扎龍濕地不同生境蘆葦種群根莖構件的年齡結構
2020-06-03 01:18:54焦德志王昱深楊允菲
生態學報 2020年7期
焦德志,王昱深,楊允菲
1 齊齊哈爾大學生命科學與農林學院,抗性基因工程與寒地生物多樣性保護黑龍江省重點實驗室, 齊齊哈爾 161006 2 東北師范大學草地科學研究所,植被生態科學教育部重點實驗室, 長春 130024
20世紀70年代,英國著名生態學家John L. Harper 在其《Population Biology of Plant》提出植物種群構件理論,把植物種群劃分群落和個體既相關聯又截然不同的層次水平。種群構件生態學的產生和建立,豐富和拓寬了植物種群生態學研究的基本理論、方法和途徑[1]。植物構件是植物對環境適應的表現單位,構件單位間的相互作用形成基株特定的性狀[2]。克隆植物構件的存活時間存在差異性,因此,克隆植物種群的構件均具有不同的齡級組成,種群內不同齡級個體的數量分布稱為種群的年齡結構[3]。年齡結構反映了植物的更新和個體發育狀態,體現了植物種群在復雜生境中的行為策略[4]。研究種群的年齡結構既可以了解現實種群的生長、生產現狀,又能夠預測種群的動態[5]。
木本植物年際變化特性比較明顯,依據年輪的方法來測度其年齡已得到廣泛應用[6-7],采用空間序列代替時間序列,以胸徑-立木徑級代替種群年齡結構也廣泛應用于實驗研究[8- 10]。對于依靠營養繁殖體或營養器官維持多年生活和更新的根莖型禾草,如羊草(Leymuschinensis)[11]、大油芒(Spodiopogonsibiricus)[12]、白草(Pennisetumcentrasiaticum)[13]、野古草(Arundinellahirta)[14]、拂子茅(Calamagrostisepigeios)[15]等植物展開的構件結構研究,發現不同構件均具有穩定的年齡組成,分株齡級均為1—4a間,均呈增長型或穩定型的年齡結構;根莖齡級均為1—5a間,呈以1a根莖為主的增長型或者以2a、3a為主的穩定型年齡結構;潛在種群芽的年齡結構與分株和根莖密切相關,同時受氣候、生境和干擾程度的影響變異較大[5]。
