阿根廷巴塔哥尼亞陸架拉氏南美南極魚(yú)(Patagonotothen ramsayi)不同組織中脂肪酸分布及食物來(lái)源指示
2020-05-23 08:42:10楊清源朱國(guó)平
海洋學(xué)報(bào) 2020年4期
楊清源,朱國(guó)平,4*
(1. 上海海洋大學(xué) 海洋科學(xué)學(xué)院,上海 201306;2. 上海海洋大學(xué) 極地研究中心,上海 201306;3. 大洋漁業(yè)資源可持續(xù)開(kāi)發(fā)教育部重點(diǎn)實(shí)驗(yàn)室 極地海洋生態(tài)系統(tǒng)研究室,上海 201306;4. 國(guó)家遠(yuǎn)洋漁業(yè)工程技術(shù)研究中心,上海 201306)
1 引言
南美洲南部大陸架水域共有14種南美南極魚(yú)[1],其中位于巴塔哥尼亞海域的拉氏南美南極魚(yú)(Patagonotothen ramsayi)數(shù)量最為豐富[2]。盡管拉氏南美南極魚(yú)具有重要的生物學(xué)意義,但對(duì)于其生物學(xué)的研究仍不夠深入。Ekau[2]發(fā)現(xiàn)這一物種明顯表現(xiàn)出性別的二態(tài)性,雄性的臀鰭、腹鰭和身體前部呈深黑色,而雌性則呈棕色至綠色。該文還指出,拉氏南美南極魚(yú)產(chǎn)卵發(fā)生在南半球秋季的阿根廷陸架以及春季的布爾德伍德淺灘。Sosiński和Janusz[3]也證實(shí)了兩個(gè)區(qū)域產(chǎn)卵時(shí)間的差異。Brickle等[4]對(duì)拉氏南美南極魚(yú)的年齡與生長(zhǎng)進(jìn)行了研究,發(fā)現(xiàn)它是一種生長(zhǎng)速度相對(duì)較慢的魚(yú)類,最大年齡可達(dá)14齡。另外,Hart[5]還提供了一些關(guān)于其攝食及捕食者的信息。因拉氏南美南極魚(yú)存在晝夜垂直移動(dòng)的特性[6],所以它不僅可以捕食一些膠狀及甲殼類的浮游生物,還會(huì)捕食部分底棲生物[2,7]。近期研究還發(fā)現(xiàn),拉氏南美南極魚(yú)偶爾也會(huì)捕食一些漁業(yè)丟棄物,尤其是魷魚(yú)[8-9]。與此同時(shí),拉氏南美南極魚(yú)也是許多大型魚(yú)類的攝食對(duì)象[7],甚至被巴布亞企鵝(Pygoscelis papua)所捕食[10]。該過(guò)程對(duì)于巴塔哥尼亞海域能量由海表至海底間的傳遞有著重要作用。因此,開(kāi)展拉氏南美南極魚(yú)攝食生態(tài)學(xué)的研究可為了解由低營(yíng)養(yǎng)級(jí)至高營(yíng)養(yǎng)級(jí)之間的能量流動(dòng)以及有機(jī)質(zhì)由上層至中下層的垂直運(yùn)輸過(guò)程提供依據(jù)。
使用傳統(tǒng)的胃含物分析法可以知曉拉氏南美南極魚(yú)的主要攝食對(duì)象、攝食的時(shí)空差異[6-7,9,11]。但該方法只能反映短期的攝食情況,對(duì)于食物同化情況以及長(zhǎng)期的攝食趨勢(shì)不能夠反映。為了解決這個(gè)問(wèn)題,已有學(xué)者提出利用肌肉組織中穩(wěn)定同位素及脂類(尤其是脂肪酸和脂肪醇)作為食性指標(biāo)[12-15]。對(duì)于脂類來(lái)說(shuō),食物中的脂肪酸組成會(huì)影響肝臟、性腺、肌肉等組織的脂質(zhì)成分[16]。