長湖浮游植物優勢種季節演替及生態位分析
2020-06-13 03:27:36魏志兵羅靜波譚鳳霞楊德國何勇鳳
水生生物學報 2020年3期
魏志兵 柴 毅 羅靜波 郭 坤 譚鳳霞 楊德國 何勇鳳
(1. 長江大學濕地生態與農業利用教育部工程研究中心, 荊州 434025; 2. 長江大學動物科學學院, 荊州 434025; 3. 中國水產科學研究院長江水產研究所, 農業部淡水生物多樣性保護重點實驗室, 武漢 430223)
生態位(Niche)是指物種在其生存環境中時間、空間分布和生物量占有等系列利用資源的能力, 受群落內生物和非生物環境的影響, 與種間競爭密切相關[1,2]。生態位寬度是指生物種群在群落中所利用不同資源的能力總和, 生態位越寬, 其競爭能力越強。生態位重疊體現了兩個物種對群落內共同資源的利用程度, 也是研究種間競爭的一個重要因子[3]。目前生態位理論已被廣泛應用于陸生生物如昆蟲、花卉、森林等[4—7]和水生生物如魚類、浮游生物和大型底棲動物[8—10]。其中浮游植物生態位的相關研究主要集中在內陸典型湖泊[11—13],可對湖泊富營養化狀態進行綜合評價并預測其發展趨勢。近年來隨著多元統計分析的廣泛運用, 多種生態學參數如OMI指數(Outlying Mean Index, 邊緣平均指數)和TOI指數(Tolerance Index, 耐受指數)等也被用于物種生態位與環境之間的相關性分析, 被認為是較好的生態位研究應用[14]。
長湖作為我國長江中下游和江漢平原地區重要的生態功能區, 具有防洪調蓄、供水灌溉、航運和旅游等多種重要功能, 2010年被農業部批建為鲌類國家級水產種資源保護區。研究發現, 由于人類活動如工業廢水排放、農業面源污染、大面積圍網圍欄養殖等影響, 長湖處于富營養化狀態[15—19]。而近年來引江濟漢、蒙華鐵路和318國道長湖特大橋等涉水工程的陸續建設也會對該水域產生不同程度的影響。本文運用多種生態學參數對長湖浮游植物優勢種生態位特征進行分析評價, 并與本課題組2012年的研究報道[11]做對比, 以期對長湖浮游植物生態位進行深入和全面的分析, 揭示其演替規律并科學預測其發展趨勢。為長湖水生生物資源的可持續發展提供重要參考, 旨在為下一步長湖生態系統的功能恢復、價值評價以及健康良好生態系統多樣性格局的維護提供科學依據。
1 材料與方法
1.1 區域概況
長湖現有流域面積約3240 km, 平均水深2.1 m,年均水溫為17.5℃, 常年光照充足, 其年降雨量在1200 mm左右[16]。如圖1所示, 長湖自西向東分為廟湖、海子湖、馬洪臺和圓心湖4個區域。受“引江濟漢”工程建設影響, 廟湖已被兩條堤壩攔截形成靜水, 故在此區域不設置采樣點。海子湖為一狹長形湖灣, 分布有5個進出水口, 馬洪臺和圓心湖為開闊水域, 均有較大面積的圍網圍欄養殖。