根莖構件是克隆植物養分儲存的資源庫[16],也是克隆植物營養繁殖的基因庫[17]。特殊的性狀特征(包括結構和表型),根莖是從構件尺度研究植物與生境之間的協同演化關系的最佳模式構件之一[18]。蘆葦(Phragmitesaustralis)是典型的長根莖型克隆植物[2,19],在扎龍濕地,蘆葦既可形成大面積單優群落,也可形成群落斑塊[2,20]。扎龍濕地不同生境蘆葦種群分株構件[19]和根莖構件[21]等數量特征,均表現出由遺傳因素控制的比較穩定的季節動態規律[2];同時,個體生長和種群動態又存在明顯的環境效應,生境間的差異及其差異序位均相對穩定,這種穩定差異是土壤因子驅動作用的結果,土壤含水量、有機質、速效氮為正向驅動,pH、速效磷為負向驅動[2,19,21]。扎龍濕地的蘆葦種群主要依靠根莖上的根莖芽進行更新,種群空間擴展主要依靠根莖的生長和延伸,根莖芽庫存量和分株密度與根莖長度顯著或極顯著正相關,根莖芽萌發所需的營養物質由根莖提供[22]。20世紀90年代初扎龍濕地持續干旱缺水,2009年地方政府實施扎龍濕地長效補水機制,生態補水成為維系濕地環境的重要舉措。筆者從2011年在扎龍濕地設置固定樣地開展補水后植被生態監測工作,補水初期(2011年6月)的調查表明,濕生和水生生境的蘆葦根莖由3個齡級組成,旱生和鹽堿生境由4個齡級組成[23]。對扎龍濕地長效補水機制實施多年后,整個生長期(6—10月份)根莖構件年齡組成的調查,了解蘆葦種群根莖構件的存活時間,掌握種群中不同齡級根莖構件的年齡配置及變化規律[2],揭示根莖構件年齡結構調節的生態學意義,預測種群動態與發展,對扎龍濕地生態系統的恢復和保護利用具有重要指導意義[2]。
1 研究地區和研究方法
1.1 研究區概況
本項研究是在扎龍國家級自然保護區 (齊齊哈爾東南23 km) 核心區進行(圖1)。地理坐標46°52′—47°32′N,123°47′—124°37′E,屬于中溫帶大陸性季風氣候[2],年均氣溫為3.2℃,最冷月平均氣溫-19.4℃,最熱月平均氣溫22.9℃,年日照時數2700—3000 h,年輻射總量是2052—2140 J/cm2,積溫2600—3000℃,平均地溫4.9℃,無霜期128 d[24]。
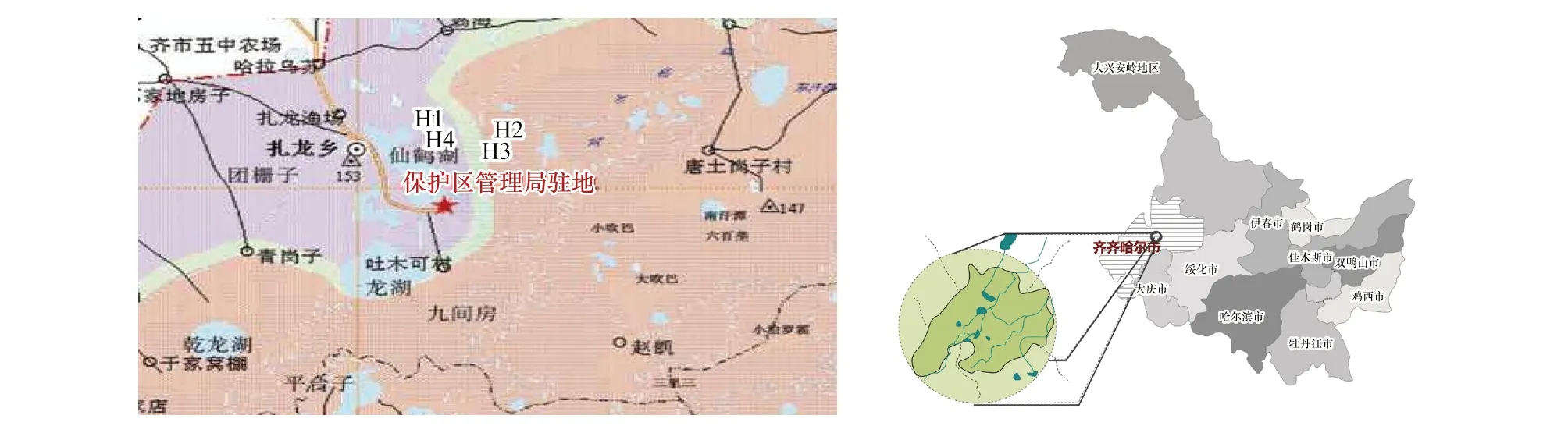
圖1 扎龍濕地位置圖及研究區Fig.1 The location and researched area of Zhalong wetland
1.2 樣地設置
為了更好地闡述和說明科學問題,共設置4個生境樣地(圖2):
1)旱生生境(H1):土壤為草甸土,樣地全年無積水或7—8月份雨季短時積水,蘆葦經常形成羊草+蘆葦混生群落,微地形中心也可形成面積不等的單優種蘆葦群落斑塊,生長旺盛期群落蓋度達到90%以上[19,21]。
