尖葉牛樟染色體制片優(yōu)化與核型分析
2020-06-19 08:53:50官錦燕羅青文譚嘉娜黃海英文明富羅劍飄
熱帶作物學(xué)報 2020年5期
官錦燕 羅青文 譚嘉娜 黃海英 文明富 羅劍飄

摘? 要:本研究以尖葉牛樟根尖為材料,采用酶解去壁低滲法制片,探討不同預(yù)處理方式和酶解時間對牛樟根尖染色體制片的影響,并對其進行核型分析,以期為牛樟的起源、演化及遺傳育種提供一定的理論依據(jù)。結(jié)果表明:于9:00—11:00進行取材,用0.002 mol/L 8-羥基喹啉液預(yù)處理2 h,4%果膠酶和5%纖維素酶酶解5 h,能獲得較佳的染色體制片。核型分析結(jié)果表明:尖葉牛樟的染色體數(shù)目為24條,核型公式為2n=2x=24=22m+2sm,第10對染色體帶有隨體,屬于2B類型,核型不對稱系數(shù)55.81%。核型對稱程度較高,這表明尖葉牛樟可能進化程度較低,屬于比較原始的物種。
關(guān)鍵詞:尖葉牛樟;根尖;染色體制片;核型分析
中圖分類號:Q943.2? ? ? 文獻標識碼:A
Optimization of Chromosome Mounting Technique and Karyotype Analysisi of Cinnamomum kanehirae Hay of Sharp Leaves
GUAN Jinyan, LUO Qingwen, TAN Jiana, HUANG Haiying, WEN Mingfu, LUO Jianpiao*
Guangdong key Lab of Sugarcane Improvement? Biorefinery / Guandong Provincial Bioengingeering Institute Guangzhou Sugarcane Industry Research Institute, Guangzhou, Guangdong 510316, China
Abstract: In order tooptimize the chromosome mounting technique and explore the chromosome karyotype of C. kanehirae Hay, and to provide important cytological evidences for the study of the evolution, evolutionary characteristics and genetic regularity of camphor plants, the effects of differentpretreatment methods and enzymatic hydrolysis time on the production of C. kanehirae were discussed by the enzymatic dissociation wall low permeability