dos Santos等[17]曾分析不同攝食條件下鱈魚(yú)不同組織內(nèi)脂肪酸組成情況,Beckmann等[18]則分析了不同喂養(yǎng)條件下,鯊魚(yú)肝臟和肌肉組織中脂肪酸分布情況。因此,若利用脂肪酸作為生物標(biāo)志物探究拉氏南美南極魚(yú)的攝食生態(tài)學(xué),需要了解各組織中脂肪酸的分布情況。目前,關(guān)于拉氏南美南極魚(yú)脂肪酸組成的研究?jī)H見(jiàn)于肌肉組織,且僅對(duì)其營(yíng)養(yǎng)價(jià)值進(jìn)行了評(píng)估[19],尚未有人將其脂肪酸組成應(yīng)用于攝食生態(tài)學(xué)研究。鑒于此,本研究旨在:(1)描述拉氏南美南極魚(yú)3種組織中脂肪酸分布情況,(2)分析3種組織中的脂肪酸是否能表征其攝食情況,以及(3)研究拉氏南美南極魚(yú)的食性及其能量轉(zhuǎn)移情況。
2 方法
2.1 樣品收集
拉氏南美南極魚(yú)樣本由中國(guó)水產(chǎn)有限公司“龍騰”號(hào)于西南大西洋水域進(jìn)行中層拖網(wǎng)作業(yè)時(shí)所收集,采樣深度范圍為20~160 m(即阿根廷巴塔哥尼亞大陸架),取樣時(shí)間為2014年12月至2015年4月,取樣站點(diǎn)如圖1所示。所有海上隨機(jī)采集的樣本立即放入-20℃冷凍條件下保存,運(yùn)回國(guó)內(nèi)實(shí)驗(yàn)室后放入-80℃的超低溫冰箱內(nèi)保存,以便進(jìn)行脂類分析。
2.2 方法
2.2.1 脂肪酸提取及測(cè)定
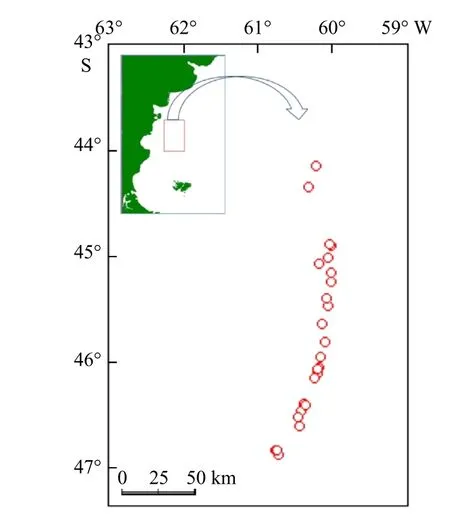
圖1 調(diào)查區(qū)域與站點(diǎn)Fig. 1 The study area with showing the sample locations
在實(shí)驗(yàn)室中,將樣本解凍后首先測(cè)定其體長(zhǎng)和體質(zhì)量等,其中體長(zhǎng)測(cè)量使用精度為1 mm的鋼尺,體質(zhì)量測(cè)定使用精度為1 g的電子秤;測(cè)量完基礎(chǔ)生物學(xué)后,取樣本背鰭附近肌肉、肝臟及性腺部分,用超純水沖洗干凈后裝入離心管,然后放入冷凍干燥機(jī)(Christ Alpha 1-4),在-55°C 下冷凍干燥 24 h,所有樣品使用混合型球磨儀(Retsch - MM440)粉碎,裝入離心管待測(cè)。根據(jù)Folch法進(jìn)行粗酯的提取[20]。依據(jù)GB/T 17376—2008《動(dòng)植物油脂脂肪酸甲酯制備》三氟化硼法進(jìn)行脂肪酸甲酯化。以37種脂肪酸混標(biāo)及內(nèi)標(biāo)十九烷酸甲酯標(biāo)品作為標(biāo)準(zhǔn),利用氣相色譜-質(zhì)譜聯(lián)用儀(Agilent,GC-MS 7890B)進(jìn)行測(cè)試。
2.2.2 指標(biāo)的測(cè)定與計(jì)算
以37種脂肪酸混標(biāo)及內(nèi)標(biāo)十九烷酸甲酯標(biāo)品作為標(biāo)準(zhǔn),通過(guò)比對(duì)保留時(shí)間對(duì)脂肪酸進(jìn)行定性分析,采用內(nèi)標(biāo)法對(duì)脂肪酸進(jìn)行定量分析。脂肪酸含量使用以下內(nèi)標(biāo)法公式得出其質(zhì)量分?jǐn)?shù),并用單位干質(zhì)量的拉氏南美南極魚(yú)肌肉組織中每種脂肪酸的含量(單位:mg/g)表示,即