2)濕生生境(H2):土壤為沼澤土,樣地平時無積水或少量積水,7—8月雨季來臨以后逐漸積水,積水深度一般情況低于30 cm,蘆葦形成單優種群落,生長旺盛期的群落蓋度可達到95%以上[19,21]。
3)水生生境(H3):土壤為腐殖沼澤土,樣地常年積水,水深變化為10—100 cm,形成濕地內面積最大的蘆葦群落,生長旺盛期群落蓋度95%以上[19,21]。
4)鹽堿生境(H4):土壤為鹽堿土,樣地全年無積水,表層土壤鹽分積累,具有明顯鹽堿斑,有的堿斑可形成單優勢種居群,生長旺盛期群落蓋度在50%以下[19,21]。
樣地多年來一直冬季收割,造紙或民用。另外,4 個生境間距離均在5 km 內,光照和溫度等環境條件可視為一致。4個生境蘆葦群落的土壤因子既存在比較穩定的空間差異,同時,也表現出明顯的季節性波動[21]。

圖2 扎龍濕地4個蘆葦生境的樣地Fig.2 The four experiment plots of Phragmites australis in Zhalong wetland
1.3 研究方法
扎龍濕地蘆葦的返青時間一般在5月10日左右,依據蘆葦的自然生長規律按時間順序取樣(2013年6月15日,7月10日,8月15日,9月12日,10月15日)。在蘆葦分株相對均勻的區域設置樣帶(20 m×20 m),采取單位土體挖掘取樣[21- 24]。按實際生活年限劃分根莖齡級[25],測量不同齡級根莖長度后計算總長度,80℃烘至恒量,稱不同齡級根莖生物量后計算總生物量[2]。
1.4 數據處理
年齡結構以不同齡級根莖長度和生物量所占比率表示。依據根莖生長發育及出現死亡年限判定幼齡、壯齡及老齡[26],以年齡錐體形狀判斷年齡結構類型[27]。旱生生境發現有7a根莖,2個樣方中根莖長度分別為75 cm和92 cm,數據統計時計入6a[28]。以SPSS 19.0軟件進行數據分析,以Excel 2013軟件繪圖。以實際生長時間(5月10日返青計)為變量[28],根莖長度和生物量的比率為因變量進行線性、指數、冪函數回歸,選取相關性最高的擬合方程作為定量刻畫模型,方程的擬合優度以R2檢驗,顯著性采用F檢驗[2]。
2 結果分析
2.1 根莖長度的年齡結構
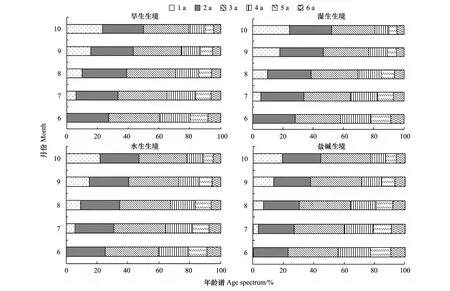
圖3 不同生境蘆葦種群根莖長度年齡結構Fig.3 Age structure on rhizome length of Phragmites australis populations in heterogeneous habitats
