豬繁殖與呼吸綜合征病毒華南株GDjm全基因組測序與遺傳進化分析
2020-06-08 03:33:18張馳董建國于林洋王爽云范蘭蘭張樂宜劉燕玲王磊宋長緒
畜牧與獸醫 2020年6期
張馳,董建國,于林洋,王爽云,范蘭蘭,張樂宜,劉燕玲,王磊,宋長緒*
(1. 華南農業大學動物科學學院,廣東 廣州 510642;2. 信陽農林學院牧醫工程學院,河南 信陽 464000)
豬繁殖與呼吸綜合征(porcine reproductive and respiratory syndrome,PRRS)可引起任何品種、性別、年齡的豬發病,其中易感性最高的是母豬和仔豬,且感染后出現嚴重的臨床癥狀[1]。PRRS是導致母豬發熱、厭食、早產、流產、死胎、弱仔等繁殖障礙和仔豬呼吸道癥狀為主要特征的傳染病[2]。目前豬繁殖與呼吸綜合征病毒(PRRSV)分為2個基因型:基因型1和基因型2,代表性毒株分別為歐洲型Lelystad Virus和美洲型VR-2332[3]。中國內陸于1995年首次發現PRRSV,并在1996年成功分離出第1株PRRSV毒株CH-1a[4];2006年,中國南方地區出現由變異性毒株引起的高發病率和高死亡率的疫情[5-6],隨后蔓延至全國大部分地區,給我國養豬業造成了嚴重的經濟損失。
PRRSV為單股正鏈RNA有囊膜病毒,基因全長約為15~15.5 kb[7],至少含有10個開放閱讀框(ORF),包括ORF1a、ORF1b、ORF2a、ORF2b、ORF3、ORF4、ORF5、ORF5a、ORF6和ORF7[8]。在ORF1a中的NSP2基因最容易變異,主要含3個區域:N末端的半胱氨酸酶區、中部的高變區和C末端疏水的轉膜區,NSP2發生的變異主要位于中部的高變區[9]。NSP2區域在許多PRRSV分離毒株中存在氨基酸缺失,這是PRRSV基因組長度差異的一個主要原因。ORF5編碼的糖基化囊膜蛋白GP5有較強的免疫原性,可誘導機體產生中和抗體,常被作為構建PRRSV診斷的重要靶標[10]。GDZS2016為近年來廣東地區流行毒株,與JXA1毒株在同一分支,相關文章報道其能造成臨床發病率為90%,死亡率為10%,不具有感染Marc-145細胞的能力,但可感染PAM細胞[11]。PRRSV發生基因重組的頻率很高,可能導致新型高致病性豬繁殖與呼吸綜合征病毒(HP-PRRSV)的出現[12-13]。
