天津周邊地區(qū)煙粉虱隱種鑒定及其系統(tǒng)發(fā)育分析
2020-06-03 03:31:53張鄧壯張艷玲胡明鑫劉佰明谷希樹(shù)
天津農(nóng)業(yè)科學(xué) 2020年3期
張鄧壯 張艷玲 胡明鑫 劉佰明 谷希樹(shù)
摘? ? 要:煙粉虱是一種世界性害蟲(chóng),入侵世界許多地區(qū),其攜帶的番茄黃化曲葉病毒(Tomato yellow leaf curl virus, TYLCV)給番茄生產(chǎn)造成了嚴(yán)重的損失。煙粉虱的種類繁多,給綠色防控帶來(lái)許多困難。為了明確煙粉虱的種類,本試驗(yàn)主要對(duì)天津周邊,還有山東、山西和湖北等省市10個(gè)地區(qū)不同寄主共16個(gè)煙粉虱種群進(jìn)行了隱種鑒定及系統(tǒng)發(fā)育分析。結(jié)果表明,其中13個(gè)煙粉虱種群只有MED隱種,3個(gè)煙粉虱種群中MEAM1和MED隱種共存。MEAM1隱種煙粉虱的同源性高達(dá)100%,MED隱種煙粉虱的同源性為99.6%~100%,入侵我國(guó)的MED隱種可能起源于地中海西部地區(qū)。本研究明確了監(jiān)測(cè)地區(qū)煙粉虱隱種及其進(jìn)化關(guān)系,為當(dāng)?shù)責(zé)煼凼姆揽靥峁┝丝茖W(xué)依據(jù)。
關(guān)鍵詞:煙粉虱;系統(tǒng)樹(shù);隱種;系統(tǒng)發(fā)育
中圖分類號(hào):Q966? ? ? ? ? ? 文獻(xiàn)標(biāo)識(shí)碼:A? ? ? ? ?DOI 編碼:10.3969/j.issn.1006-6500.2020.03.004
Species Identification and Phylogenetic Analysis of Bemisia tabaci in Tianjin and Surrounding Areas
ZHANG Dengzhuang1,2, ZHANG Yanling3, HU Mingxin2,4, LIU Baiming2, GU Xishu2
(1.College of Life Science, Tianjin Normal University, Tianjin 300387, China; 2.Tianjin Institute of Plant Protection, Tianjin 300384, China; 3.Vegetable Technology Promotion Station of Ninghe District, Tianjin City,Tianjin 301500, China;4.College of Modern Agriculture and Ecological Environment, Heilongjiang University, Harbin, Heilongjiang 150080, China)
Abstract: As a worldwide pest, Bemisia tabaci has invaded many regions of the world. The tomato yellow leaf curl (TYLCV) virus it carries has caused serious losses. There are many species of Bemisia tabaci, which brings many difficulties to green prevention and control. In order to clarify the species of Bemisia tabaci, in this experiment, a total of 16 species of Bemisia tabaci in different regions of Tianjin and its vicinity, Shandong Shanxi,Hubei were collected to perform species identification and phylogenetic analysis. The results showed that 13 populations only contained Bemisia tabaci MED, 3 populations contained both Bemisia tabaci MEAM1 and Bemisia tabaci MED. Among our collected samples, the homology of Bemisia tabaci MEAM1 was 100%, and The homology of Bemisia tabaci MED was 99.6%~100%.China's MED may originate in the western Mediterranean. This study illustrated the species and evolutionary relationships of Bemisia tabaci in the monitored areas, providing a scientific basis for the prevention and control of Bemisia tabaci in these areas.