扎龍濕地4個生境蘆葦種群根莖構件具有特定的年齡組成,6月份均由2—6a 的5個齡級組成,7—10月份均由1—6a 的6個齡級組成。根莖長度的年齡譜見圖3。由圖可知,1a、2a根莖長度比率各月份均以H4最小,依次是水生生境(H3)、旱生生境(H1),濕生生境(H2)最大;3a、4a比率均以H2最小,依次是H1、H3,鹽堿生境(H4)最大;5a、6a比率均以H1最小,依次是H2、H3,H4最大。6—10月份,1a根莖長度比率為0—24%之間,呈逐漸增加趨勢;2a,3a分別為23%—29%之間和28%—34%之間,除H4略有增加外,其他3個生境均小幅減小;4—6a分別為9%—21%之間,5%—13%之間和4%—9%之間,比率均逐漸減小[2]。共同特征是根莖長度比率均以6a最小,依次是5a,1a,4a,2a,3a最大,根莖齡級越高其長度所占比率越小。
根莖長度比率在月份間的差異上,1a根莖均最大,依次是4a,5a和6a,2a、3a根莖均最小。這種差異表明,1a的幼齡級根莖快速延伸生長,2、3a的壯齡級根莖延伸速度緩慢,4—6a的老齡級根莖逐漸死亡[2]。以此標準劃分根莖年齡階段,在4個生境中,6—9月份蘆葦種群根莖長度均呈衰退型年齡結構,10月份又均轉為穩定型年齡結構。
2.2 根莖長度年齡結構的動態規律
經回歸分析,除2a、3a根莖外,其他4個齡級根莖長度比率與返青后的實際生長時間(5月10日返青計)之間均較好地符合直線函數關系(表1),方程的擬合優度(R2)在0.913—0.998之間,并均達到了P<0.01的極顯著水平。1a根莖長度隨生育期進程按固定比率增加,4—6a根莖則按固定比率減小,不同生境根莖長度的相對比率表現出一致的季節動態規律。
表1 蘆葦種群不同齡級根莖長度比率(y,%)與生長時間(x, d)的擬合方程及顯著性檢驗(n=5)
Table 1 Simulated equations and significance test on relationship between rhizome length rate (y,%) of different age class and growth time (x, d) ofPhragmitesaustralispopulations (n=5)
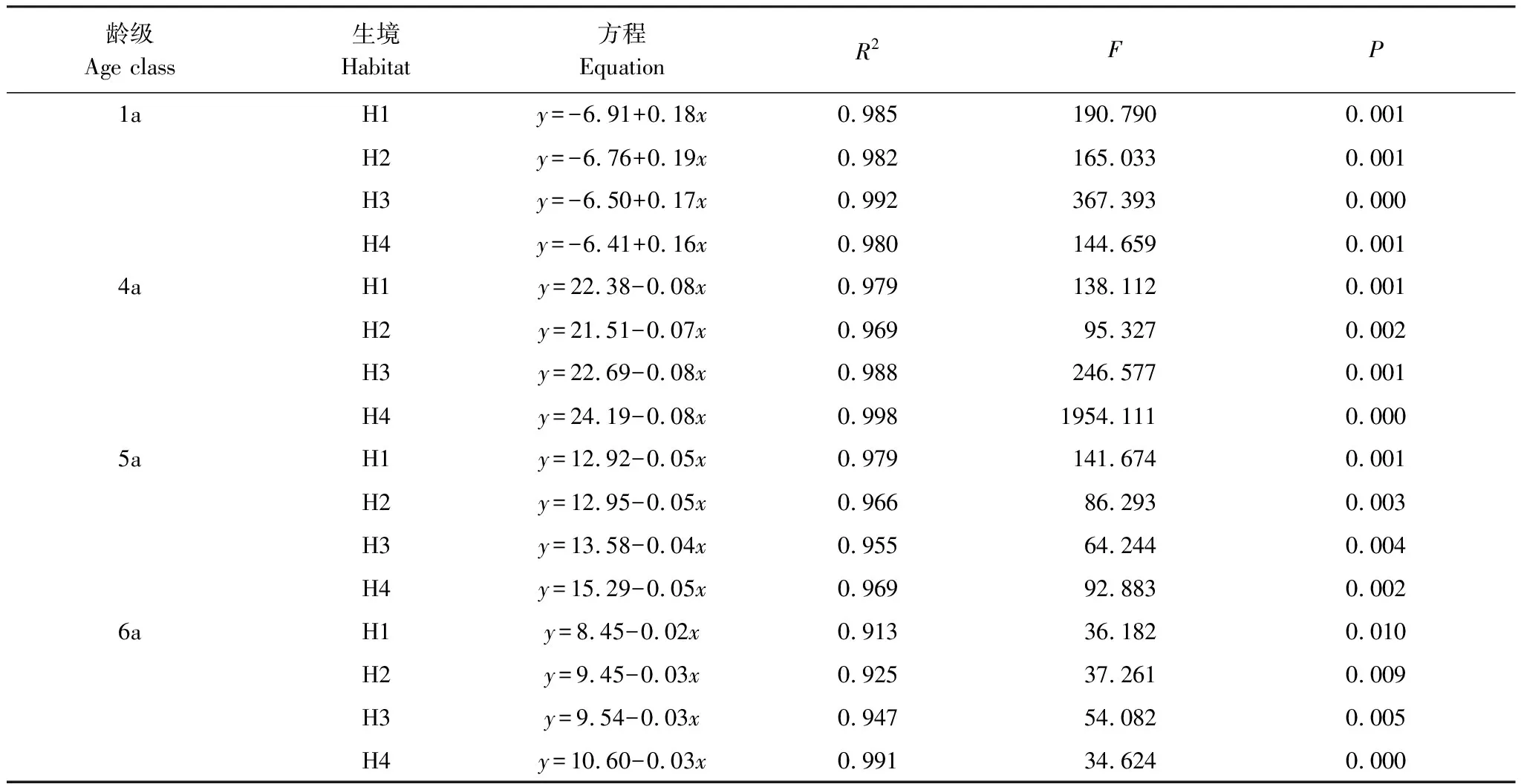