method. The observation showed that pretreated in 0.002 mol/L 8-hydroxyl for 2 h, dissociated in 4% cellulase and 5% pectinase for 5 h could achieve the optimal experimental results. The karyotype analysis showed that the chromosome number was 24, with a karyotype formula 2n=2x=24=22m+2sm. The results demonstrated that the asymmetry index was 55.81%, the karyotype was 2B type, suggesting that C. kanehirae Hay maybe a relatively primitive species.
Keywords: Cinnamomum kanehirae Hay of sharp leaves; root tip; chromosome preparation; karyotype analysis
DOI: 10.3969/j.issn.1000-2561.2020.05.007
牛樟(Cinnamomum kanehirae Hay)隸屬于樟科樟屬,又名黑樟,為中國臺灣地區(qū)特有常綠闊葉樹種[1]。牛樟具有很高的經(jīng)濟價值,根、莖、葉全株可提煉牛樟精油,木材是雕刻家具的上等材料。此外,牛樟是牛樟芝(Antrodia camphorata)的唯一宿主,牛樟芝在醫(yī)學(xué)上被稱為“藥芝之王”,常用來醒酒與緩解疲勞,具有免疫調(diào)節(jié)、保護肝臟等作用[2]。有關(guān)牛樟的研究起步晚,研究方向單一且以宏觀研究為主,主要集中在牛樟的組培快繁[3-4]、扦插[5]、引種[6]等方面。近年來,在牛樟分子生物學(xué)遺傳多態(tài)性上的研究取得了一些進展[7-8],但有關(guān)牛樟細胞遺傳學(xué)上的研究至今
仍舊空白。
染色體核型分析主要通過研究其染色體數(shù)目及形態(tài)特征,反映不同物種或品種之間存在的染色體細胞學(xué)差異[9]。不同物種的染色體核型模式在進化的過程中不受外界環(huán)境因素干擾和影響,在一定程度上反映出其進化的基本特征[10]。因此,植物核型的研究不僅能為種間遺傳變異、系統(tǒng)演化以及親緣關(guān)系提供證據(jù),還可以為雜交育種選育及雜種后代鑒定等提供理論依據(jù)[11]。樟科植物核型的研究已取得一些進展,已報道過核型研究的樟科植物共有42種,涉及樟屬、楠屬、潤楠屬、新木姜子屬、無根藤屬和檫木屬等多屬品種。Okada等[12]于1975年報道了樟科9屬16種植物的染色體數(shù)目,基數(shù)均為12。