜交育種是植物耐鹽新品種選育的重要途徑之一,而對雜種F2代耐鹽能力的綜合評價至關重要。本研究以柳枝稷(Panicum virgatum)F2代的53個單株為研究對象,以親本材料Alamo、Dacotah和F1代的DA90、DA120為參照,采用沙培法,在250 mmol/L NaCl的脅迫條件下進行耐鹽性分析。在鹽處理24 d后,測定電解質外滲率(EL)、相對含水量(RWC)、干重(DW)、凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs),通過對耐鹽指數STTI進行主成分分析和隸屬函數分析,結果表明:柳枝稷F2代的不同材料間耐鹽能力存在顯著差異。親本Alamo的耐鹽能力較高,親本Dacotah的耐鹽能力較差,F1代的DA90和DA120的耐鹽能力中等,F2代的耐鹽性有明顯分離,表現在6份F2代材料的耐鹽能力強于耐鹽親本Alamo,7份F2代材料的耐鹽能力弱于不耐鹽親本Dacotah。主成分分析結果得到3個主成分,累計貢獻率達87.47%。聚類分析將57份參試材料分為5類,分別代表不同耐鹽程度的類群。隸屬函數分析表明,F2代的39、45、27、48、35、53、7號耐鹽性最強,該結果為柳枝稷耐鹽相關性狀的遺傳圖譜構建、QTL定位、柳枝稷耐鹽育種奠定了基礎。
關鍵詞:柳枝稷;耐鹽;STTI;F2代
中圖分類號:S31? ? ? 文獻標識碼:A
Salt Tolerance Analysis of F2 Generation Population of Switchgrass Based on Salt Tolerance Trait Index STTI
LIU Yiming1, KONG Danyu2, LIU Xiaohui3, ZHANG Xunzhong4, ZHAO Bingyu5*, LIU Guodao1*
1. Tropical Crop Genetic Resources Institute, Chinese Academy of Tropical Agriculture Sciences / Key Laboratory of Crop Gene Resources and Germplasm Enhancement in Southern China, Ministry of Agriculture & Rural Affairs, Danzhou, Hainan 571737, China; 2. College of Tropical Crops, Hainan University, Danzhou, Hainan 571737, China; 3. College of Agro-grassland Science, Nanjing Agricultural University, Nanjing, Jiangsu 210095, China; 4. Department of Crop and Soil Environmental Science, Virginia Tech, Blacksburg, Virginia 24061, USA; 5. Department of Horticulture, Virginia Tech, Blacksburg Virginia 24061, USA
Abstract: Cross-breeding is one of important ways to breed new varieties of plants, and it is essential to analyze the performance of hybrid F2 generation. In this study, 53 F2 generation plants of Panicum virgatum were used as the research material. The parents Alamo, Dacotah and F1 generation DA90 and DA120 were used as the reference. Salt tolerance performance was evaluated with sand culture method under 250 mmol/L NaCl stress conditions. The electrolyte leakage rate (EL), relative water content (RWC), dry weight (DW), net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) were measured after 24 days of salt treatment. Principal component analysis and membership function analysis of salt tolerance trait index (STTI) showed that there were significant differences in salt tolerance between different F2 plants. Parent Alamo was relatively salt tolerant, Dacotah was sensitive to salt, DA90 and DA120 had moderate salt tolerance, and the F2 generation had significant segregation in salt tolerance. Six F2 plants were better in salt tolerance than Alamo. Seven F2 plants were worse in salt tolerance than Dacotah. Three principal components with a cumulative contribution rate of 87.47% were selected by the principal component analysis. The 57 test plants were divided into five categories by cluster analysis. Membership function indicated that F2 generations 39, 45, 27, 48, 35, 53 and 7 had higher salt tolerance, which would build up a good basis for the construction of the genetic map of the salt tolerance-related traits, the QTL mapping and the salt-tolerant breeding of switchgrass.