齡級Age class生境Habitat方程EquationR2FP1aH1y=-6.91+0.18x0.985190.7900.001H2y=-6.76+0.19x0.982165.0330.001H3y=-6.50+0.17x0.992367.3930.000H4y=-6.41+0.16x0.980144.6590.0014aH1y=22.38-0.08x0.979138.1120.001H2y=21.51-0.07x0.96995.3270.002H3y=22.69-0.08x0.988246.5770.001H4y=24.19-0.08x0.9981954.1110.0005aH1y=12.92-0.05x0.979141.6740.001H2y=12.95-0.05x0.96686.2930.003H3y=13.58-0.04x0.95564.2440.004H4y=15.29-0.05x0.96992.8830.0026aH1y=8.45-0.02x0.91336.1820.010H2y=9.45-0.03x0.92537.2610.009H3y=9.54-0.03x0.94754.0820.005H4y=10.60-0.03x0.99134.6240.000
H1:旱生生境Xeric habitats;H2:濕生生境Wet habitats; H3:水生生境Aquatic habitats;H4:鹽堿生境Saline-alkali habitats
2.3 根莖生物量的年齡結構
4個生境蘆葦種群根莖生物量的年齡譜見圖4。由圖4可知,1a、2a根莖生物量比率各月份均以鹽堿生境(H4)最小,依次是水生生境(H3)、旱生生境(H1),濕生生境(H2)最大;3a、4a比率均以H1最小,依次是H2和H3,H4最大;5a、6a比率均以H3最小,依次是H2和H1,H4最大。6—10月份,1—3a根莖生物量的比率逐漸增加,1a為0—16%之間,2a為12%—26%之間,3a為26%—38%之間;4—6a根莖生物量的比率逐漸減小,4a為13%—27%之間,5a為7%—16%之間,6a為4%—13%之間。共同特征均以1a根莖生物量比率最小,依次為6a、5a、2a、4a,以3a比率最大。
根莖生物量比率在月份間的差異上,1a根莖均最大,依次是6a,5a,4a和2a,3a根莖均最小。1a根莖為幼齡級,2a、3a根莖為壯齡級,4—6a根莖老齡級。如果以此劃分根莖年齡階段,6—10月份,4個生境蘆葦種群根莖生物量均呈衰退型年齡結構[2]。
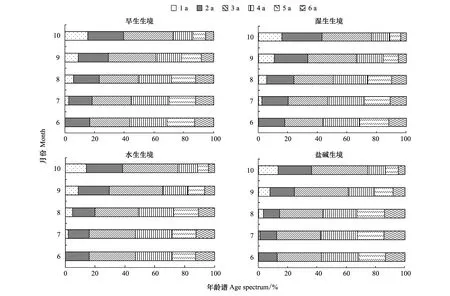