1.2 采樣點設置與采樣時間
采樣點的設置參考《水域生態系統觀測規范》[20]進行并根據長湖實際水域形態、進出水口以及水文特征進行(表1), 共設置14個采樣點, 其中海子湖區4個(H1—4), 馬洪臺區(M1—5)和圓心湖區(Y1—5)各5個, 每次取樣均用GPS儀進行樣點定位。采樣時間分別為2015年4月(春季)、7月(夏季)、11月(秋季)和2016年1月(冬季)。
1.3 樣本的采集與分析
使用多參數水質分析儀(HQ40D, Hach Loveland, USA)現場測定溶解氧(DO)、水溫(WT)和pH,利用塞氏盤測定透明度(SD)。總磷(TP)、總氮(TN)、磷酸鹽硝態氮亞硝態氮和氨氮等水化學指標按照國家水質標準方法GB 3828-2002進行測定。
浮游植物定性樣本使用25#浮游生物網(網目65 μm)進行采集, 加4%甲醛固定后帶回實驗室用于種類鑒定。定量樣本使用有機玻璃采水器(1 L)在表層至0.5 m深處和0.5—1.0 m深處分別采樣, 樣本充分混合后取1 L加入15 mL魯哥氏液固定, 帶回實驗室后沉淀、濃縮至30—50 mL用以定量計數[21]。
使用顯微鏡(XSM-20, 寧波舜宇儀器有限公司,寧波, 浙江)對浮游植物樣本進行種類鑒定和計數,浮游植物鑒定準則參考《中國淡水藻類-系統、分類及生態》, 且所有種類鑒定到種[22,23]。
1.4 數據分析
優勢種采用Mcnaughton優勢度指數(Y)來確定,Y>0.02時即視為優勢種。生態位寬度(Bi)采用以Shannon-Wiener多樣性指數為基礎的生態位寬度公式計算。生態位重疊指數(Oik)采用Levins指數計算。各公式如下:
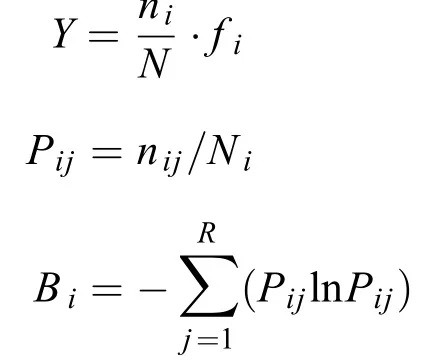
圖1 長湖采樣點示意圖Fig. 1 The sampling sites in Changhu Lake
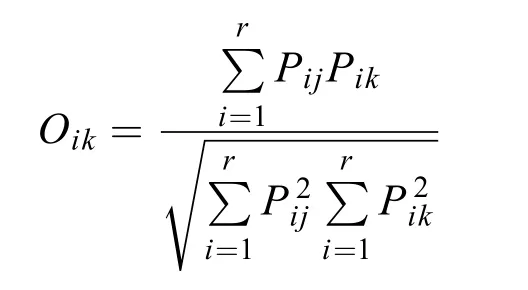
式中,ni為第i種在各采樣點的總個體數,N為各個采樣點中浮游植物的總個體數,fi為第i種在各個采樣點出現的頻率,nij為第i種在j采樣點的個體數,Pij表示種i在第j個采樣點的個體數占該物種所有個體數的比,r和R為采樣點總數,Oik為種i與種k的資源利用曲線的生態位重疊指數,Pik為種i在j采樣點的個體數占該種所有個體數的比值。
平均邊緣指數(OMI)是通過設立其適合生境是所在調查區域平均生境的假定物種,再計算該物種與假定物種之間的距離。耐受指數(TOI)是通過計算物種調查區域物種沿環境梯度分布范圍的寬窄。
豐度和優勢度的相關計算和做圖通過Excel 2013完成。其他生態位相關分析均通過R_V3.4.3的spaa和vegan程序包完成, 使用vegan包中的相關函數進行計算和制圖, 同時進行顯著性檢驗(以P表示)[24]。