Key words: Bemisia tabaci; phylogenetic tree; species; phylogenetic
煙粉虱Bemisia tabaci(Gennadius)是一種多食性農(nóng)業(yè)害蟲(chóng),能危害600多種寄主植物,為害方式包括通過(guò)吸取植物汁液、分泌蜜露影響光合作用直接危害植物,還可以通過(guò)傳播200多種植物病毒間接危害植物[1-2],番茄黃化曲葉病毒能侵染多種植物,在番茄上的為害尤為嚴(yán)重,嚴(yán)重時(shí)導(dǎo)致絕收[3]。煙粉虱有非常復(fù)雜的遺傳結(jié)構(gòu),目前認(rèn)為煙粉虱至少是由36個(gè)隱種組成的物種復(fù)合體[4-5]。其中MEAM1隱種(B型)和MED隱種(Q型)煙粉虱入侵性最強(qiáng)、為害最重。煙粉虱侵入我國(guó)時(shí)間較長(zhǎng),在20世紀(jì)90年代MEAM1煙粉虱就開(kāi)始入侵我國(guó),之后迅速蔓延到全國(guó)各地。而MED煙粉虱則入侵較晚,于2003年在云南首次發(fā)現(xiàn),但是在最近十年中MED煙粉虱已經(jīng)取代了MEAM1煙粉虱成為中國(guó)主要的煙粉虱類型[6-7]。不同煙粉虱隱種具不同的生物學(xué)特性,如MEAM1和MED隱種煙粉虱的傳播植物病毒能力和對(duì)殺蟲(chóng)劑抗性存在明顯差異[8-9]。MED隱種的遺傳多樣性要比MEAM1更加豐富,因而其適應(yīng)環(huán)境的能力更強(qiáng),能更好的抵御外界不利因素,例如農(nóng)藥、化學(xué)藥劑等,MED隱種對(duì)新煙堿類殺蟲(chóng)劑的抗性更強(qiáng)[10]。近年來(lái),國(guó)內(nèi)研究者已逐漸重視田間煙粉虱隱種的監(jiān)測(cè),監(jiān)測(cè)的地區(qū)包括新疆、北京、湖南、江蘇、陜西、湖北等,為科學(xué)防治煙粉虱提供了理論依據(jù)[11-15]。本研究主要立足于天津各區(qū)及其周邊地區(qū),同時(shí)與其他省份的地區(qū)作比較,采用煙粉虱mtDNA COI基因作為分子標(biāo)記,對(duì)天津各區(qū)、北京、山東、山西和武漢等地區(qū)不同寄主植株的煙粉虱種群進(jìn)行隱種鑒定及系統(tǒng)發(fā)育分析,旨在為合理的防治天津地區(qū)煙粉虱提供科學(xué)依據(jù)。