2018年7月,本實驗室從廣東省江門市某PRRS發病豬場采集肺臟組織樣本,PCR檢測結果為PRRSV陽性,在Marc-145細胞上無法分離純化,最終在原代豬肺泡巨噬細胞(porcine alveolar macrophages,PAM)上成功分離,將其命名為GDjm株。對全基因組進行測定,將測序結果與其他代表PRRSV毒株進行序列比對和進化樹分析,進而分析GDjm毒株與國內流行的PRRSV毒株的相似性和重組情況,為深入研究該病毒株的遺傳與變異奠定了基礎。
1 材料與方法
1.1 病料
肺臟病變組織來源于廣東省江門市疑似發生PRRS某養殖場,-20 ℃凍存備用。
1.2 細胞、菌體
Marc-145細胞、PAM細胞、大腸桿菌感受態細胞,由華南農業大學國家生豬種業工程技術研究中心種豬疾病防控實驗室保存。
1.3 主要試劑
病毒DNA/RNA提取試劑、反轉錄試劑盒以及膠回收試劑盒盒購自Promega公司;RT-PCR相關試劑盒購自Vazyme公司;DNA Marker DL2000等購自廣州東盛生物科技有限公司;EB替代染料(GoldviewⅠ)購自廣州華奇盛生物科技有限公司;pEASY-Blunt Simple Cloning Vector購自北京全式金生物技術有限公司。大腸桿菌感受態細胞購自天根生物科技(北京)有限公司。
1.4 引物的設計與合成
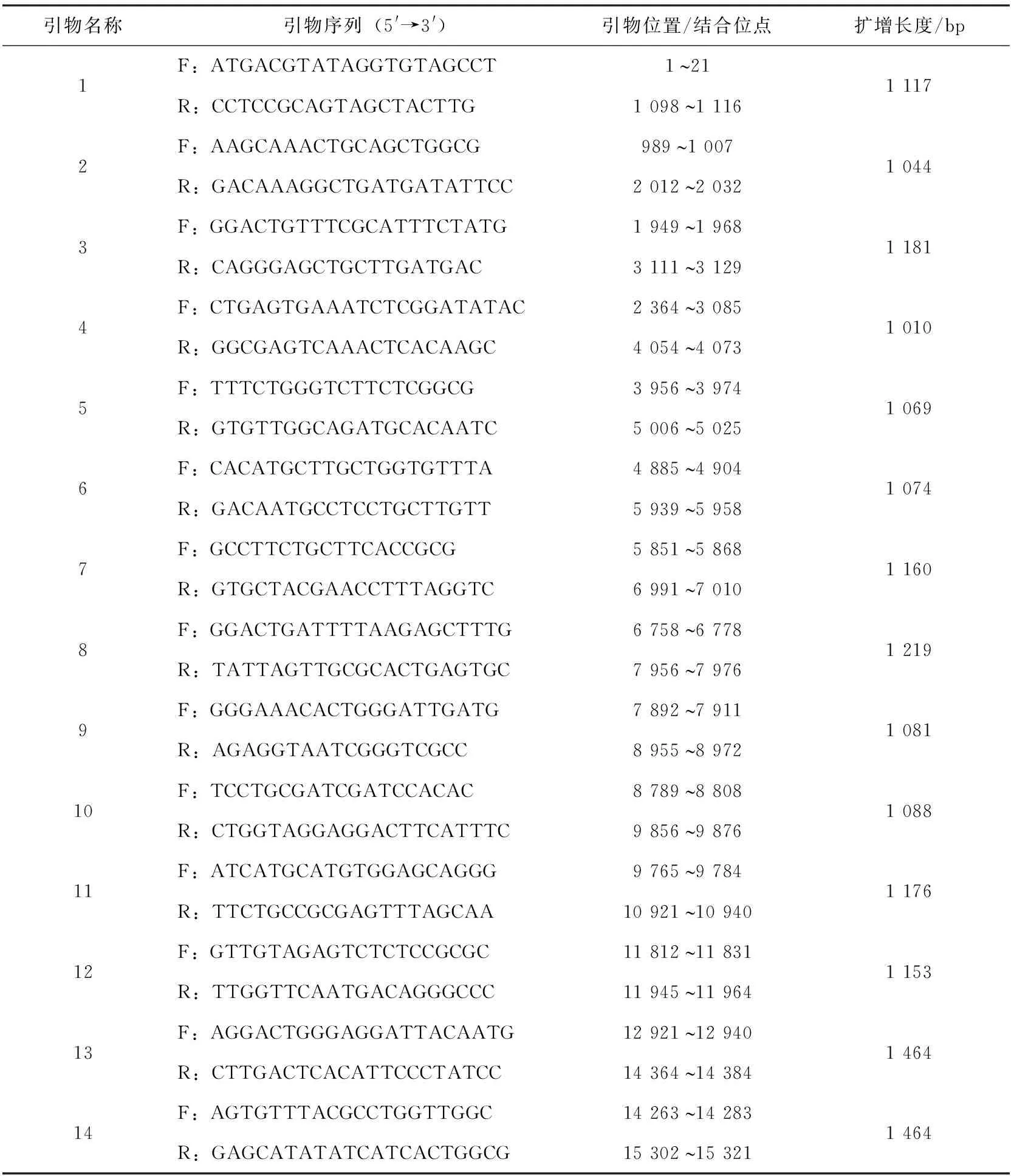
根據GenBank中發布的PRRSV毒株全基因組序列進行生物信息學分析,選取保守區域設計14對特異性引物(表1)用于擴增序列,引物由生工生物工程(上海)股份有限公司合成。
1.5 病料處理
取1 g肺臟組織剪碎,加入1 mL PBS在冰上充分研磨。將研磨液倒入潔凈的1.5 mL 離心管中,12 000 r/min離心3 min,收集上清后在超凈工作臺中用0.22 μm微孔過濾器過濾,取200 μL 濾液用于病毒核酸提取,并通過PCR進行豬繁殖與呼吸綜合征病毒(PRRSV)、豬瘟病毒(CSFV)、豬流行性腹瀉病毒(PEDV)、豬細小病毒(PPV)檢測,其余濾液置于-80 ℃保存備用。
