海洋自然資本評估中生物能值轉(zhuǎn)換率的研究
2020-05-21 13:15:12丁潔瓊魏方懿葉觀瓊曾江寧
應(yīng)用海洋學(xué)學(xué)報 2020年2期
丁潔瓊,魏方懿,葉觀瓊*,曾江寧,林 穎,韓 宇
(1.浙江大學(xué)海洋學(xué)院,浙江 舟山 316021; 2.自然資源部第二海洋研究所,浙江 杭州 310012; 3.浙江省水利河口研究院,浙江 杭州 310020)
目前,研究自然資本評估主要有三大理論體系,包括“生態(tài)足跡”、“生態(tài)系統(tǒng)服務(wù)”和 “能值分析理論”[1-3]。其中,前兩者均是以人類為導(dǎo)向?qū)ψ匀画h(huán)境提供的資源價值進行評估[4],較少關(guān)注自然資本中未來可能提供的產(chǎn)品服務(wù)流及環(huán)境資產(chǎn)的自身存量,因此較難系統(tǒng)完整地反映自然資本價值。能值分析理論則是基于生態(tài)系統(tǒng)中生物鏈概念和熱力學(xué)原理提出的,關(guān)注自然資源本身,通過能值轉(zhuǎn)換率將生態(tài)系統(tǒng)內(nèi)流動和儲存的各種不同類別的能量和物質(zhì)轉(zhuǎn)換為同一標準的能值,可衡量和比較不同等級的能量價值,包涵了物質(zhì)過去、現(xiàn)在所有的信息,能系統(tǒng)完整地反映自然資本價值。就海洋生物本身來說,能值分析理論有其獨特優(yōu)勢,可體現(xiàn)能量在生物之間的流動及其關(guān)系,而且評估結(jié)果不以人類的價值體系為轉(zhuǎn)移,從而能夠客觀認識不同物種的生態(tài)價值。
由于海洋環(huán)境具有復(fù)雜、開放、流動、均質(zhì)等屬性,海洋自然資本能值評估方法尚處于探索階段[5]。現(xiàn)有的研究多關(guān)注海岸帶,包括灘涂[6]、海草床[7]、紅樹林[8-9]和濕地[10]等,綜合性的海灣和海島海域[11]研究較少。而能值轉(zhuǎn)換率是海洋生態(tài)系統(tǒng)能值計算中最核心的一個參數(shù),它將一系列能量轉(zhuǎn)換過程中各種形式的能量聯(lián)系了起來。海洋生物作為海洋生態(tài)系統(tǒng)的重要組成部分,其能值轉(zhuǎn)換率這一關(guān)鍵要素的研究尚處于初級階段。本研究基于海洋食物網(wǎng)信息論[12-13]和食物網(wǎng)營養(yǎng)級研究[14-16]的發(fā)展,探索海洋生態(tài)系統(tǒng)中不同營養(yǎng)級之間能值轉(zhuǎn)換率的計算,旨在為海洋生物自然資本量化提供關(guān)鍵參數(shù)。
1 研究現(xiàn)狀
1.1 能值轉(zhuǎn)換率