陳成彬等[13]于1998年報道了樟科5屬9種植物的染色體數(shù)目,均為2n=24。陳細芳等[14]于2009年報道了浙江楠的染色體數(shù)目為2n=24。劉玉香[15]于2013年報道了樟科5屬17種植物的染色體數(shù)目為24。在這42種植物中,核型類型為“1A”“1B”“2A”“2B”4種類型,尚未發(fā)現(xiàn)C類型的核型。其中,樟屬中有10種植物的核型已報道,包括大葉樟、沉水樟、陰香、樟樹、油樟等,染色體均為24條,核型類型為2B、2A兩種類型。至今,有關(guān)牛樟的染色體核型分析的研究仍未見報道。因此,本研究以尖葉牛樟根尖為材料,對其染色體制片技術(shù)進行優(yōu)化并對其進行核型分析,為牛樟的演化、進化特征和遺傳規(guī)律的研究提供重要的細胞遺傳學(xué)佐證。
1? 材料與方法
1.1? 材料
試驗材料為尖葉牛樟組培苗的根尖,采自廣東省生物工程研究所湛江甘蔗研究中心良種組培實驗室。
1.2? 方法
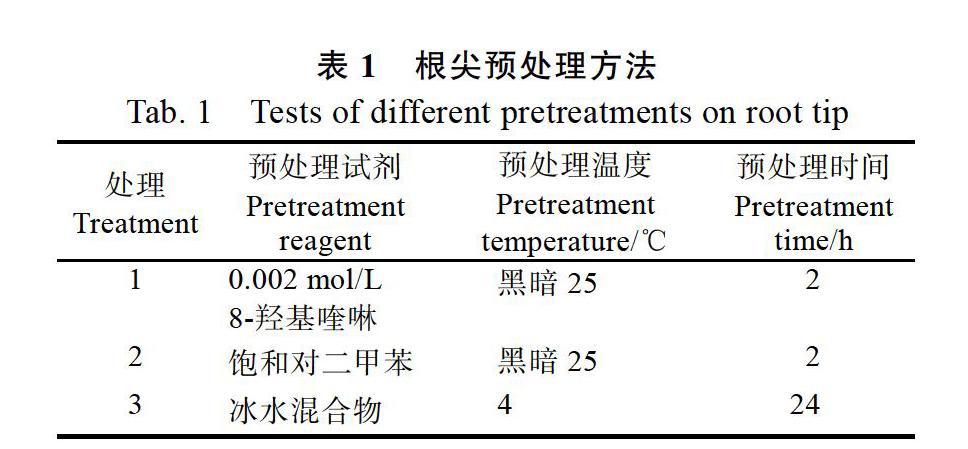
1.2.1? 取材和預(yù)處理? 于9:00—11:00采集牛樟組培苗健壯、白嫩的根尖(0.5~1 cm),分別采用以下方法(表1)進行預(yù)處理。再清水沖洗預(yù)處理液3~4次,把根尖移至卡諾固定液(無水乙醇∶冰乙酸的比例為3∶1)固定24 h,最后置于70%乙醇中4 ℃保存?zhèn)溆谩?/p>
1.2.2? 染色體標本制備? 染色體標本制備采用酶解去壁低滲法,具體步驟參照官錦燕[16]碩士論文。
(1)前低滲:把固定好根尖置于蒸餾水中進行前低滲30 min。
(2)酶解:把步驟(1)的根尖置于盛有5%纖維素酶和4%果膠酶混合液的1 mL離心管中,并置于37 ℃的恒溫水浴鍋中酶解3~5 h。
(3)后低滲:吸取酶液,緩緩地沿著管壁加入蒸餾水,后低滲30 min。
(4)后固定:吸取蒸餾水,滴加臨時配制的固定液,后固定15 min。
(5)涂片:把玻片、臨時配制的固定液均置于冰面上,取1張玻片,先滴3滴固定液,再夾取根尖置于玻片上,用解剖針挑取白色的根尖分生組織,并迅速搗碎再均勻涂抹在載玻片上,再從載玻片一端滴加1滴固定液使細胞擴散,接著在火焰上迅速烘烤,自然風(fēng)干。
(6)染色:將KH2PO4和Na2HPO4混合緩沖液(pH 6.8)與吉姆薩原液按比例為49∶1配制成工作液,再把干燥的制片置于吉姆薩染色液中染色10~20 min,清水沖洗干凈,置烘箱37 ℃烘干。
1.2.3? 核型分析? 用POTIKA正立顯微鏡觀察,Optika Vision pro拍照系統(tǒng)拍照。選擇30個染色體分散且形態(tài)較好的細胞進行染色體數(shù)目統(tǒng)計,選取5個染色體形態(tài)清晰且無重疊的細胞,用Photoshop圖像軟件進行核型分析。核型分析方法參照李懋學(xué)等[17]的標準,核型對稱性按照Stebbins[18]的標準劃分,染色體形態(tài)根據(jù)Levan等[19]的方法歸類。
臂比(r)=長臂(S)/短臂(L)