Keywords: switchgrass (Panicum virgatum); salt tolerance; STTI; F2 generation
DOI: 10.3969/j.issn.1000-2561.2020.05.009
隨著能源危機和環境污染的加劇,生物質燃料越發受到關注,發展生物質能源已成為應對氣候變化、保障能源安全的重要手段[1]。柳枝稷(Panicum virgatum)為禾本科(Gramineae)黍屬(Panicum)的多年生暖季型叢生禾草(C4),它原產于北美,在墨西哥、加拿大等地均有分布。1988年柳枝稷被引入我國,并逐步被廣泛應用[2]。柳枝稷具有多年生、產量高、品質好、耐瘠薄、耐洪澇、耐干旱、抗病蟲害、種植成本低等諸多優點,在觀賞草中被廣泛應用,同時也是一種優質牧草,目前作為模式能源植物進行研究[3]。柳枝稷主要有2種生態型:高地生態型和低地生態型。低地生態型多為4倍體(2n=4x=36),主要分布于南方潮濕地帶,植株高大粗壯;高地生態型多為8倍體(2n=8x=72),主要分布在美國中部和北部地區,莖稈矮小[4]。柳枝稷可以作為鹽堿化地區的植被種植,能夠有效改善當地土壤環境,防止水土流失[5]。為避免“與糧爭地”的現象發生,常用邊際土地(鹽堿、干旱等)種植柳枝稷。全球約有10億hm2鹽堿地,我國鹽漬土地面積約1億hm2[6],因此,可利用的邊際土地資源十分豐富。
耐鹽性屬于多基因控制的數量性狀[7],探討植物對鹽分脅迫的適應機制、耐鹽性的遺傳改良一直是國內外育種工作者關注的熱點,而利用雜交育種提高植物的耐鹽性是植物新品種培育的有效手段。由于基因分離,F2代群體中會出現多種基因型個體,雜合體表現出較強的雜種優勢[8]。柳枝稷種質資源較多,具有豐富的遺傳多樣性[9],利用柳枝稷進行雜交育種,從而改良柳枝稷的耐鹽性,將會成為柳枝稷新品種培育的重要途徑。目前,柳枝稷耐鹽性的相關研究主要集中于耐鹽綜合評價、生理和分子機制方面。例如,Liu等[10]對30多份柳枝稷材料進行耐鹽評價發現,低地生態型Alamo的抗鹽性優于高地生態型Dacotah。