1 材料和方法
1.1 材 料
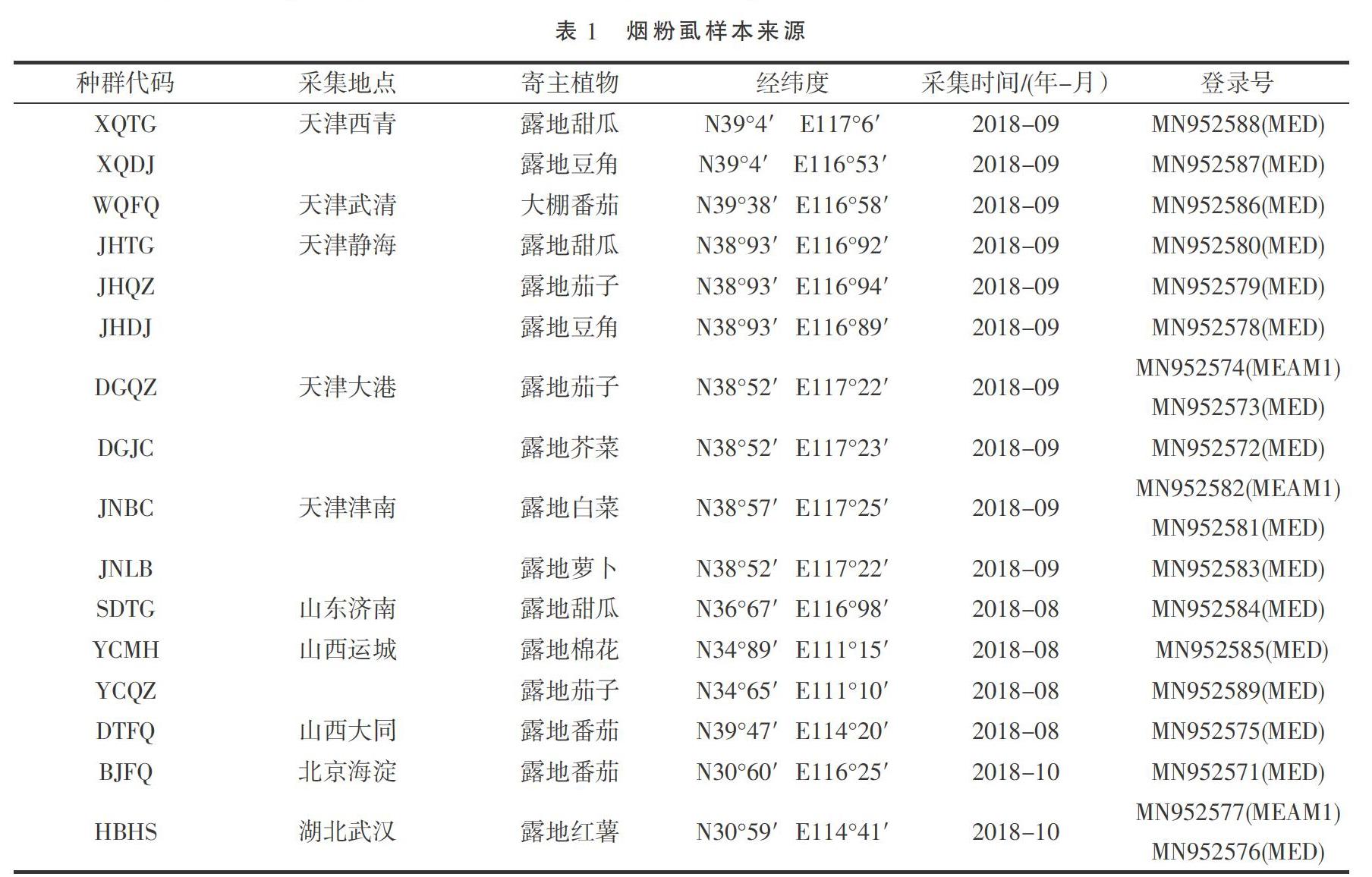
采集了天津(西青、靜海、大港、津南、武清)、北京海淀、山東濟(jì)南、山西運(yùn)城、山西大同以及湖北武漢共10個(gè)地區(qū)不同寄主的16個(gè)種群的煙粉虱成蟲(chóng),所采集到的樣品直接浸泡于1.5 mL裝有無(wú)水乙醇的離心管中,保存在-20 ℃冰箱中備用。具體采集信息及登錄號(hào)見(jiàn)表1。
1.2 方 法
1.2.1 單頭煙粉虱總DNA的提取 參照柳洋[16]的方法,使用KAPA DNA提取試劑盒對(duì)煙粉虱DNA進(jìn)行提取。用昆蟲(chóng)針取一頭煙粉虱至2 mL離心管中,加入滅菌后的研磨珠,向離心管中加入30 μL配好的DNA提取混合液,將加入提取液的離心管放入組織破碎儀中研磨1 min,8 000 rpm離心30 s,取上清至新的PCR管中并放入PCR儀中進(jìn)行酶促反應(yīng),反應(yīng)程序?yàn)椋旱鞍酌?5 ℃裂解10 min,95 ℃滅活酶5 min。-20 ℃冰箱長(zhǎng)期保存。
1.2.2 mtCOI擴(kuò)增及酶切鑒定煙粉虱生物型 進(jìn)行PCR反應(yīng)擴(kuò)增mtCOI基因片段,反應(yīng)程序參照柳洋[16],引物為:C1-J-2195'-TTGATTTTTTGGTCATCCAGAA
GT-3')和R-BQ-2819(5'-CTGAATA TCGRCGAGGCA
TTCC-3'),擴(kuò)增產(chǎn)物大小為620? bp左右。
參照 Chu 等[17]的方法,應(yīng)用限制性內(nèi)切酶 AseI 酶切已擴(kuò)增到的 620 bp mtCOI 基因片段,通過(guò)酶切產(chǎn)物可判斷煙粉虱的生物型。
用1.5%瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物,用凝膠成像儀觀察并記錄結(jié)果。