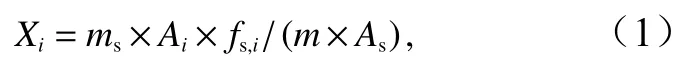
式中,Xi為待測(cè)樣品中組分i的含量,單位為mg/g;ms為待測(cè)樣品中加入內(nèi)標(biāo)物的質(zhì)量,單位為mg;Ai為組分i的峰面積,無(wú)單位;fs,i為組分i與內(nèi)標(biāo)物的校正因子之比,稱為相對(duì)校正因子,無(wú)單位;m為樣品的質(zhì)量,單位為g;As為待測(cè)樣品中內(nèi)標(biāo)物的峰面積,無(wú)單位。
2.2.3 指示食性的特征脂肪酸
特征脂肪酸可以指示生物食性,結(jié)合已有研究(表1),分析不同組織中脂肪酸組成與食性關(guān)系。
2.3 數(shù)據(jù)處理
首先利用Shapiro-Wilk檢驗(yàn)分析脂肪酸含量數(shù)據(jù)是否符合正態(tài)分布,若不符合,則將數(shù)據(jù)轉(zhuǎn)換成對(duì)數(shù)形式后再進(jìn)行檢驗(yàn),以使得數(shù)據(jù)符合正態(tài)分布。利用配對(duì)t檢驗(yàn)分析兩兩脂肪酸含量或類型之間是否存在顯著性差異(顯著性水平設(shè)為0.05)。使用主成分分析(Principal Components Analysis,PCA)以研究 3 種組織間脂肪酸特征的變化,并鑒定對(duì)這種變異影響最大的脂肪酸類型。所有平均值均以平均值±標(biāo)準(zhǔn)差表示。PCA及統(tǒng)計(jì)分析均采用R程序(R Core Team 2017)處理。

表1 在拉氏南美南極魚(yú)體內(nèi)發(fā)現(xiàn)的主要脂肪酸及直接與間接食物源Table 1 The main fatty acids found in the rock cod and their corresponding potential food sources
3 結(jié)果
3.1 拉氏南美南極魚(yú)生物學(xué)信息
拉氏南美南極魚(yú)體長(zhǎng)范圍為111.0~288.0 mm,平均體長(zhǎng)為(209.7±40.9)mm;體質(zhì)量范圍為 20.8~451.3 g,平均體質(zhì)量為(177.12±115.90)g。據(jù) Sosiński和Janusz[3]的研究,拉氏南美南極魚(yú)于體長(zhǎng)為240 mm時(shí)達(dá)到性成熟。據(jù)此,本研究將樣品體長(zhǎng)分為2組:100 mm≤L<240 mm屬小體長(zhǎng)組,240 mm≤L歸大體長(zhǎng)組(表2)。

表2 拉氏南美南極魚(yú)體長(zhǎng)分組Table 2 The size group of the rock cod
3.2 3種組織中脂肪酸組成
由西南大西洋拉氏南美南極魚(yú)3種組織中共檢測(cè)出27種脂肪酸,飽和脂肪酸(Saturated Fatty Acid,SFA)包括10種,單不飽和脂肪酸(Monounsaturated Fatty Acid, MUFA)和多不飽和脂肪酸(Polyunsaturated Fatty Acid, PUFA)分別含7種和10種(表3)。對(duì)于3個(gè)組織而言,肌肉組織中脂肪酸含量最低為(95.39±29.14)mg/g,顯著低于肝臟((423.89±287.16)mg/g)和性腺((372.48±302.71)mg/g)組織(配對(duì)t檢驗(yàn):肌肉組織與肝臟組織,t=-6.99,p<0.001;肌肉組織與性腺組織,t=-5.62,p<0.01)。