染色體相對長度=染色體長度/染色體組總度×100%
平均長度核型不對稱系數(shù)(As.k)=長臂總長/全組染色體總長×100%
2? 結(jié)果與分析
2.1? 牛樟染色體制片技術(shù)的優(yōu)化
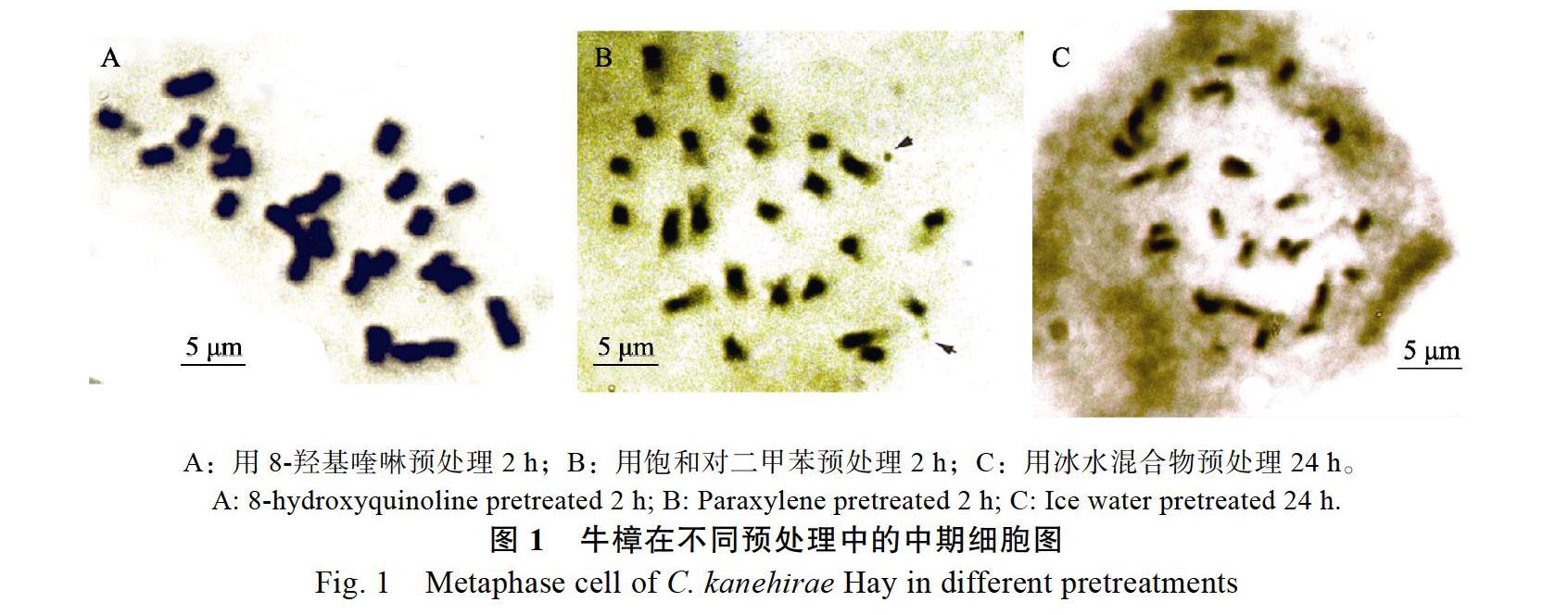
2.1.1? 不同預(yù)處理方法對染色體制片效果的影響? 3種不同預(yù)處理方法對牛樟根尖處理的結(jié)果表明(表2和圖1):不同預(yù)處理方法對中期細胞數(shù)目及染色體分散效果及清晰度都有一定的影響。8-羥基喹啉預(yù)處理2 h所得的中期細胞所占比率為14.37%,明顯高于其他處理組,染色體較分散、聚縮程度適中、形態(tài)和溢痕清晰,很適合用于核型分析研究。其次是飽和對二甲苯預(yù)處理2 h,染色體分散、聚縮,但有拖尾現(xiàn)象、縊痕不夠清晰,不適合進行核型分析。而冰水混合物處理效果最差,染色體不夠分散和聚縮、形態(tài)不清晰,不利于染色體計數(shù)和核型分析。
2.1.2? 不同的酶解時間對染色體制片效果的影響? 不同酶解時間對牛樟染色體制片效果影響的結(jié)果表明,酶解時間對牛樟染色體制片效果的影響主要體現(xiàn)在制片背景和染色體的清晰度上(圖2)。酶解5 h的細胞效果最佳,沒有細胞壁、細胞質(zhì)很稀薄、背景干凈、染色體分散,有利于進行核型分析。酶解4 h細胞效果一般,沒有細胞壁、但細胞質(zhì)比較厚、背景不夠干凈。酶解3 h制片效果最差(圖2),細胞壁沒有溶解完、細胞質(zhì)濃厚、背景不干凈,不利于染色體識別和進行核型分析。
2.2? 牛樟染色體核型分析
2.2.1? 染色體數(shù)目? 尖葉牛樟的染色體數(shù)目為2n=24,為二倍體。
2.2.2? 核型特征? 尖葉牛樟的染色體核型參數(shù)(表3)及染色體核型分析圖(圖3)分析結(jié)果表明,尖葉牛樟的核型公式為2n=2x=24=22m+2sm;
除第8對為近中部著絲點染色體(sm)外,其余為中部著絲粒染色體(m),發(fā)現(xiàn)第10對染色體上有隨體。其染色體相對長度范圍為5.35%~ 12.22%,最長與最短染色體長度比值為2.28,臂比值范圍為1.1~2.3,其中臂比值大于2的染色體有1對,為第8對染色體;核不對稱系數(shù)為55.81%,為2B型。
3? 討論
3.1? 尖葉牛樟染色體制片的優(yōu)化
染色體制片技術(shù)是核型分析、基因定位等細胞學(xué)研究的基礎(chǔ)。不同的物種在取材、預(yù)處理、解離和制片方法上有較大的差異,預(yù)處理和解離的效果直接關(guān)系到制片的質(zhì)量。劉玉香[15]對17種樟科植物進行核型分析研究,表明以根尖為材料,用0.002 mol/L 8-羥基喹啉溶液預(yù)處理2 h,采用壓片法在60 ℃的1 mol/L HCl解離3 min能達到理想的處理效果。