1.6 病毒分離
采用PAM細胞和Marc-145細胞從樣品中分離PRRSV。將處理好的臨床樣品感染PAM和Marc-145細胞,盲傳3代,并觀察細胞病變效應(cytopathic effect,CPE)。收毒后反復凍融3次,每代取200 μL細胞培養液于無RNA酶的離心管中,用于提取核酸和PCR檢測。
表1 擴增全基因序列引物
1.7 間接免疫熒光試驗(IFA)
將PAM細胞和Marc-145接至24孔細胞培養板中,置于細胞培養箱中培養24 h后棄掉培養液,PBS緩沖液洗滌3次。每孔接種10 μL上述濾過液,設立空白對照,放在于細胞培養箱中孵育1 h后棄掉液體,PBS輕輕沖洗3次,各孔加入500 μL含有2% FBS的細胞維持液,觀察培養25 d后棄掉培養液,PBS洗滌3次,各孔加進200 μL固定液于-20 ℃固定20 min,棄掉液體,PBS洗滌3次;然后各孔加入200 μL 0.5% Triton X-100室溫穿膜10 min,棄掉液體后PBS洗滌3次;加入200 μL的1∶400倍稀釋的一抗(鼠源抗N蛋白單抗),置于37 ℃孵育1 h,期間每20 min輕輕搖晃1次,確保充分吸附,棄去液體后PBS洗滌5次;避光加入200 μL稀釋后的熒光二抗(FITC標記的羊抗鼠IgG),37 ℃條件下避光作用1 h,PBS洗滌5次,置于熒光顯微鏡下進行觀察。
1.8 提取核酸及反轉錄
將收集的上清液取200 μL,采用Promega公司提供的試劑盒進行病毒核酸提取和反轉錄,反轉錄產物置于-20 ℃冰箱保存。
1.9 目的基因分段擴增
采用TAKARA公司提供的高保真酶對目的片段進行擴增,PCR擴增體系為50 μL,PrimeSTAR Max Premix (2×) 0.5 μL、5× PrimeSTAR Buffer 10 μL、dNTP Mixture 4 μL、RNase-free H2O 27.5 μL、cDNA模版4 μL,上下游引物各2 μL。PCR擴增程序:98 ℃預變性1min;98 ℃變性10s,53 ℃退火15 s,72 ℃延伸2 min,30個循環;72 ℃再延伸10 min。取6 μL PCR產物進行1%瓊脂糖凝膠電泳。
1.10 基因克隆、測序