對(duì)于 SFA,肝臟組織與性腺組織中含量均較高,顯著高于肌肉組織(肝臟組織與肌肉組織,t=-8.14,p<0.001;性腺組織與肌肉組織,t=-5.77,p<0.001)。對(duì)于 MUFA,肝臟組織中含量最高,顯著高于性腺組織和肌肉組織(肝臟組織與性腺組織,t=3.54,p<0.001;肝臟組織與肌肉組織,t=6.63,p<0.001)。就PUFA而言,性腺組織中含量最高,顯著高于肝臟和肌肉組織(性腺組織與肝臟組織,t=2.56,p<0.05;性腺組織與肌肉組織,t=4.99,p<0.001)。對(duì)于n-3 MUFA,性腺組織中含量最高,肝臟組織次之,且均顯著高于肌肉組織(性腺組織與肌肉組織,t=-3.24,p=0.001<0.05;肝臟組織與肌肉組織,t=-2.47,p=0.009<0.05);而在 n-6 PUFA 中,同樣的性腺組織含量最高,肝臟組織次之,且性腺組織含量顯著高于肝臟組織;另外,兩者含量均顯著高于肌肉組織(性腺組織與肝臟組織,t=-3.66,p<0.001;性腺組織與肌肉組織,t=-4.78,p<0.001;肝臟組織與肌肉組織,t=-6.35,p<0.001)。
在C20:5n3與C22:6n3中,性腺組織中含量最高,肝臟組織次之,均顯著高于肌肉組織(C20:5n3∶性腺組織與肌肉組織,t=-3.07,p=0.002<0.05;肝臟組織與肌肉組織,t=-3.00,p=0.003<0.05;C22:6n3:性腺組織與肌肉組織,t=-2.03,p=0.025<0.05;肝臟組織與肌肉組織,t=-1.84,p=0.037<0.05)。對(duì)于 C20:4n6 來(lái)說(shuō),性腺組織中含量最高,肝臟組織次之,且性腺組織顯著高于肝臟組織,兩者均顯著高于肌肉組織(性腺組織與肌肉組織,t=-4.96,p<0.001;肝臟組織與肌肉組織,t=-6.10,p<0.001;性腺組織與肝臟組織,t=-3.87,p<0.001)。
3.3 脂肪酸組成主成分分析
對(duì)拉氏南美南極魚(yú)3個(gè)組織脂肪酸組成進(jìn)行主成分分析后發(fā)現(xiàn),主成分1和主成分2分別解釋了3個(gè)組織脂肪酸組成總變異的71.61%和11.25%(圖2)。由因子載荷圖可以看出,主成分1維度負(fù)向上有C15:0+C17:0、C20:1+C22:1、C18:2n6以及 C20:4n6;主成分2維度正向上有C14:0、C16:0、C16:1n7和C18:1n9;負(fù)向上有C20:5n3及C22:6n3。
由因子得分圖可知,3個(gè)組織之間表現(xiàn)出明顯的差異。肝臟組織主要分布在主成分1正向及主成分2的正向上,該組織中C14:0、C16:0、C16:1n7和C18:1n9這4種脂肪酸對(duì)其影響較大;而性腺組織主要分布于主成分1的負(fù)向上,C15:0+C17:0、C18:2n6、C20:4n6及C20:1+C22:1對(duì)其脂肪酸組成的貢獻(xiàn)較大;肌肉組織主要分布于主成分1的負(fù)向和主成分2的負(fù)向上,除與性腺組織的脂肪酸具有同源脂肪酸外,C20:5n3和C22:6n3對(duì)其的貢獻(xiàn)也較大。
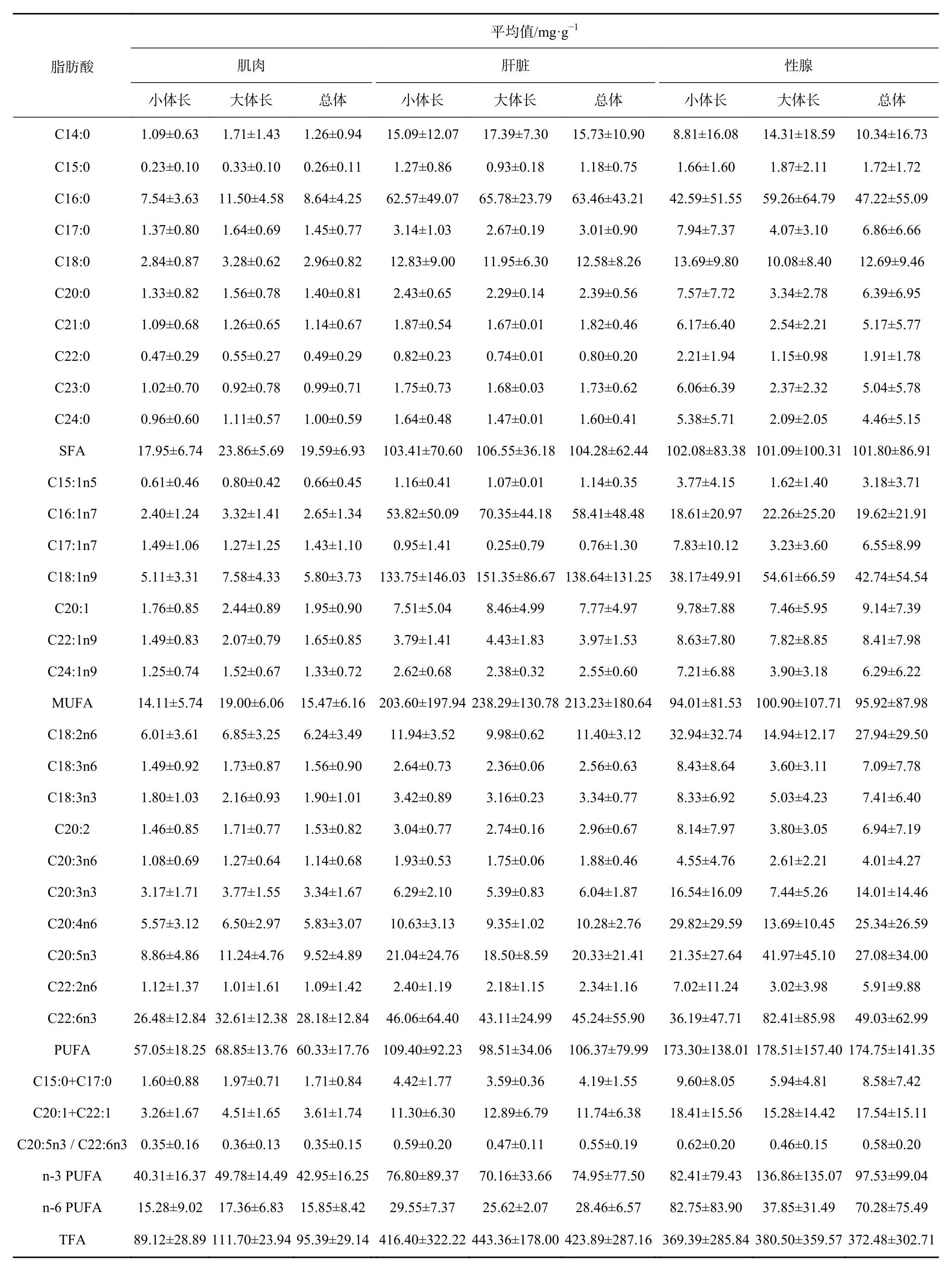
表3 拉氏南美南極魚(yú)不同組織中脂肪酸含量Table 3 Fatty acid contents in different tissues of the rock cod