此外,柳枝稷耐鹽性表現出多種生理機制[11];在柳枝稷耐鹽過程中,抗氧化酶(如CAT、POD、SOD)和microRNA發揮著重要作用[12-14];通過轉基因手段,超量表達AANAT和HIOMT基因,能顯著提高柳枝稷的耐鹽性,轉基因植株中的褪黑素含量顯著升高[3];鹽堿脅迫能夠抑制柳枝稷種子的萌發[15]。目前,利用雜交手段提高柳枝稷耐鹽性以及分析F2代柳枝稷群體的耐鹽能力研究尚未見報道。
本研究以耐鹽能力不同的柳枝稷親本Alamo、Dacotah和二者雜交得到的F1代DA90、DA120以及53個F2代單株為材料進行耐鹽能力分析,測定它們在鹽脅迫下的生理指標變化情況,利用相關分析和主成分分析,了解柳枝稷F2代耐鹽能力的表現,為柳枝稷的耐鹽研究提供理論參考。
1? 材料與方法
1.1? 材料
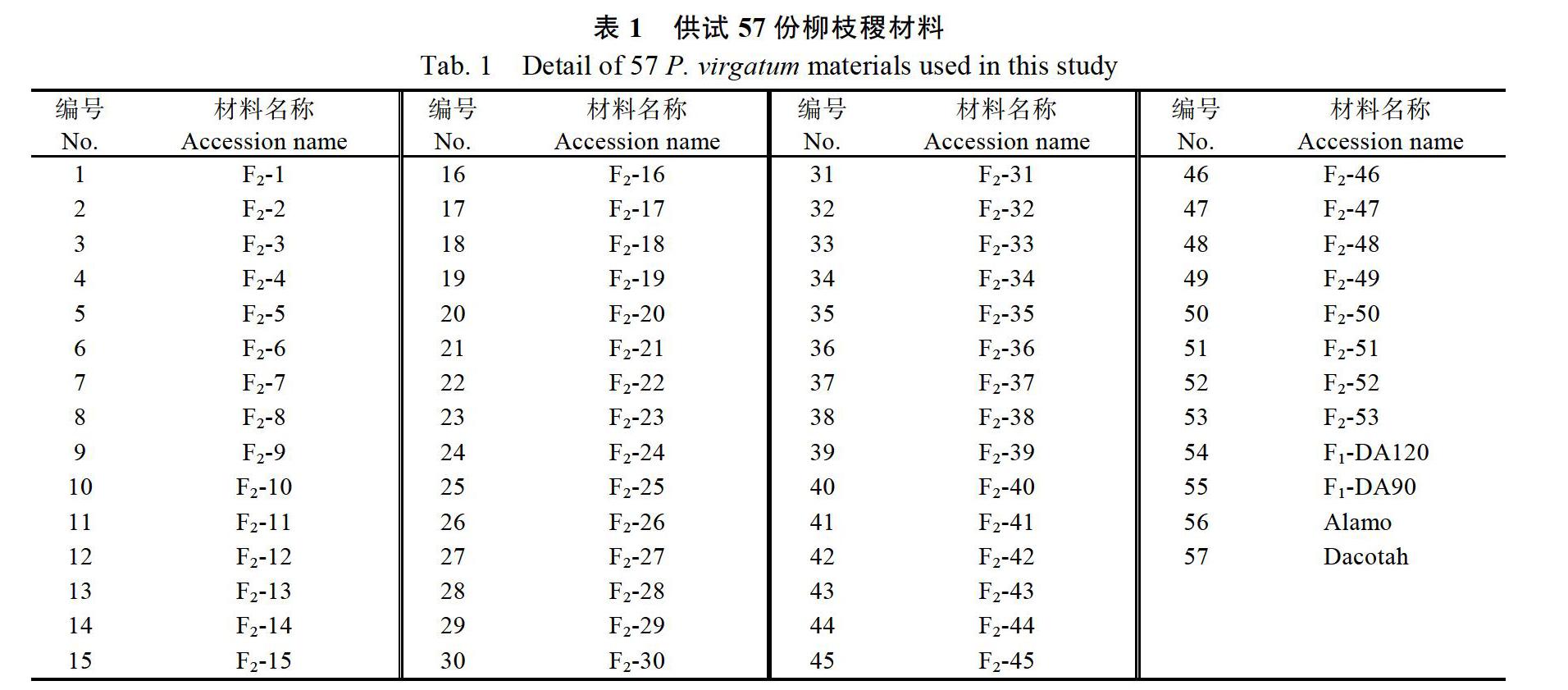
以低地型Alamo為父本,以高地型Dacotah為母本,進行雜交,獲得165個F1代單株,從F1代中選取不同單株進行雜交:單株5×49(母本),單株23×139(母本),單株90×120(母本),3組F1代雜交獲得的F2代群體性狀差異明顯。本研究所用的53份F2代材料由90和120單株雜交所得,編號分別為1~53,親本Alamo、Dacotah和F1代 DA90、DA120作為耐鹽能力的參照(表1)。
1.2? 方法
1.2.1? 實驗設計? 采用沙培法,取柳枝稷材料進行分栽,種植于塑料盆(直徑18 cm,高20 cm)中,每盆3株,每盆裝4 kg沙(粒徑0.1~1.0 mm)。定植后,在晝夜溫度為(30±2)℃/(25±2)℃,光周期為12 h,光照強度為500 μmol/(m2·s),相對濕度為75%的溫室內培養。當柳枝稷在溫室內生長2個月后進行鹽脅迫處理,鹽處理選擇250 mmol/L NaCl。鹽處理濃度每12 h遞增一次,依次從50 mmol/L增加至250 mmol/L,防止鹽沖擊效應。當盆內NaCl濃度達到250 mmol/L后,將塑料盆放在盛有Hoagland營養液的托盤中,營養液中的NaCl濃度為250 mmol/L,每天用蒸餾水補充蒸發的水分,使托盤內的鹽濃度保持恒定。實驗采用隨機區組設計,4次重復。鹽處理后的第24天進行指標測定。