1.2.3 mtCOI基因序列測(cè)序及數(shù)據(jù)分析 進(jìn)行PCR反應(yīng),反應(yīng)程序參照 Simon 等[18]的方法,引物為:C1-J-2195(5'-TTGATTTTTTGGTCATCCAGAAGT-3')和L2-N-3014(5'-TCCAA TGCACTAATCTGCCA
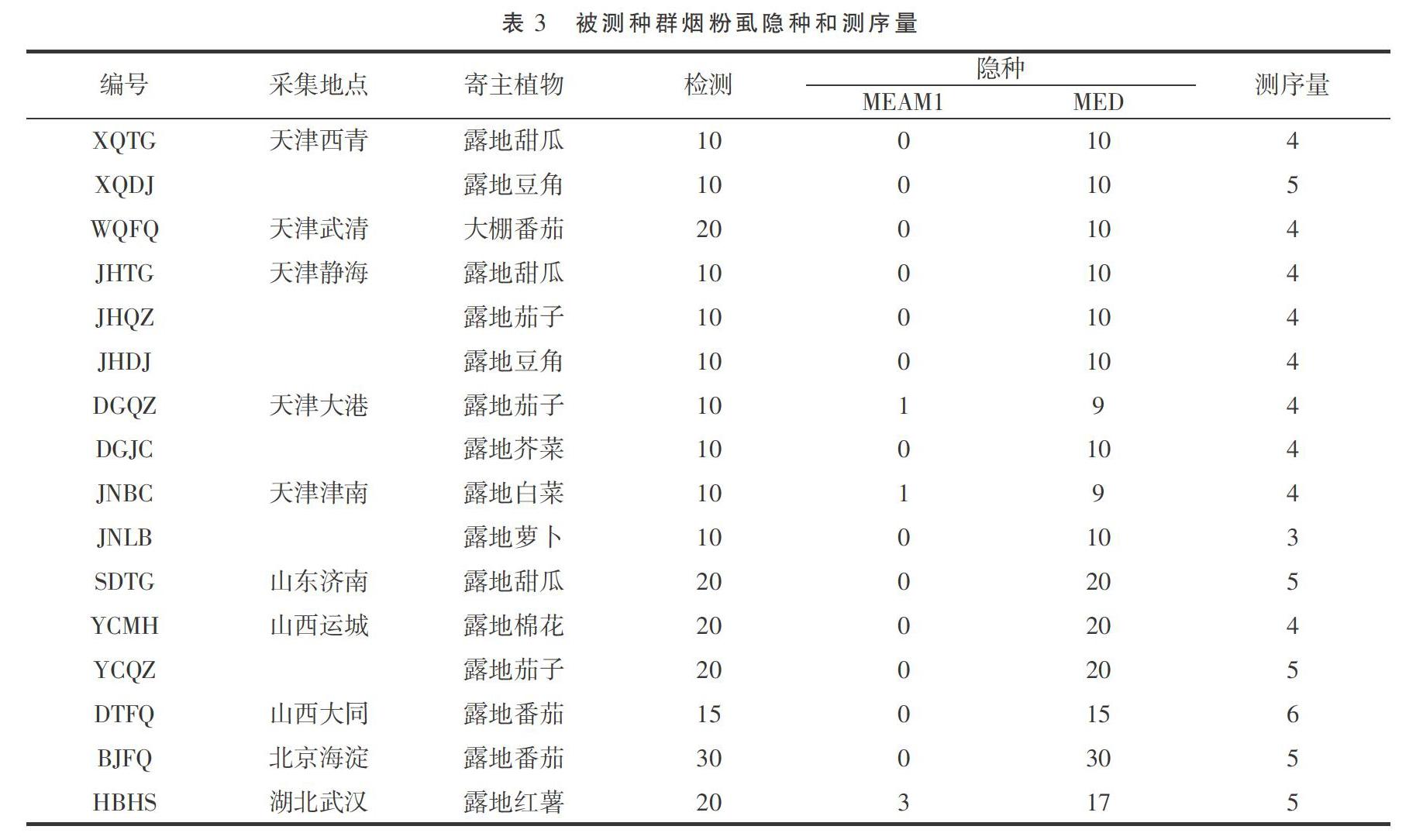
TATTA-3'),擴(kuò)增產(chǎn)物為COI基因3'末端的部分序列,大小為840 bp左右。將PCR產(chǎn)物純化后送往天津擎科生物科技有限公司進(jìn)行雙向測(cè)序,每個(gè)種群至少檢測(cè)3頭煙粉虱。
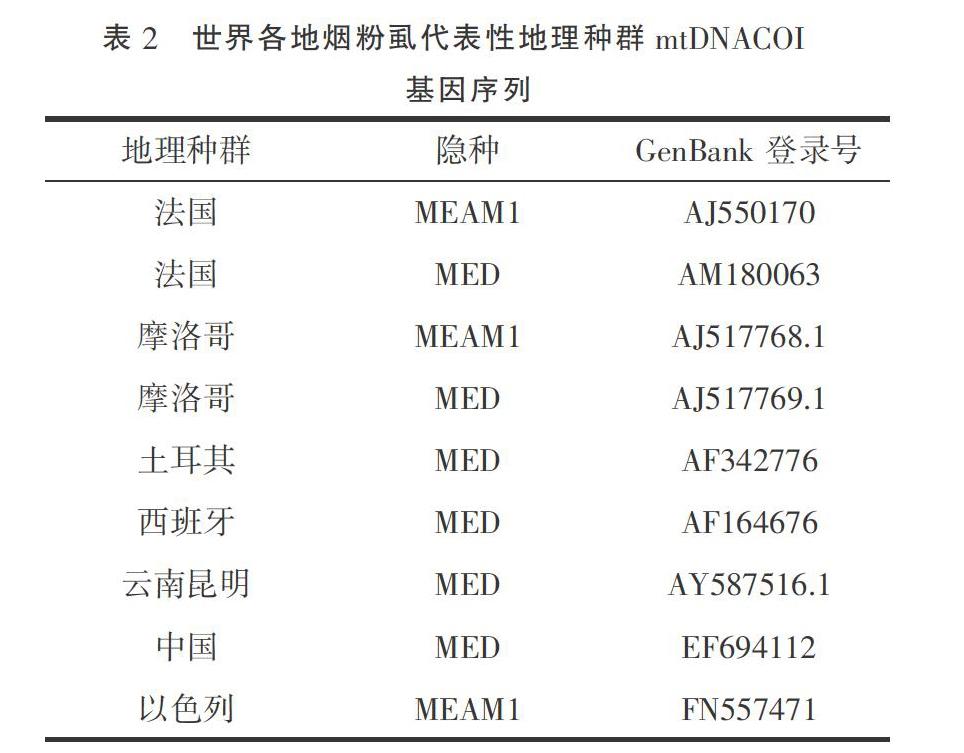
將測(cè)序結(jié)果提交 GenBank,獲取登錄號(hào)(表1),通過(guò)BLAST方法檢索有代表性國(guó)外煙粉虱COI序列(表2)。在16個(gè)種群中,從13個(gè)只有MED煙粉虱的種群各抽選一條測(cè)序結(jié)果;在MEAM1、MED煙粉虱共存的3個(gè)種群中,不同的隱種煙粉虱各抽選一條測(cè)序結(jié)果。以非洲小粉虱[Bemisia afer( Priesner & Hosny)](AF418673.2)為外群[19]。運(yùn)用MEGA 7軟件,根據(jù) Kimura 2- paramter 模型計(jì)算出采樣地區(qū)煙粉虱與國(guó)內(nèi)外不同地理種群及不同生物型煙粉虱的進(jìn)化分歧矩陣,利用鄰位法(neighbor-joining,NJ)和最大似然法(maximum likelihood,ML)分別構(gòu)建系統(tǒng)發(fā)育樹(shù),各分支進(jìn)行1 000次的重復(fù)檢驗(yàn)。
2 結(jié)果與分析
2.1 鑒定不同地區(qū)煙粉虱生物型檢測(cè)結(jié)果
MED煙粉虱的 mtCOI 基因片段存在一個(gè) AseI 酶切位點(diǎn),因而可以被酶切成兩個(gè)片段(498 bp 和122 bp),而 MEAM1 煙粉虱的 mtCOI 基因由于不存在 AseI 酶切位點(diǎn),因此不能被切開(kāi)[10]。
結(jié)果顯示16個(gè)種群中,有13個(gè)種群的煙粉虱都是MED型,有3個(gè)種群是MEAM1、MED混合,且混合種群中以MED為主(表3)。