圖4 不同生境蘆葦種群根莖生物量年齡結構Fig.4 Age structure on rhizome biomass of Phragmites australis populations in heterogeneous habitats
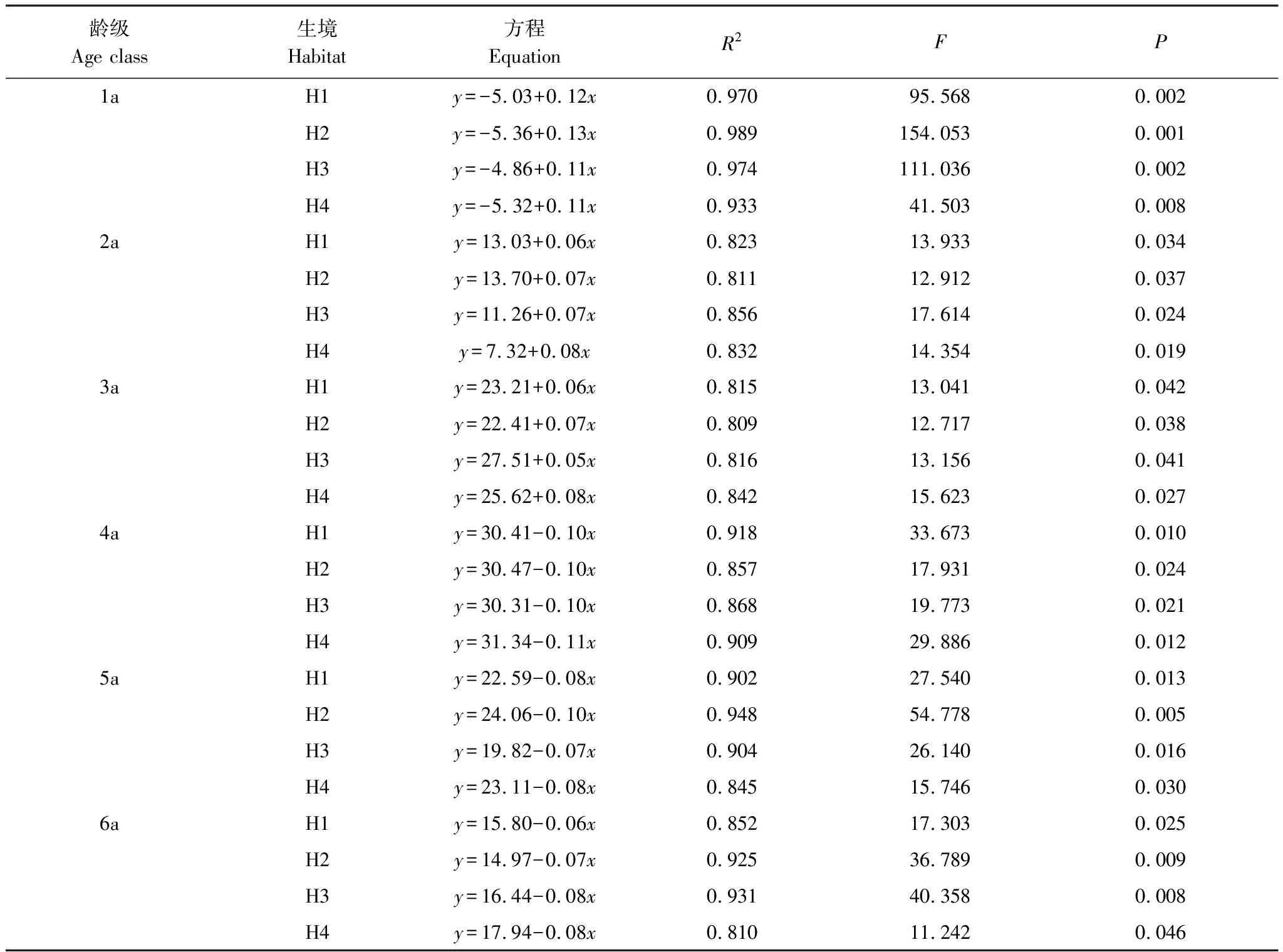
2.4 根莖生物量年齡結構的動態規律
進一步回歸分析,4個生境不同齡級根莖生物量比率與返青后的實際生長時間(5月10日返青計)之間均較好地符合直線函數關系[2](表2),方程的擬合優度(R2)在0.810—0.989之間,并均達到了P<0.05的顯著水平[2]。幼齡級(1a)和壯齡級(2a、3a)根莖生物量按固定比率升高,老齡級根莖(4—6a)則降低,不同生境根莖生物量的相對比率具有一致的季節動態規律[2]。
3 討論
3.1 構件年齡結構穩定和調節的生物學意義
種群的年齡結構對種群的穩定性和種群的數量動態有很好的詮釋作用,也可以釋譯種群與環境的相互關系及其在群落中的地位和作用[29]。克隆生長是克隆植物實現生存空間拓展的主要手段,形成現實種群鞏固已占據的空間是其首先表現,而保持根莖具有較高存活力對潛在空間的拓展意義更重大[22]。開展根莖構件年齡結構研究,不僅可以獲得其數量和發展動態等種群統計信息,而獲得其生產和調節等功能信息的生物學意義更有價值[2,22]。
對扎龍濕地蘆葦種群根莖長度和生物量的研究表明,其生境間數量特征的穩定差異是土壤因子驅動作用的結果,土壤含水量和pH的驅動力最大,含水量為正向驅動,pH為負向驅動[21]。但種群在新根莖的形成、老根莖的衰老和死亡以及養分貯藏上表現出相對穩定的生物學特性。整個生長季內,幼齡級根莖(1a)保持較快的延伸生長,是其擴展水平生態位的重要手段,同時對根莖種群進行不斷補充更新[2,30],保持種群具有長久生活力;壯齡級根莖(2a,3a)在種群中所占比率(51%—63%)較大且相對穩定,借以維持著種群的相對穩定;根莖從4a開始出現死亡現象,齡級增高所占比率隨之降低直至消失。種群中幼齡級根莖長度的比率高于(10%—14%)老齡級,根莖生物量的比率正相反,這種分配比率保證老齡級根莖仍具有較高的物質貯藏水平,確保種群較高的繁殖能力[31]。因此,蘆葦種群根莖構件具有特定的年齡組成,也形成穩定的年齡結構,不同齡級根莖構件在種群的更新和擴展中的功能和作用不同。
表2 蘆葦種群不同齡級根莖生物量比率(y,%)與生長時間(x, d)的擬合方程及顯著性檢驗(n=5)[2]