1.2.2? 指標測定及方法? (1)電解質外滲率。取50 mL離心管,加入20 mL去離子水,用電導率儀(SR60IC, VWR, Radnor, PA, USA)測定初始電導率E0;剪取充分伸展的柳枝稷上部成熟葉片,用蒸餾水洗凈,吸干葉片上的水分,準確稱取0.5 g葉片,剪成1 cm的小段,置于試管中振蕩24 h,然后測定電導率E1。然后,將試管蓋緊沸水浴30 min后取出,冷卻至室溫,搖勻后測定電導率E2。最后,計算電解質外滲率EL(Leaf Ele c trolyte leakage, %)[16],EL=(E1?E0)/(E2?E0)×100%。
(2)葉片相對含水量。剪取葉片約0.5 g,用天平稱其鮮重(W1);然后將其浸入裝有去離子水的試管中,封口浸泡24 h,稱其飽和重(W2);然后,在105 ℃的烘箱中殺青30 min,85 ℃烘干至恒重,最后稱其干重(W3)。計算公式為:葉片相對含水量=[(W1?W3)/(W2?W3)]×100%[17]。
(3)光合速率。采用美國Li-6400 XT光合儀測定葉片凈光合速率[Pn, ?mol/(m2·s)]、氣孔導度[Gs, mmol/(m2·s)]和蒸騰速率(Tr, mmol/mol)進行測定。光合測定參數設定:CO2濃度為400 ?mol/mol,流速為500 ?mol/s,光照強度為1000 ?mol/(m2·s)。測量時,葉片選上部的成熟葉片,每盆測3個葉片,取平均值。
(4)干重。在實驗結束后,取所有植物植株,洗凈植株根系表面的沙子,用吸水紙吸干表面殘留的水分,置于烘箱中105 ℃殺青30 min,然后 85 ℃烘干至恒重,稱其干重(DW)。