能值分析理論與方法是美國著名生態(tài)學(xué)家Odum(1986)在能量系統(tǒng)分析的基礎(chǔ)上發(fā)展形成的[17]。能值是直接或間接用于形成某產(chǎn)品或服務(wù)的一種有效能。考慮到任何能量都始于太陽,實際應(yīng)用中以太陽能值來衡量某一能量的能值。能值轉(zhuǎn)換率是實現(xiàn)能量和能值之間轉(zhuǎn)換的橋梁,實際使用的是太陽能值轉(zhuǎn)換率,即形成每單位某種產(chǎn)品(能量或物質(zhì))所需的能值量[18]。其計算公式[3]如下:
式(1)中:T為某種物質(zhì)(能量)的太陽能值轉(zhuǎn)換率(sej/J),E(t)為直接或間接應(yīng)用于形成該物質(zhì)的總太陽能值(sej),N為該物質(zhì)或能量的量(J)。
能量在傳遞過程中衰竭,而能值不變,因此其所含的能值轉(zhuǎn)換率不斷增加。能值轉(zhuǎn)換率是一種有量綱的效率比,用以衡量能量的質(zhì)量即其在整個能量等級中的地位,其值越大表明該能量的等級越高。
能值轉(zhuǎn)換率的計算是能值分析的基礎(chǔ)和最大難點,也是能值分析理論得到廣泛認可、應(yīng)用和進一步發(fā)展的必要條件[19]。國內(nèi)外學(xué)者對具體能值轉(zhuǎn)換率的計算開展了廣泛研究。生物能值轉(zhuǎn)換率的計算一般通過繪制能流圖[20],包含能量進出、儲存、分散等各種途徑,將已知系統(tǒng)投入的總太陽能值除以流入各組分能量得到系統(tǒng)內(nèi)各對象的能值轉(zhuǎn)換率。李璇等(2016)通過定量分析舟山大黃魚(Larimichthys crocea)養(yǎng)殖系統(tǒng)的輸入能值和輸入熱值,得到大黃魚的能值轉(zhuǎn)換率[21]。化石能源及其衍生物的能值轉(zhuǎn)換率可通過分析其生成的地球化學(xué)過程[22]、實地考察煉油工藝[23]計算得到。賀成龍(2016)在考慮水電工程建設(shè)主要投入產(chǎn)出的基礎(chǔ)上計算出水力發(fā)電的能值轉(zhuǎn)換率[24]。縱觀前人研究,能值轉(zhuǎn)換率的計算方法以投入產(chǎn)出法為主,此外還有基于最小特征值模型[25-26]、線性優(yōu)化模型、矩陣模型[27]等方法。基于目前靜態(tài)的研究現(xiàn)狀,動態(tài)的能值轉(zhuǎn)換率研究和完善是下一步的研究熱點[28]。
1.2 海洋生物能值研究現(xiàn)狀