陳細芳等[14]對浙江楠進行核型分析研究采用對二氯苯飽和預(yù)液處理根尖3~5 h,用壓片法在60 ℃的1 mol/L HCl解離12~ 15 min也取得較好的效果。陳成彬等[13]采用酶解去壁低滲法對樟科5屬9種植物進行核型分析。本研究首次報道牛樟染色體制片技術(shù)和核型,研究表明采用8-羥基喹啉在黑暗條件下對根尖預(yù)處理2 h,采用酶解去壁低滲法制片并以5%纖維素酶和4%果膠酶混合液酶解5 h,所得制片效果最佳,預(yù)處理方式和劉玉香[15]所采用的相同。采用壓片法和酶解去壁低滲法均能獲得良好效果。
3.2? 牛樟染色體數(shù)目及核型特征的探討
同一物種或品種的染色體數(shù)目在進化過程中是比較恒定的,染色體核型穩(wěn)定特征被作為分類指標,對闡述植物進化程度和分類方面有重要意義[20]。前人已開展核型研究的樟科植物共有42種,染色體基數(shù)均為12,但染色體倍性不同[12-15]。有38種植物的染色體數(shù)目為2n=24條,為二倍體。另4種植物為多倍體,其中無根藤、月桂和美洲檫木3種植物的染色體數(shù)目為2n=4x=48條,為四倍體。臺灣新木姜子的染色體數(shù)目為2n=6x=72,為六倍體。本研究首次報道了尖葉牛樟的染色體數(shù)目為24條,核型為2n=2x=24=22m+2sm,符合樟科植物的染色體基數(shù)為12的規(guī)律。
Stebbins[18]對大量核型資料進行分析,認為核型由對稱向不對稱發(fā)展是植物界核型進化的基本趨勢;核型不對稱系數(shù)越接近50%,核型的對稱程度越高,進化程度越低,屬于越原始的植物。對已報道的42種樟科植物,發(fā)現(xiàn)核型有“1A”“1B”“2A”“2B”4種類型,沒有發(fā)現(xiàn)“C”類型的核型,說明樟科植物的核型總體上較為對稱,在系統(tǒng)進化中可能屬于較為原始的類型。其中,樟屬中大葉樟、沉水樟等4種屬于2B類型,核不對稱系數(shù)在56.44%~60.78%之間。本研究得出尖葉牛樟核型不對稱系數(shù)為55.81%,接近50%,且屬于2B類型,由此推斷尖葉牛樟可能屬于比較原始的物種,進化程度較低。
尖葉牛樟常被誤認為樟科樟屬中的另一樹種沉水樟(Cinnamomum micranthum Hayata),兩者外形極似,都是高大的常綠喬木[1]。劉玉香等[15]曾報道了沉水樟核型為2n=2x=24=20m+4st,第12號染色體有隨體,核不對稱系數(shù)為58.60%,為2B類型。與本研究中尖葉牛樟的核型和核不對稱系數(shù)相似,且都有隨體(牛樟在第8號染色體上),均為2B類型。可見牛樟和沉水樟不僅外形極似,核內(nèi)的染色體核型也相似。核型是物種特有的再現(xiàn)性很高的細胞遺傳學(xué)信息,核型分析的研究對牛樟資源的應(yīng)用具有非常重要的價值。
參考文獻
官錦燕, 譚嘉娜, 羅劍飄, 等. 牛樟的組織培養(yǎng)和植株再生[J]. 南京林業(yè)大學(xué)學(xué)報(自然科學(xué)版), 2016, 40(4): 63-68.