圖2 拉氏南美南極魚(yú)不同組織脂肪酸含量主成分分析Fig. 2 Principal component analysis plot in different tissues of the rock cod
3.4 體長(zhǎng)與特征脂肪酸含量的關(guān)系
表4顯示了拉氏南美南極魚(yú)3種組織脂肪酸含量與體長(zhǎng)之間的相關(guān)性。由表4可見(jiàn),肌肉組織的特征脂肪酸中,有3種脂肪酸含量與體長(zhǎng)之間存在著明顯的線性關(guān)系,分別為C15:0+C17:0、C16:0及C20:1+C22:1,且隨著體長(zhǎng)的增加而增加;就肝臟組織而言,有3種特征脂肪酸含量與體長(zhǎng)之間存在著顯著的線性關(guān)系,分別為 C15:0+C17:0、C18:2n6、C20:4n6,隨著體長(zhǎng)的增加而降低。性腺組織中,有4種特征脂肪酸與體長(zhǎng)之間存在顯著的線性關(guān)系,分別為C15:0+C17:0、C18:2n6、C20:1+C22:1以及 C20:4n6,同樣隨著體長(zhǎng)的增加而降低。
4 討論
4.1 3種組織對(duì)脂肪酸的吸收與分配
魚(yú)類在其不同生活史階段可能會(huì)轉(zhuǎn)變食性,且隨著魚(yú)類的生長(zhǎng),身體各組織部位對(duì)脂肪酸的吸收以及利用均有所不同。在魚(yú)類未達(dá)性成熟之前,魚(yú)類由外界食物來(lái)源獲取的能量(脂肪酸)用于其身體及各組織的生長(zhǎng);而當(dāng)魚(yú)類達(dá)到性成熟之后,性腺部位獲取的脂肪酸在繁殖階段則更多地用于繁殖。基于Ekau[2]的研究,本研究中的拉氏南美南極魚(yú)樣本均未達(dá)到性成熟。因此,本文中僅由3種組織的能量分配用于生長(zhǎng)這個(gè)角度進(jìn)行分析。

表4 拉氏南美南極魚(yú)體長(zhǎng)與特征脂肪酸含量之間的關(guān)系Table 4 The relationship between standard length and percentage of specific fatty acids in the different tissues of rock cod
脂肪酸是魚(yú)體的重要組成成分。有研究表明,肝臟是脂肪合成與轉(zhuǎn)化的主要場(chǎng)所,肝臟中有豐富的脂肪積累,可為性腺、肌肉等組織的發(fā)育提供能量[27-29]。拉氏南美南極魚(yú)3種組織中,肝臟中總脂肪酸含量為(423.89±287.16)mg/g,遠(yuǎn)高于性腺中的(372.48±302.71)mg/g 及肌肉的(95.39±29.14)mg/g。由此可見(jiàn),拉氏南美南極魚(yú)更傾向于在肝臟中存儲(chǔ)及合成脂類,之后通過(guò)血液、淋巴系統(tǒng)等將脂肪酸轉(zhuǎn)移到其他組織[21]。此外,肝臟中SFA和MUFA含量均為3個(gè)組織中最高,而PUFA含量卻相對(duì)較低,這可能由于食物中的PUFA被優(yōu)先輸送到性腺和肌肉等組織中,以供應(yīng)生長(zhǎng)和發(fā)育的需求。類似的結(jié)果在貝類中也得到證實(shí)[22],這種差異的存在也因各組織具有的不同生理功能所致。性腺的PUFA含量顯著高于肝臟和肌肉,這是因?yàn)镻UFA在卵巢中選擇性地積累規(guī)律所致,豐富的PUFA可以為卵子和胚胎的發(fā)育提供相應(yīng)的營(yíng)養(yǎng),如形成細(xì)胞膜結(jié)構(gòu)、調(diào)節(jié)細(xì)胞膜的離子通道和促進(jìn)發(fā)育等[30];其中n-3 PUFA和n-6 PUFA最為重要,特別是 C22:6n3(DHA)、C20:5n3(EPA)和C20:4n6(ARA)。