2 結果
2.1 浮游植物優勢種群落結構特征
浮游植物組成及優勢種本次調查共鑒定出浮游植物7門67屬120種, 主要包括綠藻門(61種,占比50.8%)、硅藻門(25種, 占比20.8%)和藍藻門(18種, 占比15.0%), 其余裸藻門、黃藻門、金藻門和甲藻門共16種, 共占比13.4%。如表1所示, 優勢種共14種, 隸屬于5門14屬, 其中扭曲小環藻(Cyclotella comta)、小球藻(Chlorella vulgaris)和梭形裸藻(Euglena acus)是4個季節的共有優勢種。
以出現頻率超過90%的優勢種的季節分布來看, 春季是小球藻、鐮形纖維藻和四尾柵藻; 夏季是湖泊色球藻(Chlorella limneticus)、類顫魚腥藻(Osicellariordes)、微小平裂藻(Merismopedia tenuissima)、小球藻和四尾柵藻, 其中除四尾柵藻外其余4種的出現頻率均為100%; 秋季是湖泊色球藻、四尾柵藻、肘狀針桿藻(Synedra ulna)和梭形裸藻, 其中梭形裸藻為100%; 冬季是扭曲小環藻、肘狀針桿藻、帽形菱形藻(Nitzschia cap)和梭形裸藻, 其中肘狀針桿藻的為100%。

表1 長湖浮游植物優勢種出現頻率和優勢度指數Tab. 1 Occurrence frequency and dominance index of dominant species phytoplankton in Changhu Lake
以各季節分布的優勢度來看, 春季優勢度指數最高的是扭曲小環藻(0.196), 而小球藻在夏、秋、冬3個季節的優勢度指數均為最高, 分別為0.401、0.235和0.222。
優勢種豐度如圖2所示, 總體來看, 總豐度呈夏>春>秋>冬的變化趨勢, 各藻門豐度在季節和采樣點的分布上則表現出較明顯的差異性。硅藻門在春季整體占比較高, 其中采樣點H2(7.86×106cells/L)、H4(9.86×106cells/L)、M2(0.53×106cells/L)和M3(3.83×106cells/L)的占比均超過50%, 其中H4占比最高, 達79.9%。藍藻門在夏季的占比最高, 其中采樣點Y1(9.94×106cells/L)、Y2(11.50×106cells/L)、Y4(4.15×106cells/L)和Y5(9.73×106cells/L)的占比均超60%, 最高達80%(Y5)。綠藻門的整體占比均較高, 其中超過60%的有春M1(2.21×106cells/L)、M4(3.27×106cells/L)、M5(6.83×106cells/L)、Y1(1.60×106cells/L)、Y2(1.94×106cells/L)和夏H3(22.70×106cells/L)、M1(12.10×106cells/L)、M2(8.38×106cells/L)、M3(33.50×106cells/L)以及秋Y1(5.63×106cells/L)、Y2(3.60×106cells/L)和冬M4(2.46×106cells/L)共計12個采樣點。其中春M5占比最高, 達83.7%。裸藻門在四季的整體占比均較低。