1.2.3? 柳枝稷不同生態型的耐鹽能力綜合評價? 采用耐鹽指數(salt tolerance trait index, STTI)評價柳枝稷的耐鹽性,STTI的計算公式為:STTI=各指標的處理值/各指標的對照值×100%[18]。利用模糊數學中的隸屬函數對所有柳枝稷進行綜合評價。
1.3? 數據處理
采用SAS 8.1軟件進行方差分析和最小顯著差異性檢驗(LSD法),采用DPS 7.55軟件進行相關分析、主成分分析和聚類分析,采用Excel 2016對F2代群體的耐鹽相關指標進行頻率分布分析。
2? 結果與分析
2.1? 鹽脅迫對F2代53個柳枝稷電解質外滲率的影響
在鹽處理(250 mmol/L NaCl)24 d后,所有實驗材料的電解質外滲率明顯高于對照,且各材料的電解質外滲率STTI存在顯著差異(圖1)。參照親本Alamo和Dacotah的電解質外滲率STTI分別為133.60%和284.83%,F1代的DA90和DA120的電解質外滲率STTI分別為172.11%和226.56%,親本Alamo的耐鹽性顯著高于Dacotah,
F1代DA90的耐鹽性高于DA120,但差異不顯著。F2代的14、2、19、7、45、27號的電解質外滲率STTI均低于126%,表現出較好的耐鹽性,甚至高于耐鹽親本Alamo;F2代的26、49、4、36、33、30號的電解質外滲率STTI均高于317%,耐鹽性相對敏感,比不耐鹽親本Dacotah表現還差。
2.2? 鹽脅迫對F2代53個柳枝稷相對含水量的影響
與對照相比,在鹽處理24 d后,所有材料的相對含水量均呈下降趨勢,且不同柳枝稷材料間相對含水量差異顯著(圖2)。柳枝稷親本、F1代及53個F2代的相對含水量的趨勢與電解質外滲率基本一致,耐鹽能力依次為:Alamo>DA90>DA120> Dacotah,53個F2代材料的耐鹽能力間隔分布其中,其中F2代的45、1、48、37、35號的耐鹽性高于Alamo,相對含水量STTI均大于72.6%,而F2代的25、30、22、9、17號的耐鹽能力比親本Dacotah還要差,相對含水量STTI均小于56.2%。