DHA和EPA是海水魚(yú)類不能完全依靠自身合成以滿足其生長(zhǎng)發(fā)育所需的長(zhǎng)鏈不飽和脂肪酸,必須由食物中獲取,ARA在某些海水魚(yú)類的正常發(fā)育中也起著重要作用[31]。由本文結(jié)果來(lái)看,拉氏南美南極魚(yú)3個(gè)組織中DHA、EPA和ARA的含量均以性腺最高,肝臟次之,肌肉最低。這表明拉氏南美南極魚(yú)性腺組織在生長(zhǎng)過(guò)程中選擇性地儲(chǔ)存這些脂肪酸。由于本研究所使用的樣本體長(zhǎng)多低于27.5 cm,未達(dá)到性成熟體長(zhǎng)[2,32]。所以,可推測(cè)現(xiàn)階段對(duì)于脂肪酸的積累為后期性腺發(fā)育儲(chǔ)備了條件,如ARA可以提升精子的生存能力[33],DHA和EPA可保障幼體的視覺(jué)系統(tǒng)發(fā)育等[34-35]。
4.2 哪種組織脂肪酸組成最適合用于指征食性?
Jeffries[36]發(fā)現(xiàn),某一海區(qū)生態(tài)系統(tǒng)中脂肪酸組成特征由該海區(qū)存在的浮游植物所決定,而每一個(gè)門類的浮游植物均由其特征脂肪酸指示。于此,本研究總結(jié)了拉氏南美南極魚(yú)體內(nèi)脂肪酸潛在的食物來(lái)源(表 1)。
對(duì)于肝臟組織來(lái)說(shuō),在兩個(gè)體長(zhǎng)組的樣品中,C14:0、C16:0、C16:1n7及C18:1n9這4種脂肪酸含量均較高,而PUFA并未體現(xiàn)出明顯特征,這與肝臟本身的功能有關(guān),其通常儲(chǔ)存大量的SFA和MUFA,而PUFA優(yōu)先供給其他組織。此結(jié)果似乎暗示拉氏南美南極魚(yú)可能會(huì)大量攝食硅藻、甲藻以及褐藻等,然而對(duì)比已有研究[2,7,9]可知,其胃含物中并未發(fā)現(xiàn)這些藻類的存在。另外,C15:0+C17:0、C18:2n6、C20:4n6的含量隨體長(zhǎng)增加而降低,暗示隨著拉氏南美南極魚(yú)的生長(zhǎng),其對(duì)于浮游細(xì)菌、海藻的攝食減少,且底棲攝食特性減弱。而基于已有胃含物分析[2]發(fā)現(xiàn),相對(duì)于小體長(zhǎng),大體長(zhǎng)魚(yú)類的攝食范圍較廣,且更多地?cái)z食底棲魚(yú)類。Rossi等[37]曾指出,DHA/EPA的比值越高,其指征的營(yíng)養(yǎng)級(jí)越高,在兩個(gè)體長(zhǎng)組中比值分別為0.59±0.20及0.47±0.11,說(shuō)明大體長(zhǎng)拉氏南美南極魚(yú)呈現(xiàn)出較低的營(yíng)養(yǎng)級(jí)水平。此外,本研究水域處于西南大西洋重要的魷魚(yú)漁場(chǎng)。已有多項(xiàng)研究顯示,拉氏南美南極魚(yú)胃中發(fā)現(xiàn)了漁業(yè)丟棄物的存在,如魷魚(yú)和各種經(jīng)濟(jì)魚(yú)種等[2,7-9]。綜合這些證據(jù)可知,該海域的拉氏南美南極魚(yú)應(yīng)處于一個(gè)較高的營(yíng)養(yǎng)級(jí)水平。據(jù)此,以肝臟組織中脂肪酸組成指示其食性會(huì)存在一定的誤差。