PCR擴增產物經過電泳鑒定后,使用Promega公司提供的膠回收試劑盒進行膠回收。取4 μL膠回收產物1 μL pEASY-Blunt Simple Cloning Vector 25 ℃連接25 min,轉化后挑菌鑒定,將陽性菌液送往生物公司測序。
1.11 序列分析
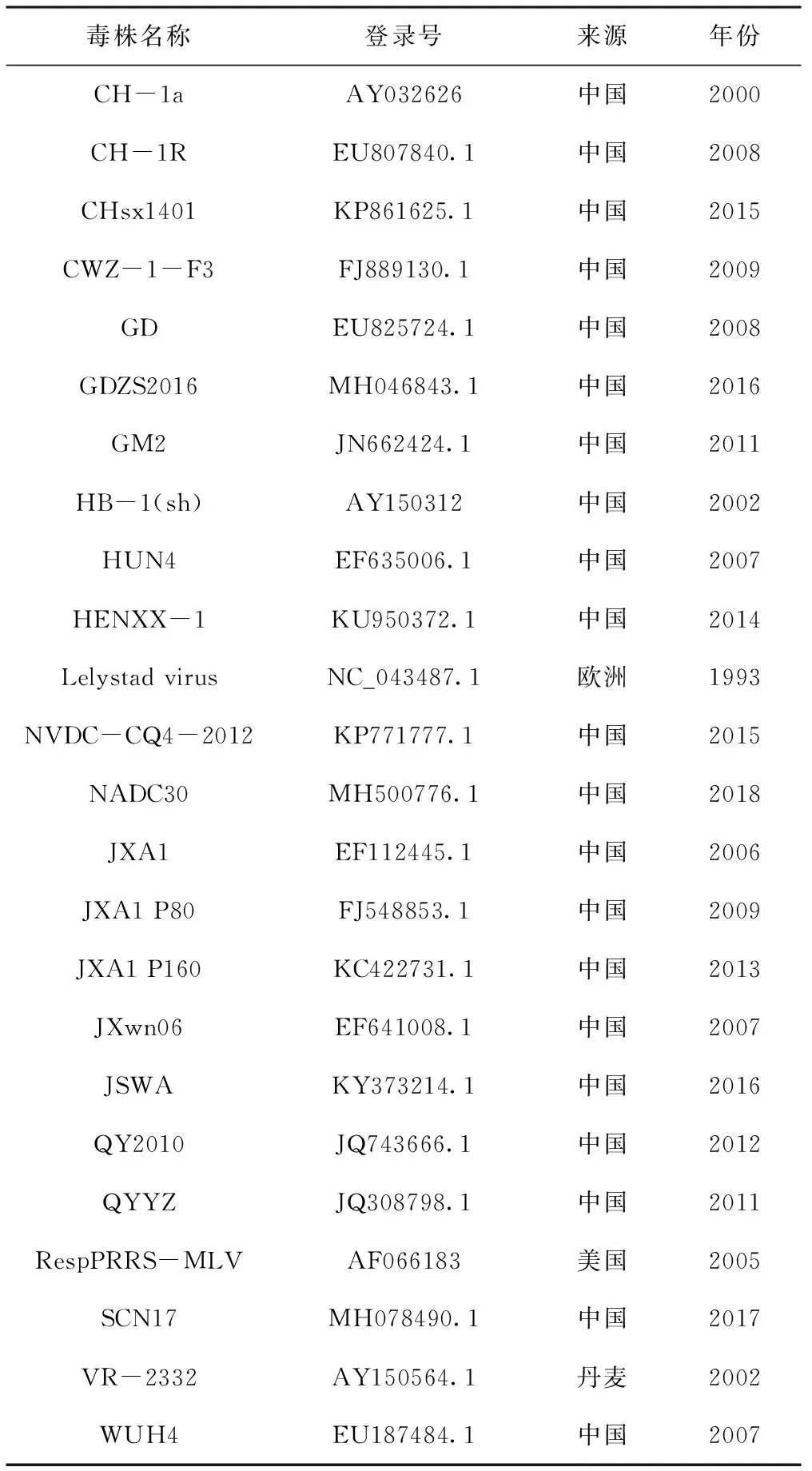
表2所示PRRSV代表毒株基因序列均來自GenBank。運用Lasergene與Mega 5.0軟件將測序毒株與國內外代表毒株進行序列比對和進化樹構建,并將GDjm毒株NSP2與GP5基因氨基酸序列與國內外代表毒株進行比對分析。通過SimPlot軟件對GDjm株進行全基因重組分析。
表2 PRRSV參考毒株信息
毒株名稱登錄號 來源年份CH-1aAY032626中國2000CH-1REU807840.1中國2008CHsx1401KP861625.1中國2015CWZ-1-F3FJ889130.1中國2009GDEU825724.1中國2008GDZS2016MH046843.1中國2016GM2JN662424.1中國2011HB-1(sh)AY150312中國2002HUN4EF635006.1中國2007HENXX-1KU950372.1中國2014Lelystad virusNC_043487.1歐洲1993NVDC-CQ4-2012KP771777.1中國2015NADC30MH500776.1中國2018JXA1EF112445.1中國2006JXA1 P80FJ548853.1中國2009JXA1 P160KC422731.1中國2013JXwn06EF641008.1中國2007JSWAKY373214.1中國2016QY2010JQ743666.1中國2012QYYZJQ308798.1中國2011RespPRRS-MLVAF066183美國2005SCN17MH078490.1中國2017VR-2332AY150564.1丹麥2002WUH4EU187484.1中國2007
2 結果與分析
2.1 病料的RT-PCR檢測
使用ORF5基因檢測引物對樣品進行擴增,1%瓊脂糖凝膠電泳分析結果表明(圖 1),檢測結果與預期結果(864 bp)相符。