2.3? 鹽脅迫對F2代53個柳枝稷凈光合速率、氣孔導度及蒸騰速率的影響
在250 mmol/L NaCl處理24 d后,所有柳枝稷材料的凈光合速率、氣孔導度、蒸騰速率都有所下降(圖3,圖4,圖5)。57份材料間的凈光合速率STTI、氣孔導度STTI、蒸騰速率STTI均差異顯著。根據3個光合相關指標的STTI可以看出,柳枝稷親本Alamo的耐鹽能力均優于F1代,F1代的耐鹽能力均優于親本Dacotah,而F1代DA90和DA120的耐鹽能力在3個指標中的表現則有所區別,凈光合速率和蒸騰速率的STTI顯示DA90的耐鹽能力優于DA120,而氣孔導度STTI表明DA120的耐鹽能力優于DA90。根據3個指標的STTI,F2代27、39、53、45號表現出較好的耐鹽性,而F2代33、9、22、25、20號則表現出較敏感的耐鹽能力。
2.4? 鹽脅迫對F2代53個柳枝稷干重的影響
在鹽處理24 d后,干重分析結果表明,耐鹽的柳枝稷材料的干重維持在較高水平,不耐鹽的柳枝稷材料干重則明顯降低(圖6)。F2代48、Alamo、F2代45、53、32、11、7號表現出較高的耐鹽能力,干重STTI均高于75.0%;F2代20、42、17、31、9、19號、Dacotah則表現出較差的耐鹽能力,干重STTI均低于44.0%。
2.5? 柳枝稷F2代鹽脅迫下各指標STTI的相關性分析和主成分分析
相關性分析表明,電解質外滲率與相對含水量、凈光合速率、蒸騰速率、氣孔導度呈極顯著負相關,相關系數依次為0.34、0.39、0.35、0.32(表2)。相對含水量、干重、光合速率、蒸騰速率、氣孔導度之間都呈極顯著正相關,其中凈光合速率與氣孔導度的相關系數最高,為0.88。
各指標STTI的主成分分析表明,主成分1、主成分2和主成分3的貢獻率分別為64.95%、15.17%和7.35%,累計貢獻率達87.47%(表3),主成分的公式為
PC1=?0.2298EL+0.4409RWC+0.3919DW+0.4614 Pn+0.4700Tr+0.4067Gs