2.2 根據(jù)mtDNA COI序列構(gòu)建進(jìn)化分歧和系統(tǒng)發(fā)育樹(shù)
2.2.1 構(gòu)建進(jìn)化分歧 非洲小粉虱和其他種群煙粉虱的遺傳距離都在28%以上,MEAM1煙粉虱和MED煙粉虱的遺傳距離基本在4.7%~5.6%之間,我們檢測(cè)地區(qū)的MED煙粉虱的遺傳距離在0%~0.4%之間,與中國(guó)Q型和云南昆明煙粉虱的遺傳距離基本在0.2%之間,低于已知MED煙粉虱的內(nèi)部差異。三個(gè)地區(qū)的MEAM1煙粉虱同源性高達(dá)100%(表4、表5)。
2.2.2基于mtDNA COI基因構(gòu)建系統(tǒng)發(fā)育樹(shù) 使用NJ法構(gòu)建系統(tǒng)樹(shù),各分支均進(jìn)行1 000次的重復(fù)檢驗(yàn)(圖1)。從進(jìn)化樹(shù)中可以看出, MED煙粉虱以99%的置信度聚為一個(gè)分支,且測(cè)得的所有MED煙粉虱與來(lái)自法國(guó)、摩洛哥和西班牙的MED煙粉虱親緣關(guān)系更近,而來(lái)自土耳其的則單獨(dú)聚為一支。所有的MEAM1煙粉虱聚為一支。使用ML法和NJ法構(gòu)建的系統(tǒng)發(fā)育樹(shù)結(jié)果相似(圖2)。
3 討 論
近十年來(lái),我國(guó)煙粉虱的優(yōu)勢(shì)隱種逐漸由MED代替了MEAM1,其傳播的TYLCV對(duì)我國(guó)大部分地區(qū)的番茄造成嚴(yán)重的危害,對(duì)殺蟲(chóng)劑抗藥性的問(wèn)題也日益突出[20]。煙粉虱是一個(gè)復(fù)雜的物種復(fù)合體,不同隱種之間傳播TYLCV和對(duì)殺蟲(chóng)劑抗藥性也都有所差異。明確天津各地區(qū)煙粉虱的隱種現(xiàn)狀,與世界各地不同隱種煙粉虱之間的系統(tǒng)發(fā)育分析,可對(duì)外來(lái)入侵煙粉虱的防治具有重要的指導(dǎo)意義。