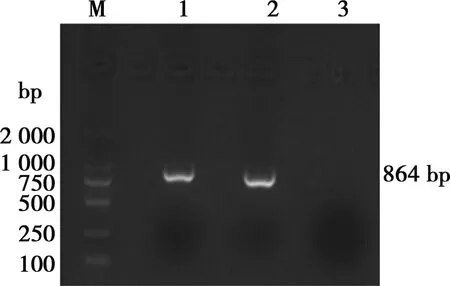
M. DNA Marker DL2000;1. 樣品;2. 陽性對照;3. 陰性對照
圖1 PRRSV ORF5基因片段擴增電泳結果
2.2 病毒分離與IFA
在Marc-145細胞多次接毒后未出現CPE,隨后將處理后的病料分別接種于Marc-145細胞與PAM細胞,培養48 h后進行間接免疫熒光檢測N蛋白。如圖2所示,Marc-145細胞無熒光,PAM細胞上有明顯的熒光信號,證明分離毒可感染PAM細胞但不感染Marc-145細胞。
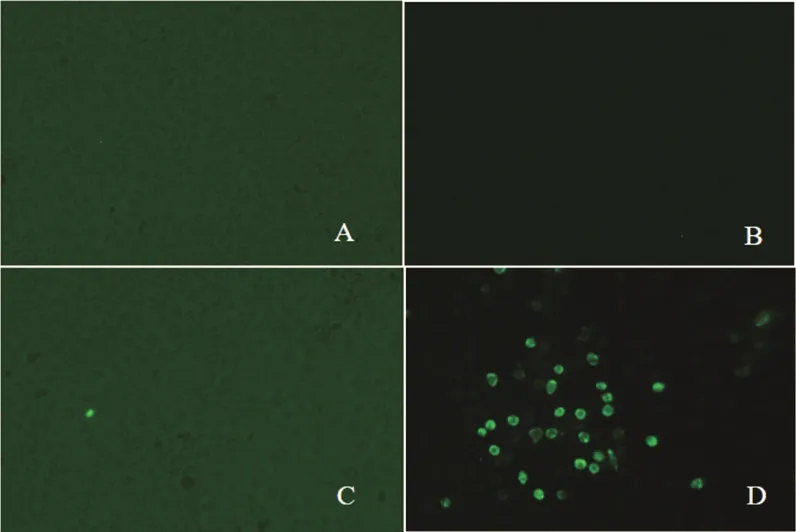
A. 未接毒Marc-145細胞;B. 接毒Marc-145細胞;C. 未接毒PAM細胞;D. 接毒PAM細胞
圖2 間接免疫熒光檢測結果
2.3 PRRSV GDjm全基因組測定
使用14對相互重疊的特異性引物擴增全基因。瓊脂糖凝膠電泳分析顯示,擴增的14個產物大小與預期一致(圖 3)。將擴增片段膠回收連接載體后測序,使用SeqMan軟件拼接序列,獲得全基因序列全長為15 249 bp,命名為GDjm株。
M.DNA Marker DL2000;1~14. PCR擴增的14個片段
圖3 PRRSV GDjm全基因RT-PCR擴增瓊脂糖凝膠電泳
2.4 PRRSV GDjm基因進化樹構建與同源性分析
應用DNAStar軟件對GDjm毒株與國內外代表性毒株進行全基因組進化樹構建,結果如圖4所示。遺傳進化樹分為歐洲型代表毒株Lelystad virus和美洲型代表毒株VR-2332兩大分支,以VR-2332為代表的美洲型又可進一步分為以JXA1、JXwn06為代表的高致病毒株分支,以我國CH-1a與VR-2332為代表的低致病性毒株分支,以及美國NADC30為代表的分支和近年來出現的QYYZ為代表的分支。如圖所示,GDjm株與華南地區近年新出現的重組毒GDZS2016同源性最高,并位于JXA1、JXwn06等為代表的高致病性毒株和以CH-1a為代表的經典毒株中間。故判定GDjm株可能為重組毒株。