PC2=0.9074EL+0.0597RWC+0.4127DW?0.0198Pn+0.0429Tr+0.0231Gs
PC3=0.0859EL?0.3383RWC?0.2005DW?0.2396Pn?0.0029Tr+0.8835Gs
以上3個主成分可以作為F2代柳枝稷耐鹽能力評價的主因子。
2.6? F2代柳枝稷群體耐鹽相關指標的頻率分布分析
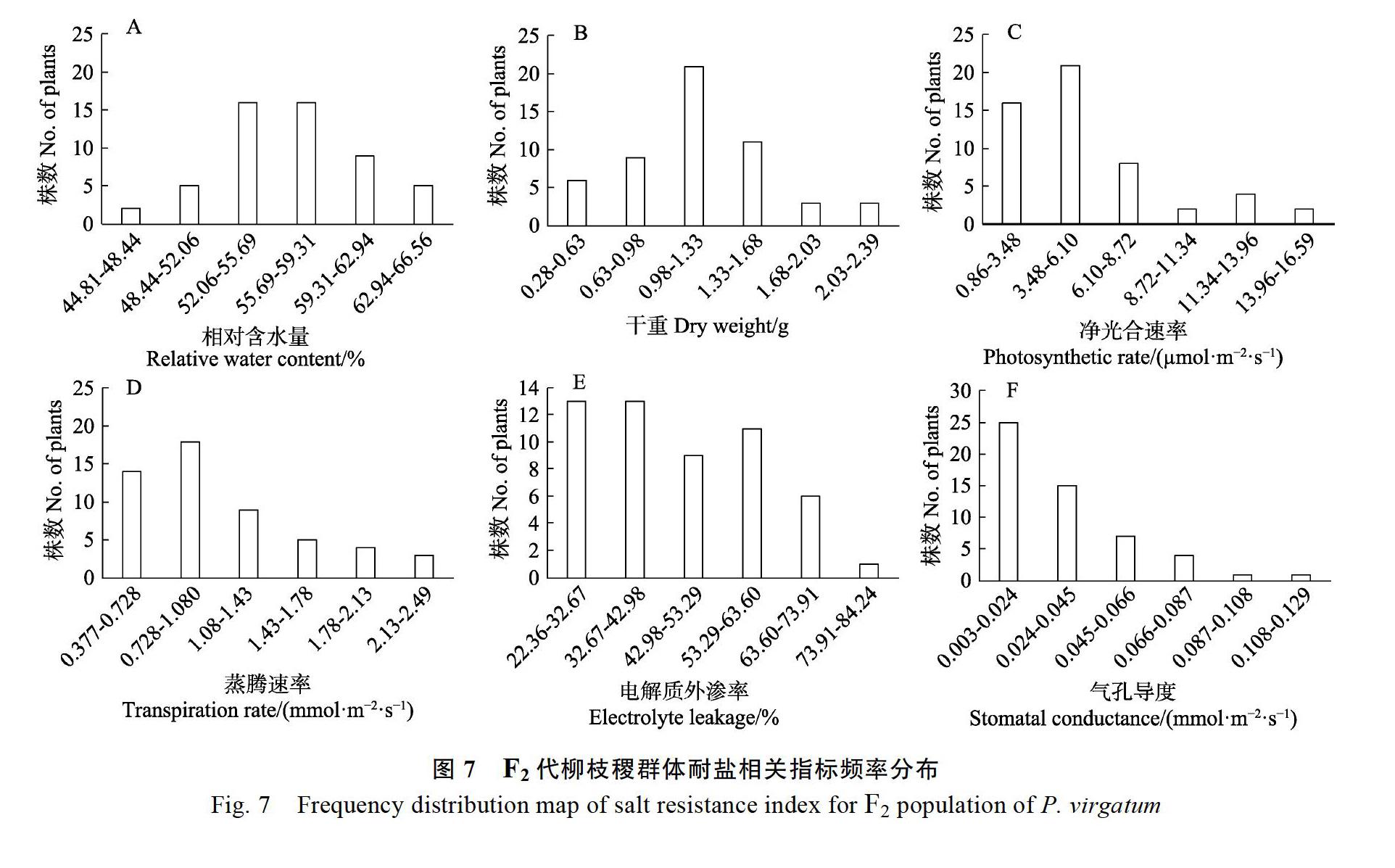
在鹽處理24 d后,分別對F2代53個柳枝稷材料的電解質外滲率、相對含水量、干重、凈光合速率、蒸騰速率、氣孔導度6個指標進行正態分布檢驗,分別以其分布范圍為橫坐標,以頻數為縱坐標,采用Excel 2016軟件對F2代群體的耐鹽相關指標作圖。結果表明,鹽脅迫下F2代群體均出現較大變異,呈不同程度的連續分布,除電解質外滲率和氣孔導度外,其他指標接近正態分布(圖7)。
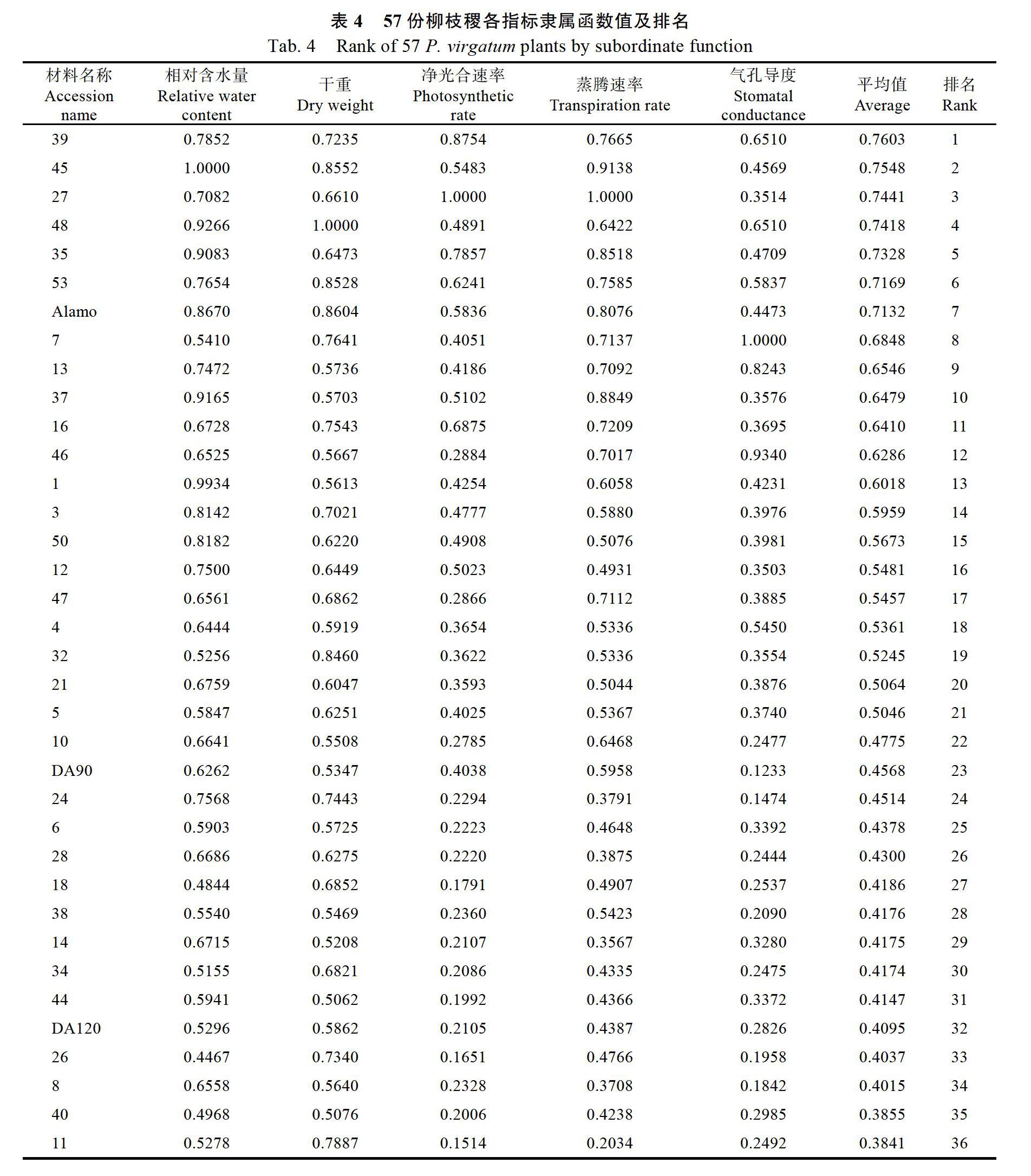
2.7? F2代柳枝稷群體的耐鹽能力綜合評價
Song L, Huang S C, Wise A, et al. A transcription factor hierarchy defines an environmental stress response network[J]. Science, 2016, 354(6312): aag1550.
陳全求, 詹先進, 藍家樣, 等. 棉花雜種F2代利用及發展研究進展[J]. 現代農業科技, 2014(23): 65-67, 70.
Liu Y M, Zhang X Z, Tran H, et al. Assessment of drought tolerance of 49 switchgrass (Panicum virgatum) genotypes using physiological and morphological parameters[J]. Biotechnology for Biofuels, 2015, 8(1): 152.