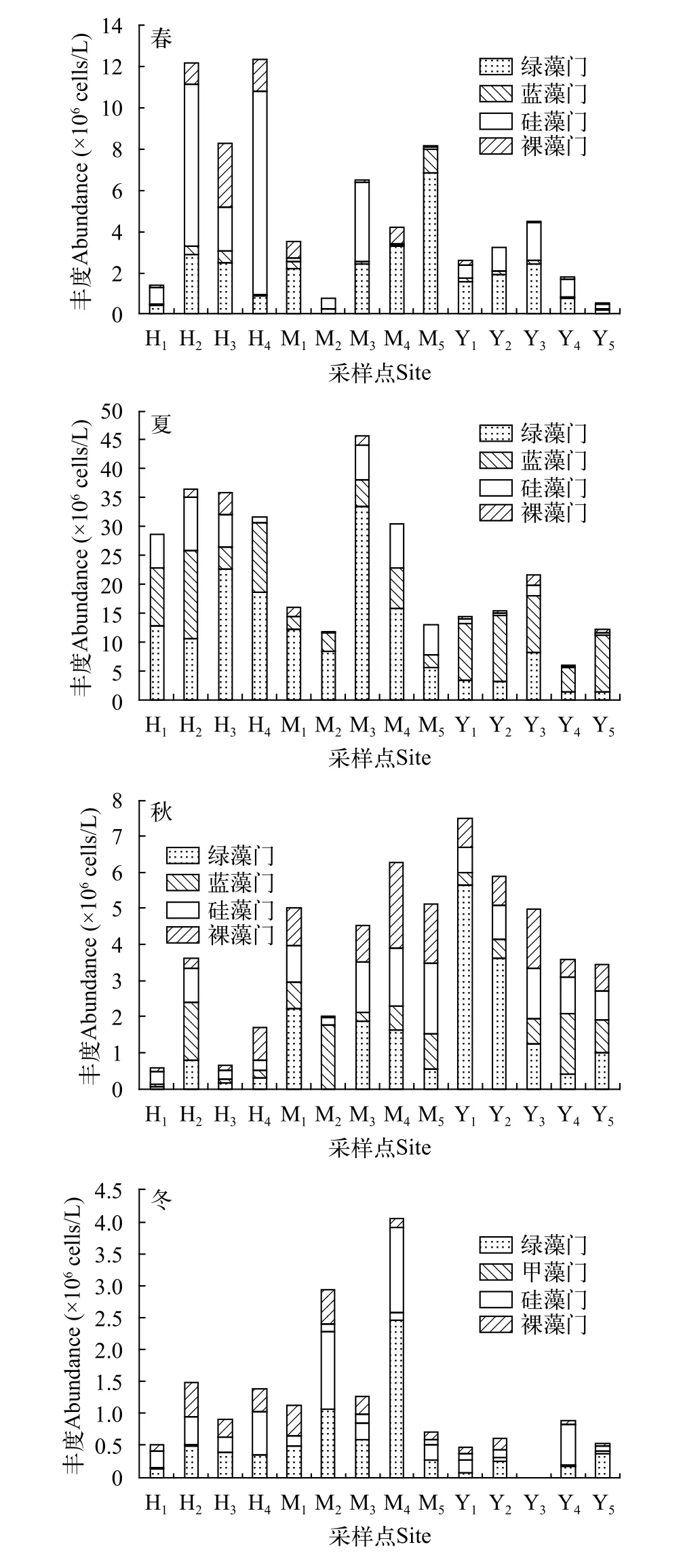
圖2 長湖浮游植物優勢類群豐度變化Fig. 2 Variation of abundance of phytoplankton dominant group in Lake Changhu
2.2 優勢種生態位特征
對長湖浮游植物優勢種生態位分析可知(圖3),生態位寬度變化范圍為1.002—2.377, 其中夏季小球藻最高, 其余大于2的種類分別是春季四尾柵藻、扭曲小環藻、顆粒直鏈藻, 夏季類顫魚腥藻、兩棲顫藻, 秋季類顫魚腥藻、兩棲顫藻、小球藻、帽形菱形藻以及冬季裸甲藻, 以上11種可以劃歸為廣生態位種類。平均邊緣指數(OMI)變化范圍為0.25—3.71, 其中小于1的物種共12種。耐受指數變化范圍為0.13—3.35, 夏季兩棲顫藻最高, 其次是夏季類顫魚腥藻(3.08)、冬季裸甲藻(2.67)和春季小球藻(2.41)。
通過對P值小于0.05的優勢種進行OMI分析結果表明, 共有16種優勢種的生態位分化較為顯著。對優勢種生態位和生態位寬度在第一軸和第二軸上分布情況的OMI分析表明(圖4), 優勢種和環境因子在各個季節的分化程度差異較明顯。
2.3 優勢種生態位重疊
優勢種生態位重疊值的變化范圍為0.02—0.93(圖5), 春季肘狀針桿藻和梭形裸藻以及秋季扭曲小環藻和梭形裸藻的重疊值均為最高值0.93。生態位重疊值超過(含)0.90的還有春季顆粒直鏈藻和梭形裸藻(0.91), 秋季湖泊色球藻和四尾柵藻(0.92)、類顫魚腥藻和微小平裂藻(0.92)以及冬季小球藻和鐮形纖維藻(0.90)、小球藻和扭曲小環藻(0.91)。生態位重疊值處于0.80(含)—0.90的共有19組優勢種。
2.4 浮游植物優勢種與環境因子的CCA分析
先將物種數據進行平方根轉化再進行除趨勢對應分析(DCA), pH以外的所有理化數據進行lg(x+1)轉換, 各個季節4個排序軸的最大梯度皆處于3—4之間, 選用CCA單峰模型進行排序。選取溶氧(DO)、水溫(WT)、酸堿度(pH)、透明度(SD)、總磷(TP)、總氮(TN)、氮磷比(N∶P)共7個環境因子與浮游植物優勢種進行CCA排序(圖6), 四個季節在軸一、軸二的特征值分別為0.632和0.143(春季)、0.591和0.286(夏季)、0.585和0.182(秋季)以及0.761和0.187(冬季), 其對應的解釋度分別為73.6%和16.7%(春季)、74.2%和16.4%(夏季)、69.4%和21.7%(秋季)以及72.4%和17.8%(冬季), 再經蒙特卡羅檢驗(Monte Carlo test)第1軸和第2軸均P<0.05, 表示CCA排序結果可信。