利用DNAStar軟件對PRRSV全基因核苷酸序列分析,結果如圖5所示,GDjm與GDZS2016、HUN4、NVDC-CQ4-2012、WUH4、JXA1的同源性分別為96.2%、96.0%、95.7%,95.5%、95.4%,與NADC30同源性為82%,與美洲型代表毒株VR2332同源性為87.4%,與歐洲型代表株Lelystad virus同源性僅為57.9%。
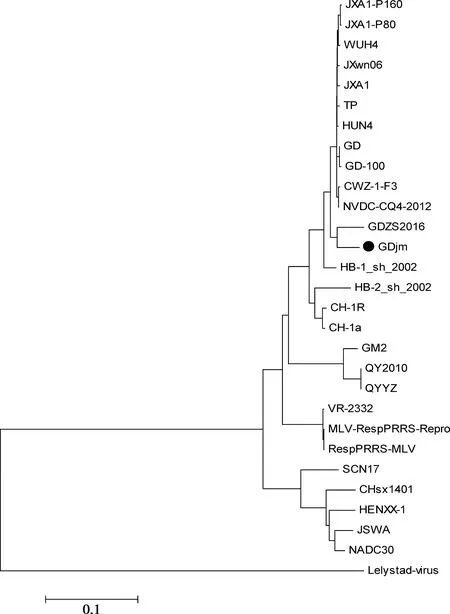
●標注毒株為所測毒株
圖4 PRRSV全基因組的進化樹分析

圖5 PRRSV-GDjm全基因序列比對結果
2.5 GDjm NSP2與GP5氨基酸序列比對及核苷酸同源性分析
運用DANStar對GDjm和國內代表典毒株進行NSP2氨基酸序列比對,結果如圖6所示,GDjm毒株在511、534~562位存在30個不連續氨基酸的缺失,此外在531位存在T531I突變。
運用DNAStar軟件對GDjm和國內外參考毒株進行GP5氨基酸序列比對(圖7)。GDjm在29位有A29V變異;在185位有V185A突變;在189位有I189L突變;在39位有1個L39I突變。此外GDjm還存在多個突變位點,在34位存在D34G突變,58位有N58E突變,107位有V107A突變,196位有Q196R突變。
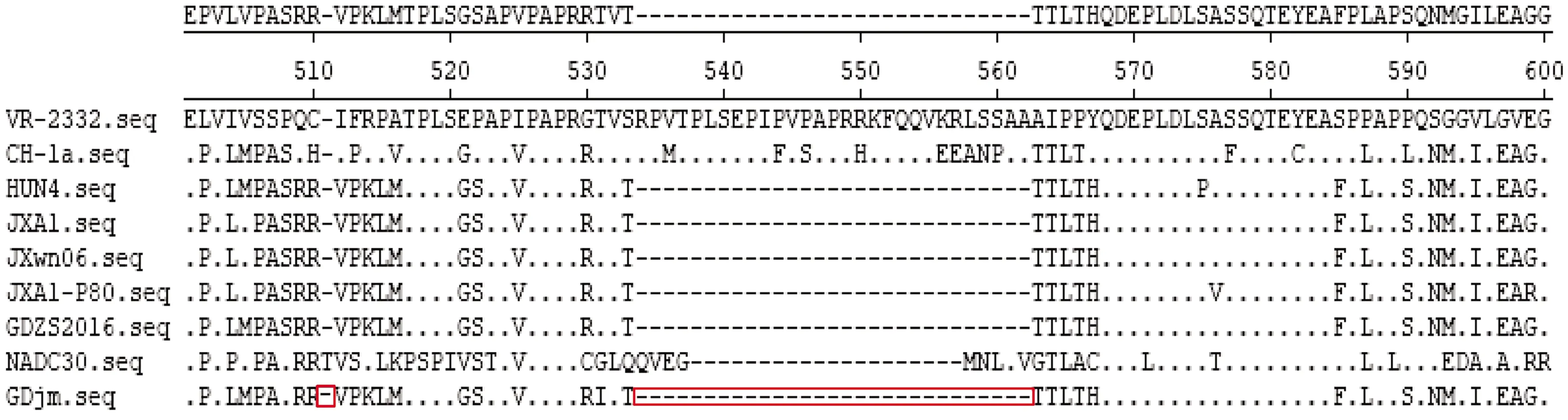
方框中為核苷酸變異位點,下同
圖6 PRRSV-GDjm NSP2氨基酸序列比對結果

圖7 PRRSV GDjm GP5氨基酸序列比對結果
2.6 PRRSV全基因重組部位特征分析
運用SimPlot軟件將GDjm株與高致病性毒株HUN4和近年來華南地區毒株GDZS2016進行全基因組重組分析。經分析得出(圖8):GDjm株為高致病毒株HUN4與GDZS2016進行重組的重組毒株。重組區域分別為6 723~9 521 bp、11 998~15 019 bp。
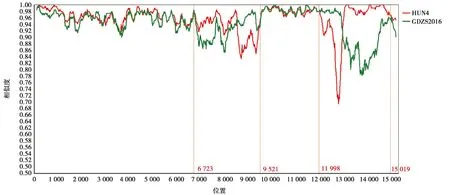
坐標尺:200 bp;區分度:20 bp
圖8 PRRSV GDjm毒株重組分析
3 討論