本文通過(guò) mtDNA COI基因序列分析鑒定了天津各區(qū)及其附近、山東、山西、湖北等不同地區(qū)煙粉虱的隱種組成和系統(tǒng)生物發(fā)育分析,檢測(cè)的隱種和系統(tǒng)發(fā)育樹(shù)共同表明在監(jiān)測(cè)的各個(gè)地區(qū)的不同寄主植株上的煙粉虱絕大部分是都是MED煙粉虱,只有極少數(shù)的種群存在MEAM1煙粉虱,這與現(xiàn)在MED煙粉虱是主要隱種的現(xiàn)狀是相符的。在2009年,Pan等[7]調(diào)查了天津西青區(qū)的煙粉虱隱種,煙粉虱全部是MEAM1隱種,而今調(diào)查天津各區(qū)煙粉虱隱種幾乎都是MED,與褚棟等[21]的結(jié)論也一致。
許麗麗等[22]推測(cè)中國(guó)MED煙粉虱可能起源于地中海西部地區(qū),我們調(diào)查地區(qū)的MED隱種與來(lái)自法國(guó)、摩洛哥和西班牙的MED煙粉虱親緣關(guān)系更近,土耳其的則單獨(dú)聚為一支,結(jié)果相似,為這一推測(cè)再添實(shí)例,更進(jìn)一步的證明推測(cè)的正確性。
不同隱種對(duì)寄主植株的危害程度是不同的,MED煙粉虱在番茄上的危害程度比MEAM1煙粉虱更加嚴(yán)重[23]。有研究表明,MED煙粉虱比MEAM1煙粉虱對(duì)新類煙堿農(nóng)藥具有更高的抗性[24]。這意味著這些地區(qū)的煙粉虱種群對(duì)新類煙堿農(nóng)藥有較高的抗性,而目前煙粉虱的治理主要通過(guò)噴施農(nóng)藥等化學(xué)途徑,這為防治煙粉虱加大了難度。
本研究中由于采樣地點(diǎn)、數(shù)量、方式等局限性,不排除13個(gè)種群只檢測(cè)出MED煙粉虱的種群是混合種群的可能,因此需要持續(xù)對(duì)天津各地區(qū)煙粉虱進(jìn)行隱種鑒定及系統(tǒng)發(fā)育分析,明確各地區(qū)煙粉虱的詳細(xì)情況,為科學(xué)的防治煙粉虱提供理論依據(jù)。
參考文獻(xiàn):
[1]BROWN J K. Phylogenetic biology of the Bemisia tabaci sibling species group[M]//Bemisia: bionomics and management of a global pest. Springer, Dordrecht, 2009: 31-67.