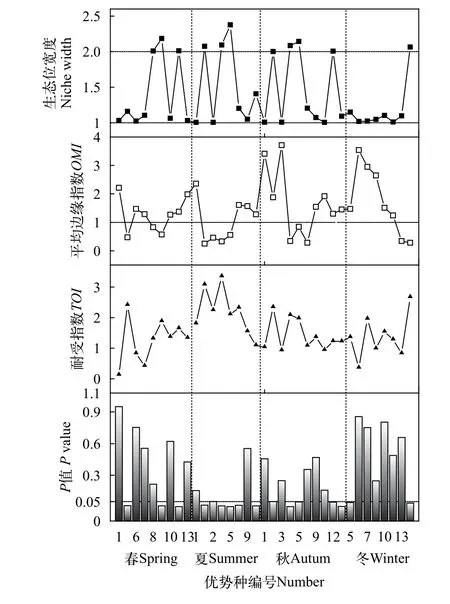
圖3 浮游植物優勢種生態位參數Fig. 3 Niche index of dominant species of phytoplankton
結合各環境因子與排序軸相關性分析可知, 春季小球藻與pH呈正相關, 鐮形纖維藻與TN有較大的相關性, 湖泊色球藻、四尾柵藻、扭曲小環藻、顆粒直鏈藻和梭形裸藻與環境因子的相關性都很低。夏季微小平裂藻、兩棲顫藻、四尾柵藻、扭曲小環藻以及梭形裸藻分布在排序軸的右下方, 表現出與TN和TP呈較強的正相關性, 與其他環境因子相關性較低。秋季湖泊色球藻、類顫魚腥藻、微小平裂藻、小球藻、四尾柵藻、扭曲小環藻、肘狀針桿藻、帽形菱形藻和梭形裸藻共計9種優勢種表現出與pH和DO有正相關性。冬季肘狀針桿藻與SD有較大正相關性, 梭形裸藻與pH有較大正相關性。
3 討論
3.1 優勢種群落結構特征
浮游植物的種類組成、豐度及分布等群落結構特征對水域環境具有較強的指示作用, 種類數量變化越小、豐度越高, 其富營養化程度就越高[25]。本次調查結果顯示綠藻門占比最高(50.8%), 其次為硅藻門(20.8%)和藍藻門(15.0%), 表明長湖屬典型綠藻門為主的水域類型, 浮游植物種類組成單一,多樣性較差, 群落結構不穩定。有研究發現[26]當水體受到有機污染時會導致金藻減少甚至消失, 而此次調查未發現金藻門種類, 表明除富營養化程度較高外, 長湖還存在受到有機污染的可能性。
本次調查共鑒定出優勢種14種。其中出現頻率為100%的種類有春季小球藻、鐮形纖維藻和四尾柵藻以及夏季湖泊色球藻、類顫魚腥藻、微小平裂藻和小球藻, 以上種類均為富營養化水體的常見種。從優勢度來看, 夏、秋、冬3季優勢度指數最高的種類均為小球藻, 表明多優勢度較集中, 多樣性較差。優勢度較高的種類將會占據更多的生態位, 種間競爭作用降低, 從而導致群落結構穩定性較差, 水體中發生水華的風險將會增加[25]。
從浮游植物豐度的平面分布來看, 各采樣點優勢種的豐度均超過了106cells/L, 參照湖泊富營養化評價標準[27]表明長湖處于富營養化狀態。從4個季節的總豐度來看, 呈現出海子湖、馬洪臺和圓心湖逐漸降低的分布特征, 這與各湖區的地理環境特點緊密相關。海子湖呈狹長型, 且隨著農村城鎮化發展, 沿岸帶人為因素干擾不斷增多, 此外與之相連多個進出水口也可增加該水域營養鹽的水平, 以上諸多因素均可影響浮游植物豐度的累積。馬洪臺和圓心湖為開闊水域, 較大面積的圍網圍欄養殖會造成水體交換率低、營養鹽水平升高等現象, 從而導致浮游植物豐度較高, 水體呈富營養化狀態。