PRRS是全球養豬業的重大傳染病之一。我國分離得到的毒株以北美型PRRSV為主。防控PRRS的疫苗主要有經典毒株和高致病性毒株減毒活疫苗以及滅活疫苗[14]。我國PRRSV流行的種類眾多,研發和生產疫苗的廠家眾多,毒株的選用以及生產方式皆有不同,導致使用混亂。養殖場也可能在免疫流程中使用不同型號的疫苗,使毒株間發生遺傳重組的可能性增加,加上RNA病毒復制酶低保真性和自身免疫系統的影響[15],進而加速PRRSV變異。
本試驗對廣東省江門市疑似發生PRRS的豬場采集病料進行PCR檢測,結果呈陽性;通過對GDjm毒株全基因組測序,并對其與其他PRRSV毒株同源性進行比較和構建進化樹分析。全基因組分析中GDjm與GDZS2016同源性最高,為96.2%。NSP2是PRRSV最大的非結構蛋白,容易發生缺失和突變,基因長度約為2.9 kb。 GDjm毒株NSP2氨基酸序列比對結果與高致病性毒株HUN4、JXA1、JXwn06和高致病疫苗株JXA1-P80相似,在511、534~562位存在30個不連續氨基酸的缺失,符合高致病性毒株特點。美洲型毒株GP5蛋白的表位有3個,aa27~aa30和aa180~aa197為非中和表位,aa37~aa45為中和表位[16]。與美洲型代表毒株VR-2332相比,GDjm在非中和表位中有1個A29V的變異;在185位有V185A突變,該突變與高致病性毒株及其疫苗株相同;在189位有I189L突變,該突變與經典毒株和高致病性毒株及其疫苗株相同。中和表位中,在39位有1個L39I突變,該突變與高致病性毒株及其疫苗株相同。
GDZS2016株是華南地區2016年出現的重組毒株,相關文章報道其能造成臨床發病率為90%,死亡率為10%,現研究表明該毒株只能感染PAM細胞,對Marc-145不具有感染性[17]。HUN4毒株是高致病毒株,臨床試驗表明其能造成發病率和死亡率高達100%,均能使Mare-145和PAM感染[18]。全基因重組部位特征分析表明HUN4與GDZS2016重組,重組區域8 558~9 462 bp、11 995~12 982 bp。這2個片段編碼病毒RNA復制酶、聚合酶GP2、GP3、GP4、GP5[19],它們的重組可能與GDjm毒株不能感染Marc-145細胞相關。綜上所述,GDjm毒株為HUN4與GDZS2016的重組毒株,具有致病性增強的潛在風險,加大了廣東地區PRRSV防控的難度,因此有必要開發新型疫苗預防疾病的發生,并早日解決PRRSV感染與流行的問題。