Tien A J, Chien C Y, Chen Y H, et al. Fruiting bodies of
Antrodia cinnamomea and its active triterpenoid, antcin K, ameliorates N-nitrosodiethylamine-induced hepatic inflammation, fibrosis and carcinogenesis in rats[J]. The American Journal of Chinese Medicine, 2017, 45(1): 173-198.
戴小英, 雷昌全, 劉新亮, 等. 牛樟組培快繁體系的建立與優(yōu)化[J]. 南方林業(yè)科學(xué), 2018, 46(6): 39-44.
辛亞龍, 唐軍榮, 楊宇明, 等. 牛樟組織培養(yǎng)技術(shù)研究[J]. 中南林業(yè)科技大學(xué)學(xué)報, 2017, 37(8): 48-53.
章? 挺, 邱鳳英, 劉新亮, 等. 插穗和基質(zhì)對牛樟扦插繁殖的影響[J]. 南方林業(yè)科學(xué), 2018, 46(4): 15-18, 23.
邢文婷, 許? 奕, 陳? 培, 等. 臺灣牛樟在海南引種栽培及生長特性研究[J]. 熱帶作物學(xué)報, 2018, 39(4): 630-634.
郭? 鶯, 孟紅巖, 林文珍, 等. 牛樟EST-SSR標記的開發(fā)及遺傳多態(tài)性分析[J]. 熱帶作物學(xué)報, 2018, 39(8): 1561- 1569.
Liao P C, Kuo D C, Lin C C, et al. Historical spatial range expansion and a very recent bottleneck of Cinnamomum kanehirae Hay. (Lauraceae) in Taiwan inferred from nuclear genes[J]. BMC Evolutionary Biology, 2010, 10(1): 124.
徐洪國, 祁宏英, 顧靈杰. 黃果龍葵和龍葵染色體制片優(yōu)化及核型分析[J]. 西北植物學(xué)報, 2017, 37(2): 387-393.
謝卓宓, 牛? 歡, 古? 力, 等. 9種金線蓮資源的染色體倍性及其核型分析[J]. 中國現(xiàn)代中藥, 2018, 20(8): 920-927.
楊漢波, 饒龍兵, 郭洪英, 等. 5種榿木屬植物的核型分析[J]. 植物遺傳資源學(xué)報, 2013, 14(6): 1203-1207.
Okada H, Tartaka R. Karyological studies in some species of Lauraceae[J]. Taxon, 1975, 24(2/3): 271-280.
陳成彬, 李秀蘭, 孫成仁, 等. 中國樟科5屬9種植物的核型研究[J]. 武漢植物學(xué)研究, 1998, 16(3): 219-222, 294.
陳細芳, 郝明明, 陳? 菽, 等. 浙江楠染色體核型分析[J]. 浙江林業(yè)科技, 2009, 29(6): 26-28.
劉玉香. 樟科主要屬種核型研究及其親緣關(guān)系的ISSR分析[D]. 南昌: 江西農(nóng)業(yè)大學(xué), 2013.
官錦燕. 巴西橡膠樹SUT和RZF基因家族物理定位的研究[D]. 海口: 海南大學(xué), 2014.
李懋學(xué), 陳瑞陽. 關(guān)于植物核型分析的標準化問題[J]. 武漢植物學(xué)研究, 1985, 3(4): 297-302.
Stebbins G L. Variantion and Evolution in Plants[M]. Co1umbia: Columbia University Press, 1950.
Levan A, Fredga K, Sandberg A A. Nomenclature for centromeric position on chromosomes[J]. Hereditas, 1964, 52(2): 201-220.
任文娟, 郭小菲, 姜立娜, 等. 菜用大黃染色體制片優(yōu)化及核型分析[J]. 華北農(nóng)學(xué)報, 2013, 28(5): 128-132.