3.2 優勢種生態位特征分析
生態位寬度是衡量生物利用各種資源的一個指標, 可反映出物種的分布和數量, 依據其大小可分為狹生態位和廣生態位[28]。生態位越寬, 表示物種對環境中各種資源的利用能力越強, 其在環境中的競爭能力就越強, 即為廣布種或是泛化種, 反之則為特化種。從本次調查結果來看, 生態位寬度大于2的種類共11種(表2), 如夏季小球藻(2.377)、兩棲顫藻(2.092)和類顫魚腥藻(2.075)等, 可劃歸為廣生態位種類。而生態位寬度較小的種類主要如夏季湖泊色球藻(1.005)和微小平裂藻(1.004)等可劃歸為狹生態位種類。有研究[29]發現優勢種生態位寬度側重于物種和資源位點的數量, 物種與分布站點密切相關。這與本次調查結果表現出了較強的一致性, 廣生態位的優勢種豐度值較高, 分布站點也較多, 從而獲得較高的生態位寬度, 說明對生境有較強的適應性, 這也與長湖夏季常出現區域性綠藻水華(主要是小球藻)一致。
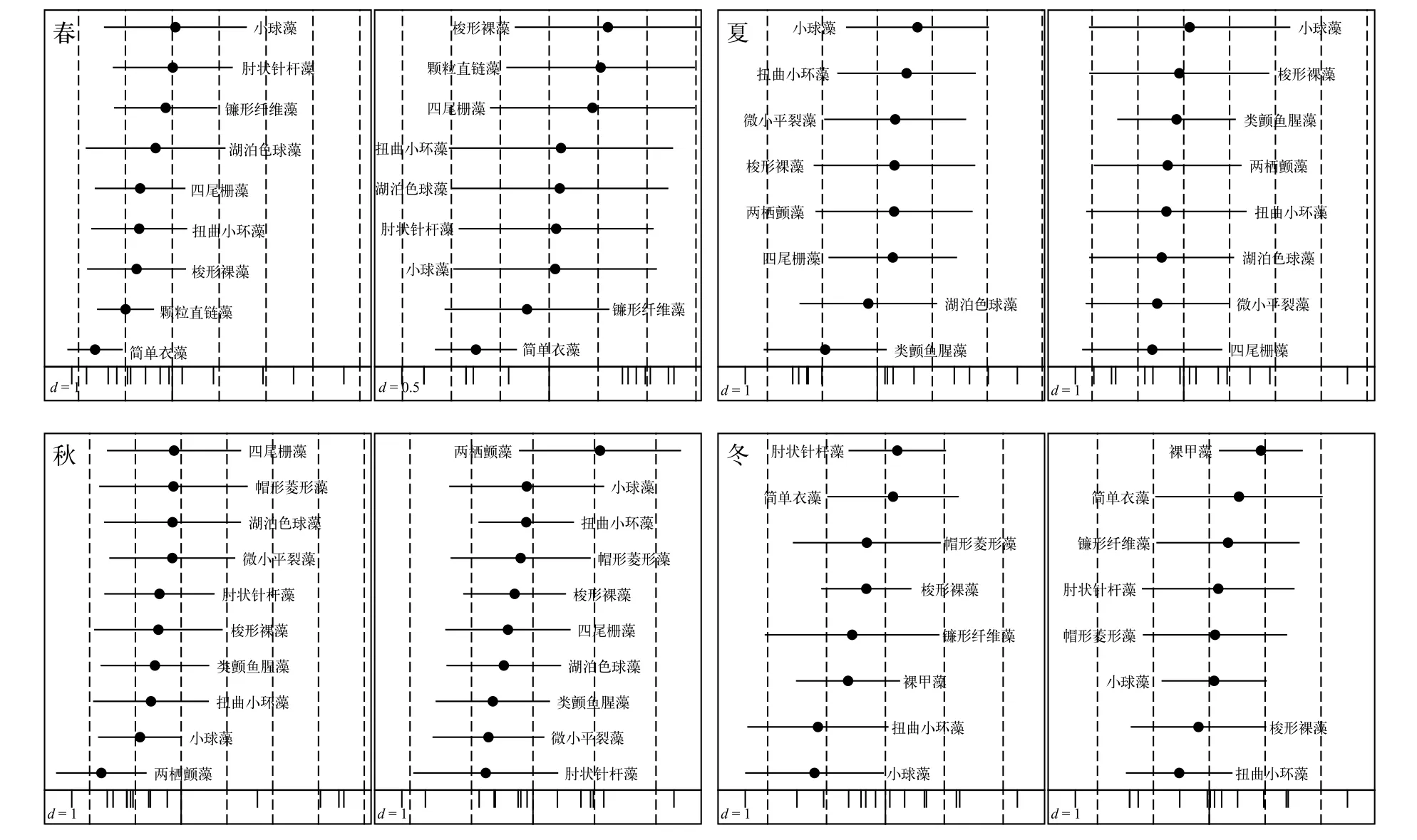
圖4 四季優勢種生態位和生態位寬度在平均邊緣指數分析第一軸和第二軸的分布Fig. 4 Niche position and niche breadth distribution on the first two axes of OMI analysis in each season
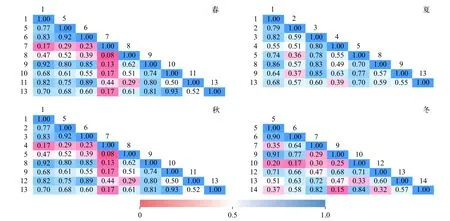
圖5 四季優勢種群間優勢種重疊值Fig. 5 Seasonal niche overlaps among dominant species in each season
平均邊緣指數可反映出區域內物種利用資源的能力, 通過OMI值的大小可將物種劃分為邊緣物種和非邊緣物種[30]。本次調查中OMI值較低(小于1)的種類共12種(圖2), 利用資源能力較強, 可劃歸為非邊緣物種。通過顯著性檢驗(P<0.05)的OMI分析發現, 除春、秋四尾柵藻以及冬季梭形裸藻外,其余9種OMI值較低的優勢種均出現了較明顯的生態位分化。以夏季為例,OMI值較小的是類顫魚腥藻(0.25)、微小平裂藻(0.46)、兩棲顫藻(0.33)和小球藻(0.56), 同時這幾種也是生態位較寬的藻類, 可能通過分泌化感物質和水生植物競爭從而獲得較強的競爭能力, 抑制其他藻類生長, 也可能是對生境特殊因子的較強適應能力從而占有更多的環境資源。
耐受指數側重于反映物種對環境的適應能力,其TOI值的大小可反映出物種對環境適應能力的強弱[31]。通常來看, 耐受性較強的種類生態位寬度也較大, 如TOI值較高的夏季顫藻(3.25)、類顫魚腥藻(3.08)和冬季裸甲藻(2.67), 它們的生態位寬度均超過2。