在兩個(gè)體長(zhǎng)組樣品的性腺組織中,C15:0+C17:0、C18:2n6、C20:1+C22:1及C20:4n6的貢獻(xiàn)率均較大,表現(xiàn)出大量攝食浮游細(xì)菌、陸源有機(jī)質(zhì)以及浮游動(dòng)物的情況,同時(shí)也表現(xiàn)出底棲性攝食特征。考慮到本研究的樣本基本未達(dá)性成熟,性腺內(nèi)儲(chǔ)存的脂肪酸大部分用于正常生長(zhǎng)。但性腺組織中DHA與EPA含量較低,且小體長(zhǎng)組中DHA/EPA為0.62±0.20,大體長(zhǎng)組中為0.46±0.15,對(duì)于食性的指征體現(xiàn)出大體長(zhǎng)的拉氏南美南極魚(yú)營(yíng)養(yǎng)級(jí)低于小體長(zhǎng)魚(yú)類,這與胃含物分析結(jié)果存在偏差。考慮到處于未成熟階段的個(gè)體性腺組織還未發(fā)育,對(duì)于DHA和EPA并未進(jìn)行選擇性儲(chǔ)存。此外,性腺組織在不同的發(fā)育階段對(duì)于不同類型脂肪酸的需求程度有異[16]。因此,利用性腺組織對(duì)其食性進(jìn)行指征也會(huì)出現(xiàn)一定的差異。
綜上分析,肝臟和性腺組織呈現(xiàn)的結(jié)果稍顯偏差,這與各組織對(duì)于脂肪酸的儲(chǔ)存及使用情況有關(guān)。對(duì)于拉氏南美南極魚(yú)來(lái)說(shuō),肌肉組織更新時(shí)間長(zhǎng),能夠反映較長(zhǎng)時(shí)間,甚至一生對(duì)食物中脂肪酸的吸收情況。
4.3 拉氏南美南極魚(yú)在生長(zhǎng)過(guò)程中如何轉(zhuǎn)換食性?
通過(guò)胃含物分析發(fā)現(xiàn),拉氏南美南極魚(yú)整個(gè)生活史過(guò)程中存在著食性的轉(zhuǎn)換[2,6-7,9]。在肌肉組織中,DHA與EPA的貢獻(xiàn)率較高,且小、大體長(zhǎng)組DHA/EPA值分別為0.35±0.16和0.36±0.13,說(shuō)明大體長(zhǎng)的拉氏南美南極魚(yú)體現(xiàn)出稍高的營(yíng)養(yǎng)級(jí)[37]。此外,Budge和Parrish[38]指出,在捕食者中只有植食性橈足類能夠通過(guò)其特有的途徑將C18:1和C20:1通過(guò)碳鏈延長(zhǎng)大量合成C20:1和C22:1。因此,C20:1和C22:1可作為橈足類攝食其他浮游植物的指示。Falk-Petersen等[26]提出,這兩種脂肪酸也可以作為高營(yíng)養(yǎng)級(jí)捕食者攝食植食性橈足類的指示。在本研究中,大體長(zhǎng)拉氏南美南極魚(yú)體內(nèi)C20:1+C22:1含量高于小體長(zhǎng)魚(yú)類,且隨著體長(zhǎng)的增加,其含量呈顯著性增加趨勢(shì)。由此推測(cè),隨著拉氏南美南極魚(yú)的生長(zhǎng),其攝食以植食性橈足類為食的生物的程度有所增加。就C18:2n6來(lái)說(shuō),曾有研究提出其為一種典型的陸源脂肪酸[25],其在肌肉組織脂肪酸中貢獻(xiàn)率較大。這表明,拉氏南美南極魚(yú)在巴塔哥尼亞陸架區(qū)對(duì)于陸源有機(jī)物或以其為食物的生物有一定的攝食。考慮到拉氏南美南極魚(yú)在近岸和大洋水域均有分布,而本研究區(qū)域位于陸架區(qū)。因此,這種陸源有機(jī)物可能由東福克蘭流由福克蘭群島或阿根廷漂流由阿根廷近岸攜帶至本研究海區(qū)[34],也有可能是本研究區(qū)域的拉氏南美南極魚(yú)在近岸水域攝食,然后洄游至陸架區(qū)所致[39]。