Kim J, Liu Y, Zhang X, et al. Analysis of salt-induced physiological and proline changes in 46 switchgrass (Panicum virgatum) lines indicates multiple response modes[J]. Plant Physiology and Biochemistry, 2016, 105: 203-212.
Liu Y, Zhang X, Miao J, et al. Evaluation of salinity tolerance and genetic diversity of thirty-three switchgrass (Panicum virgatum) populations[J]. BioEnergy Research, 2014, 7(4): 1329-1342.
Sun G, Stewart C N, Xiao P, et al. MicroRNA expression analysis in the cellulosic biofuel crop switchgrass (Panicum virgatum) under abiotic stress[J]. PLoS One, 2012, 7(3): e32017.
Wang Q, Wu C, Xie B, et al. Model analysing the antioxidant responses of leaves and roots of switchgrass to NaCl-salinity stress[J]. Plant Physiology and Biochemistry, 2012, 58: 288-296.
Xie F, Stewart C N, Taki F A, et al. High-throughput deep sequencing shows that microRNAs play important roles in switchgrass responses to drought and salinity stress[J]. Plant Biotechnology Journal, 2014, 12(3): 354-366.
Liu Y, Wang Q, Zhang Y, et al. Synergistic and antagonistic effects of salinity and pH on germination in switchgrass (Panicum virgatum L.)[J]. PloS One, 2014, 9(1): e85282.
Houimli S I M, Denden M, Mouhandes B D. Effects of 24-epibrassinolide on growth, chlorophyll, electrolyte leakage and proline by pepper plants under NaCl-stress[J]. EurAsian Journal of BioSciences, 2010, 4(1): 96-104.
Liu Y, Du H, Wang K, et al. Differential photosynthetic responses to salinity stress between two perennial grass species contrasting in salinity tolerance[J]. HortScience, 2011, 46(2): 311-16.
Tavakkoli E, Fatehi F, Rengasamy P, et al. A comparison of hydroponic and soil-based screening methods to identify salt tolerance in the field in barley[J]. Journal of Experimental Botany, 2012, 63(10): 3853-3867.
董榮書. 接種根瘤菌對柱花草耐鹽性的影響及機理研究[D]. 海口: 海南大學, 2014.
崔彥茹. 利用選擇育種群體進行水稻高產、抗旱和耐鹽QTL定位[D]. 北京: 中國農業科學院, 2015.
邵? 安. 利用小麥F2代(SR3 X JN17)群體進行鹽脅迫相關主效QTL的SSR及EST-SSR定位[D]. 濟南: 山東大學, 2010.
王喜濤. 黃瓜種質資源耐鹽性評價及其遺傳分析[D]. 哈爾濱: 東北農業大學, 2014.
張艷芳. 番茄特異PCR遺傳連鎖圖譜的構建及苗期耐鹽QTL定位[D]. 呼和浩特: 內蒙古農業大學, 2008.
龍衛華. 油菜發芽期耐鹽評價、篩選與鹽脅迫下根轉錄組分析[D]. 北京: 中國農業科學院, 2015.
孫? 飛. 高粱雜交種耐鹽性評價及耐鹽生理機制研究[D]. 沈陽: 沈陽農業大學, 2018.
孫涌棟, 郝峰鴿, 徐? 斌. 種子萌發期南瓜雜交品種耐鹽評價[J]. 河南農業科學, 2009(10): 111-114.
Negr?o S, Schm?ckel S M, Tester M. Evaluating physiological responses of plants to salinity stress[J]. Annals of Botany, 2017, 119(1): 1-11.
Munns R, Tester M. Mechanisms of salinity tolerance[J]. Annual Review of Plant Biology. 2008, 59: 651-681.
Parida A K, Das A B. Salt tolerance and salinity effects on plants: a review[J]. Ecotoxicology and Environmental Safety, 2005, 60(3): 324-349.
Munir S, Siddiqi E H, Bhatti K H, et al. Assessment of inter-cultivar variations for salinity tolerance in winter radish (Raphanus sativus L.) using photosynthetic attributes as effective selection criteria[J]. World Applied Sciences Journal, 2013, 21(3): 384-388.