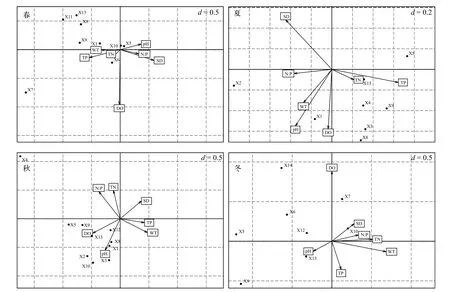
圖6 各季節優勢種與環境因子之間的典范對應分析Fig. 6 Canonical correspondence analysis between dominant species and environmental factors in different seasons
生態位重疊反映了物種間分享或競爭共同資源的現象, Wathne等[32]認為生態位重疊值大于0.6表示種間生態位重疊較顯著。本研究發現生態位重疊值超過0.90的共有7組優勢種, 超過0.80的共有19組優勢種。這19組優勢種的種間生態位重疊顯著, 反映出這些物種對資源利用的方式相似, 可能在棲息水層、溶氧、溫度、光照、水流、營養鹽水平等方面有相似的需求。一些硅藻如菱形藻、小環藻、針桿藻等屬浮游和底棲(附生)的兼性浮游植物, 在富營養化水體中一年四季都能形成優勢種群, 以春季和秋季最盛。裸藻多喜歡生活在有機質豐富的靜水水體中, 溫暖季節常大量繁殖成優勢種形成膜狀或云彩狀水華, 是水肥的標志[21]。因此在適溫季節, 小環藻與裸藻對環境資源的需求比較相似, 極易出現生態位重疊顯著的情況, 這也與本研究結果中春季肘狀針桿藻和梭形裸藻以及秋季扭曲小環藻和梭形裸藻重疊值最高(0.93)一致。此外,種間對于資源利用的競爭還受到資源供應的影響,資源供應短缺時競爭程度會趨于加強。
從本研究整體來看, 各優勢種生態位參數表現出了較好的一致性。以四季共有優勢種小球藻為例, 在夏秋兩季, 其生態位寬度均超過2, 屬廣生態位種類, 平均邊緣指數均小于1, 屬非邊緣物種, 耐受指數也較高, 表現出對環境較強的適應能力。以上表明小球藻生態位寬度、平均邊緣指數和耐受指數具有較好的一致性。
3.3 優勢種與環境因子的關系
CCA排序可以分析不同物種與環境因子的內在關系, 反映出物種對環境的需求程度[2]。而物種對環境的利用能力也是決定其生態位寬度的重要因素, 環境因子的改變將導致物種對資源的利用分化, 從而使物種間的生態位重疊發生改變[33]。從本次浮游植物優勢種與環境因子的CCA分析可以看出, 夏季常見富營養化水體指示種如兩棲顫藻、四尾柵藻和梭形裸藻等種類與TN和TP具有較強的正相關性, 而高溫和高營養鹽水平正是富營養化湖泊在夏季水華爆發的主要原因之一, 表明長湖與其他長江中下游典型富營養化淺水湖泊的研究一致[29]。春季水溫由低升高, 此時小球藻生態位寬度適中,而平均邊緣指數較低, 耐受指數較高, 表現出了對低溫較強的適應性, 這與王菁等[34]的研究結果一致。秋季水溫開始較夏季有所回落, 此時硅藻門帽形菱形藻表現出較強的環境適應能力, 與pH有較大正相關性, 生態位寬度和耐受指數均較高。此外,pH、DO、N/P以及SD也是影響長湖浮游植物優勢種分布的重要環境因子, 這與長湖地理形狀、地貌特征及其他因素如水流速度、降雨量、進出水口及水草分布等有關。