在開創(chuàng)能值分析理論后,Odum等(1991)對封閉的蝦類養(yǎng)殖池進行相關(guān)研究,通過投入的總太陽能值(可更新資源與不可更新資源能值之和)與產(chǎn)出蝦的能量之比得到養(yǎng)殖蝦的能值轉(zhuǎn)換率[29]。Brown等(1993)[31]依據(jù)McRoy等(1992)研究[30]中一系列營養(yǎng)網(wǎng)絡(luò)和能量流構(gòu)建了一個詳細的威廉王子灣營養(yǎng)網(wǎng)絡(luò)模型。結(jié)合林德曼效率,將威廉王子灣營養(yǎng)網(wǎng)絡(luò)模型中的各種能量流輸入NETWRK3軟件[32],得到不同物種的營養(yǎng)級和高營養(yǎng)級物種的能值轉(zhuǎn)換率,同一營養(yǎng)層級的生物能值轉(zhuǎn)換率相同。由NETWRK3軟件得出的營養(yǎng)級與DeGange等(1987)的研究結(jié)果[33]相差很小,但是同一物種的能值轉(zhuǎn)換率在取不同的林德曼效率(5%~30%)計算時,其值相差近一個數(shù)量級,在高營養(yǎng)級物種中差異更明顯。該營養(yǎng)網(wǎng)絡(luò)模型在缺乏能量流定量描述的情況下,將林德曼效率的值設(shè)為10%,納入能值轉(zhuǎn)換率的核算。但是在海洋生態(tài)系統(tǒng)中林德曼效率的變化范圍很大,實際能量流的傳遞效率可能顯著不同于文中核算結(jié)果。
Brown等(2006)基于食物網(wǎng)對佛羅里達大沼澤地的生物多樣性進行分析[34],將簡單的線性優(yōu)化方法[35]應(yīng)用于矩陣中計算禾本科沼澤濕地各組分的能值轉(zhuǎn)換率。能值轉(zhuǎn)換率的準確性取決于源數(shù)據(jù)的有效性,不同季節(jié)計算出的能值轉(zhuǎn)換率有明顯差異,高能值轉(zhuǎn)換率往往出現(xiàn)于濕季。雖然系統(tǒng)中每個組分計算的能值轉(zhuǎn)換率存在相當大的不確定性,但與其他方法所得結(jié)果相比有很高的一致性。Vassallo等(2007、2009)[36-37]在分析近海魚類養(yǎng)殖系統(tǒng)過程中運用Odum等(1996)的能值代數(shù)法和投入產(chǎn)出法[3],對以年為期進行能值分析的方法進行改進,構(gòu)建了金頭鯛(Sparus aurata)水產(chǎn)養(yǎng)殖能值瞬時分析模型。上述兩個研究案例表明,時間單位、投入的產(chǎn)品類別和服務(wù)以及產(chǎn)出效率,會對計算結(jié)果產(chǎn)生較大影響,如金頭鯛的年能值轉(zhuǎn)換率為1.32×106sei/J,其春季的能值轉(zhuǎn)換率則為5.55×105sei/J。目前,海洋生物能值轉(zhuǎn)換率的方法主要有投入產(chǎn)出法、能流圖法和線性優(yōu)化法,但相關(guān)研究涉及的海洋生物種類并不多,部分海洋生物的能值轉(zhuǎn)換率如表1所示。
2 基于食物網(wǎng)信息論的能值轉(zhuǎn)換率
2.1 食物網(wǎng)信息論