[2]HOGENHOUT S A, AMMAR E D, WHITFIELD A E, et al. Insect vector interactions with persistently transmitted viruses[J]. Annual review of phytopathology, 2008, 46(1):327-359.
[3]PICD B, DIEZ M J, NUEZ F.Viral diseases causing the greatest economic losses to the tomato crop.II. The Tomato yellow leaf curl virus-a review[J]. Scientia horticulturae,1996,67(3):151-196.
[4]HU J, DE BARRO P, ZHAO H, et al. An extensive field survey combined with a phylogenetic analysis reveals rapid and widespread invasion of two alien whiteflies in China[J]. PloS one,2011,6:el061.
[5]劉銀泉,劉樹(shù)生.煙粉虱的分類地位及在中國(guó)的分布[J].生物安全學(xué)報(bào),2012,21(4):247-255.
[6]CHU D, ZHANG Y J, BROWN J K, et al. Theintroduction oftheexotic Q biotypeofBemisia tabacifromthemediterranean REGION INTO CHINA ON ORNAMENTAL CROPS[J]. Florida entomologist, 2006, 89(2):168-174.
[7]PAN H, CHU D, GE D, et al. Further spread of and domination by Bemisia tabaci(Hemiptera: Aleyrodidae) biotype Q on field crops in China[J]. Journal of economic entomology, 2011, 104(3):978-985.