此外,大體長(zhǎng)拉氏南美南極魚(yú)體內(nèi)C18:2n6含量高于小體長(zhǎng)魚(yú)類,說(shuō)明大體長(zhǎng)魚(yú)類洄游能力高于小體長(zhǎng)魚(yú)類,攝食范圍也隨著增加。飽和脂肪酸中,C15:0和C17:0可用來(lái)指示食物中細(xì)菌的貢獻(xiàn)[22],肌肉組織中C15:0+C17:0也表現(xiàn)出明顯的特征,其大體長(zhǎng)拉氏南美南極魚(yú)體內(nèi)含量較高,由此推測(cè)拉氏南美南極魚(yú)在攝食浮游生物過(guò)程中可能順帶攝入浮游細(xì)菌。總的來(lái)講,肌肉組織脂肪酸表現(xiàn)出的特征表明拉氏南美南極魚(yú)呈現(xiàn)出浮游生物食性[40]。另一方面,肌肉組織中C20:4n6含量較高,貢獻(xiàn)率較大,是能夠指征底棲生物食性的脂肪酸[26]。由此可見(jiàn),拉氏南美南極魚(yú)能夠較易地在浮游及底棲生活之間轉(zhuǎn)換。該結(jié)果與Kock[41]的分析一致。這也是為什么拉氏南美南極魚(yú)肌肉中δ15N和δ13C含量與體長(zhǎng)之間無(wú)線性關(guān)系存在的原因[39]。針對(duì)PUFA,主要由n-3系列和n-6系列脂肪酸組成,其中n-3系列脂肪酸可以指示浮游植物通過(guò)捕食食物鏈向更高營(yíng)養(yǎng)級(jí)捕食者傳遞[42],而n-6系列脂肪酸中,C20:4n6所占比重最高。所以,n-6 PUFA/n-3 PUFA的比值被用來(lái)估計(jì)腐生食物鏈和捕食食物鏈對(duì)于高營(yíng)養(yǎng)級(jí)捕食者的貢獻(xiàn)程度[42]。在拉氏南美南極魚(yú)體內(nèi),n-6 PUFA/n-3 PUFA的比值與體長(zhǎng)之間并無(wú)明顯的線性關(guān)系。這可能是因?yàn)榇蟆⑿€(gè)體拉氏南美南極魚(yú)胃含物中均發(fā)現(xiàn)了漁業(yè)丟棄物,從而導(dǎo)致腐生食物鏈和捕食食物鏈的貢獻(xiàn)率特征弱化。此外,該現(xiàn)象也可能是因?yàn)殡S著生長(zhǎng),其在浮游與底棲環(huán)境之間頻繁轉(zhuǎn)換所致。
在本研究中,隨著拉氏南美南極魚(yú)的生長(zhǎng),其對(duì)于浮游細(xì)菌、浮游動(dòng)物以及底棲魚(yú)類的攝食均會(huì)增加,表明其攝食特性由浮游性攝食方式轉(zhuǎn)變?yōu)楦∮?底棲性攝食,且增強(qiáng)對(duì)于陸源有機(jī)質(zhì)的攝食,推測(cè)其洄游能力有所增強(qiáng)。但由于漁業(yè)丟棄物的影響,導(dǎo)致其營(yíng)養(yǎng)級(jí)水平升高,腐生食物鏈和捕食食物鏈的貢獻(xiàn)率特征弱化。從這個(gè)角度來(lái)看,人類活動(dòng),尤其是漁業(yè)活動(dòng),可能會(huì)影響或干擾大洋性生態(tài)系統(tǒng)結(jié)構(gòu)。
致謝:本研究所使用樣品均為中國(guó)水產(chǎn)有限公司所屬漁船“龍騰”號(hào)船長(zhǎng)及全體船員協(xié)助收集,實(shí)驗(yàn)過(guò)程在上海海洋大學(xué)極地研究中心和大洋漁業(yè)資源可持續(xù)開(kāi)發(fā)教育部重點(diǎn)實(shí)驗(yàn)室完成,在此深表謝忱!