Table 2 Simulated equations and significance test on relationship between rhizome biomass rate(y,%)of different age class and growth time(x, d)ofPhragmitesaustralispopulations(n=5)[2]
齡級Age class生境Habitat方程EquationR2FP1aH1y=-5.03+0.12x0.97095.5680.002H2y=-5.36+0.13x0.989154.0530.001H3y=-4.86+0.11x0.974111.0360.002H4y=-5.32+0.11x0.93341.5030.0082aH1y=13.03+0.06x0.82313.9330.034H2y=13.70+0.07x0.81112.9120.037H3y=11.26+0.07x0.85617.6140.024H4y=7.32+0.08x0.83214.3540.0193aH1y=23.21+0.06x0.81513.0410.042H2y=22.41+0.07x0.80912.7170.038H3y=27.51+0.05x0.81613.1560.041H4y=25.62+0.08x0.84215.6230.0274aH1y=30.41-0.10x0.91833.6730.010H2y=30.47-0.10x0.85717.9310.024H3y=30.31-0.10x0.86819.7730.021H4y=31.34-0.11x0.90929.8860.0125aH1y=22.59-0.08x0.90227.5400.013H2y=24.06-0.10x0.94854.7780.005H3y=19.82-0.07x0.90426.1400.016H4y=23.11-0.08x0.84515.7460.0306aH1y=15.80-0.06x0.85217.3030.025H2y=14.97-0.07x0.92536.7890.009H3y=16.44-0.08x0.93140.3580.008H4y=17.94-0.08x0.81011.2420.046
H1:旱生生境Xeric habitats;H2:濕生生境Wet habitats; H3:水生生境Aquatic habitats;H4:鹽堿生境Saline-alkali habitats