[8]HOROWITZ A R, DENHOLM I, GORMAN K, et al. Biotype Q of Bemisia tabacii dentified in Israel[J]. Phytoparasitica, 2003, 31(1):94-98.
[9]BROWN J K, CZOSNEK H. Whitefly transmission of plant viruses[J]. Advances in botanical research, 2002, 36(2):65-76.
[10]PAN H P, PREISSER E L, CHU D, et al. Insecticides promote viral outbreaks by altering herbivore competition[J]. Ecological applications, 2016, 25(6):1585-1595.
[11]王少麗, 張友軍, 李如美,等. 北京和湖南煙粉虱生物型及其抗藥性監(jiān)測(cè)[J]. 應(yīng)用昆蟲(chóng)學(xué)報(bào), 2011, 48(1):27-31.
[12]譚永安, 肖留斌, 孫洋,等. 江蘇煙粉虱生物型鑒定與危害程度分析[J]. 環(huán)境昆蟲(chóng)學(xué)報(bào), 2012, 34(3):277-282.
[13]趙瑞華, 鄧振山, 賀曉龍. 陜西地區(qū)煙粉虱生物型的鑒定及其危害分析陜西地區(qū)煙粉虱生物型的鑒定及其危害分析[J]. 延安大學(xué)學(xué)報(bào)(自然科學(xué)版), 2015, 34(4): 82-87.
[14]周新改,馬偉華,劉美剛. 湖北省棉花主產(chǎn)區(qū)煙粉虱生物型分布及系統(tǒng)發(fā)育分析[J]. 植物保護(hù), 2011, 37(5):72-76.
[15]曹騫,李晶,買(mǎi)熱木古麗·克依木,等. 新疆地區(qū)煙粉虱生物型的區(qū)域分布及其攜帶的番茄黃化曲葉病毒檢測(cè)[J]. 昆蟲(chóng)學(xué)報(bào), 2013, 56(6):652-664.
[16]柳洋. 中國(guó)煙粉虱生物型分布、帶毒率及抗藥性監(jiān)測(cè)[D].北京:中國(guó)農(nóng)業(yè)科學(xué)院,2015:6-9.
[17]CHU D, ZHANG Y J, WAN F H. Cryptic invasion of the exotic Bemisia tabaci biotype Q occurred widespread in Shandong Province of China[J]. Florida entomologist, 2010,93(2): 203-207.
[18]SIMON C, FRATI F, BECKENBACH A, et al. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase Chain reaction primers[J]. Annals of the entomological society of America, 1994, 87(6):651-701.
[19]羅晨.煙粉虱的寄主調(diào)查、生物學(xué)特性和分子系統(tǒng)學(xué)研究[D].北京:中國(guó)農(nóng)業(yè)大學(xué),2002.
[20]褚棟, 張友軍.近10年我國(guó)煙粉虱發(fā)生為害及防治研究進(jìn)展[J]. 植物保護(hù), 2018, 44(5):56-60.
[21]褚棟,潘慧鵬,國(guó)棟,等.Q型煙粉虱在中國(guó)的入侵生態(tài)過(guò)程及機(jī)制[J].昆蟲(chóng)學(xué)報(bào),2012,55(12):1399-1405.
[22]許麗麗,蔡力,沈偉江,杜予州.中國(guó)部分地區(qū)煙粉虱生物型種類及系統(tǒng)發(fā)育關(guān)系分析[J].應(yīng)用生態(tài)學(xué)報(bào),2014,25(4):1137-1144.
[23]MUNIZ M, NOMBELA G. Differential variation in development of the B-and Q-Biotypes of Bemisia tabaci (Homoptera: Aleyrodidae) on sweet pepper at constant Temperatures[J]. Environmemology, 2001, 30(4):720-727.
[24]RAUCH N, NAUEN R. Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci(Hemiptera: Aleyrodidae)[J]. Archives of insect biochemistry & physiology, 2010, 54(4):165-176.