3.4 長湖浮游植物生態位的演替
本次調查是繼2012年[11]以來第二次對長湖浮游植物生態位的系統研究, 通過比較發現, 長湖浮游植物的種類組成、優勢種以及豐度均沒有明顯變化, 種類組成仍以綠藻門和藍藻門為主, 優勢種也主要是富營養化水體常見種類, 其中小球藻和扭曲小環藻在兩次調查中均為四季共有優勢種。各站點豐度均超過106cells/L, 仍處于富營養化狀態。本次調查的豐度較2012年有升高趨勢, 其中冬季裸甲藻數量明顯增多, 為冬季優勢種, 而裸甲藻并未出現在2012年的優勢種中。根據生態位寬度而劃分的廣生態位種類和狹生態位種類數量有所增加, 廣生態位種類仍以小球藻和扭曲小環藻為代表, 又增加了兩棲顫藻、類顫魚腥藻和裸甲藻等種類, 這些種類通常是富營養化水體的常見優勢種,表明長湖水域仍處在富營養化狀態, 這與浮游植物種類組成、優勢種以及豐度所對長湖的評價結果一致。本研究增加了平均邊緣指數和耐受指數分析, 對生態位寬度和生態位重疊進行物種水平分布的測度分析, 揭示一定環境梯度下物種的生態位分布, 并通過種間關系深入分析浮游植物的生態位變化, 也較好地反映了物種對所在環境的適應性和對資源的利用能力。以四季共有優勢種扭曲小環藻、小球藻和梭形裸藻為例, 可劃歸為廣生態位種類, 屬非邊緣物種, 耐受指數也較高, 表現出對環境較強的適應能力, 這些優勢種的存在對于湖泊的富營養化治理以及水華的預防都是較難解決的問題。通常認為外部環境因子改善和內部生物種群恢復二者有機結合是恢復湖泊健康生態系統的重要途徑, 主要包括控源截污、高等水生植物尤其是沉水植物的種群恢復以及大型濾食性底棲動物的增加等, 上述生態修復措施可以更好地發揮種間競爭和密度制約機制作用下的生態系統組織功能, 實現抑制先鋒種或優勢種的生長與擴散, 促進其他邊緣物種的生長與繁殖, 最終形成良性平衡狀態的湖泊生態系統[25,27]。
建議長湖下一步增加對各進出水口營養鹽水平和水動力條件等監測并同時開展全面生態調查如濾食性魚類的攝食壓力、高等水生植物、底棲生物及周叢生物的研究, 充分考慮多種環境因子如水文和高等水生植物的克藻效應等措施以控制或減緩藻類水華暴發, 是較好的生態修復手段[35,36]。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
趣味(語文)(2020年3期)2020-07-27 01:42:46
中國生殖健康(2020年6期)2020-02-01 06:28:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