克隆植物構件的累計數量與生境條件和種間競爭有關,但相對數量指標年齡譜則比較接近,前者反映了生境間差異引起的生態學特性,后者反映了物種遺傳因素決定的生物學特性。不同生境斑苦竹(Pleioblastusmaculatus)可以自我調節實現種群的穩定,種群的年齡結構從波動的增長型轉為穩定型[2,32]。相對潮濕、疏松的土壤環境中假葦拂子茅(Calamagrostispseudophragmites)根莖較長,但均以1a或2a根莖比率最高,隨齡級升越其比率呈降低趨勢[33]。恢復4—5年賴草(LeymusSecalinus)種群的根莖由3個齡級組成,而恢復11—12年根莖由4個齡級組成[34],但不同植被恢復期內,賴草種群根莖的年齡結構均為穩定型,隨著恢復時間的延長,穩定性也明顯增強[34]。松嫩平原的山黧豆(Lathyrusquinquenervius),生長季后期根莖由3個齡級組成,其中,長度以1a和2a為主,3a大幅減少,種群對新生根莖的物質投資有所增加,通過指數函數方式增加子代數量,占據有限空間[2,35]。
3.2 構件年齡結構對種群動態預測的可靠性
種群的年齡結構可以反映植物的更新和個體發育狀態,而且體現植物種群在復雜生境中所表現出的行為策略[4]。種群的年齡結構不僅可以客觀地反映出種群內各齡級的生活力、生產力以及現存狀態,還對分析種群動態與預測種群未來發展趨勢有重要價值[36]。
4個生境蘆葦種群根莖長度和生物量的年齡結構存在季節動態。根莖長度由衰退型結構轉為穩定型結構[2],在下一個生長季根莖長度至少維持在原有水平;根莖生物量在整個生長季均為衰退型年齡結構,這種年齡結構能夠保持老齡級根莖具有更強的繁殖力,也是蘆葦種群對根莖構件的物質分配策略。根莖和根莖芽為蘆葦潛在種群,對種群統計而言,分株才具有更高的實際意義和價值。因此,現實種群對種群動態預測的真實性更高,潛在種群對種群預測具有局限性,一般在上一個生長季末期或下一個生長季初期會更真實[15,37]。
2011年6月對扎龍濕地蘆葦種群根莖的調查結果表明:旱生和鹽堿生境根莖由2—5a的4個齡級組成,濕生和水生生境由2—4a 的3個齡級組成[23,28]。2013年4個生境蘆葦種群根莖均由1—6a的6個齡級組成,在旱生生境個別樣方有7a根莖存在。2009年以來,地方政府對扎龍濕地實施長效補水,有效地改善了濕地嚴重缺水的現象,濕地土壤含水量的正向驅動作用增強,pH的負向驅動作用減弱,促進了濕地的正向演替進程,使濕地逐漸恢復穩定和健康。因此,研究結果為扎龍濕地的科學管理實踐提供重要理論支持。