信息論作為一門學(xué)科,通常被認為是從二戰(zhàn)期間的破譯密碼行為中發(fā)展形成的。信息論普及后不久,生態(tài)學(xué)家Macarthur(1955)闡明了信息論及相關(guān)概念在生態(tài)流網(wǎng)絡(luò)的適用性[40]。Ulanowicz(1986)認為信息論對定量生態(tài)學(xué)十分重要,提出了多個系統(tǒng)生長和發(fā)育的信息論指標用于分析食物網(wǎng)的大小和復(fù)雜性[41];并總結(jié)了信息論在生態(tài)學(xué)中的應(yīng)用,不但可以用來量化種群分布和生物數(shù)量,還可以定量研究營養(yǎng)過程相互作用的模式[42]。Moniz等(2007)利用豐度時間序列上的信息論技術(shù)試圖重構(gòu)食物網(wǎng)的拓撲結(jié)構(gòu),發(fā)現(xiàn)信息統(tǒng)計數(shù)據(jù)能正確識別簡單模型的食物網(wǎng)拓撲結(jié)構(gòu),比如驗證消費者對某種食物的偏好[43]。在生態(tài)環(huán)保方面,食物網(wǎng)理論為建立一整套基于生態(tài)系統(tǒng)的海洋資源管理方法奠定了基礎(chǔ)。
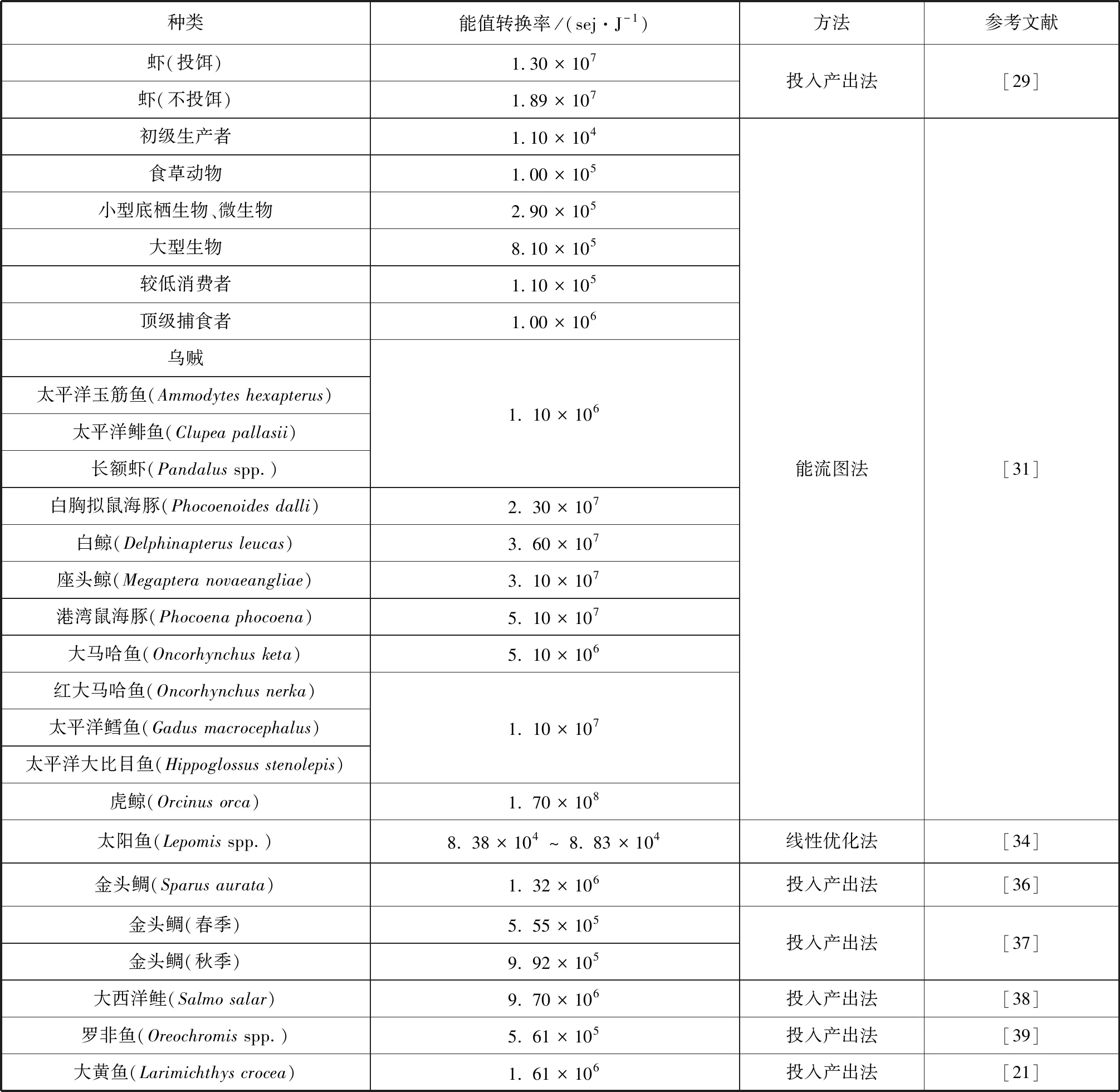
表1 部分海洋生物能值轉(zhuǎn)換率及方法Tab.1 Transformities for some marine organisms
2.2 海洋生物能值轉(zhuǎn)換率經(jīng)驗公式
現(xiàn)階段海洋食物網(wǎng)能量傳遞效率和生物能值研究較少,大部分仍然沿用Brown等計算的浮游植物、浮游動物、草食性魚類、哺乳動物、頂級捕食者等海洋生物大類的能值轉(zhuǎn)換率[31]。為方便且準確的估算海洋自然資本能值價值,我們參考文獻[44],構(gòu)建了海洋食物網(wǎng)信息能流圖(圖1),圖中假定每年輸入食物網(wǎng)的總初級生產(chǎn)力為1 000 kcal/m2,外源有機質(zhì)輸入為100 kcal/m2。最后結(jié)合生態(tài)系統(tǒng)能量傳遞規(guī)律,建立生物營養(yǎng)級與其能值轉(zhuǎn)換率的經(jīng)驗公式。
根據(jù)生態(tài)系統(tǒng)能量流動的基本原理[45],第n營養(yǎng)級生物固定的能量為
式(2)中:Nn為第n級營養(yǎng)生物固定的能量(J),N1為初級生產(chǎn)者固定的能量(J),EL為林德曼效率。
假定能流網(wǎng)處于平衡、穩(wěn)定的狀態(tài),處于不同營養(yǎng)級的生物都能充分利用太陽能值總量,則第n營養(yǎng)級生物的能值轉(zhuǎn)換率公式如下:
E=N1·T1
(4)
式(3、4)中:Tn為第n營養(yǎng)級生物的能值轉(zhuǎn)換率(sej/J),E為流入整個系統(tǒng)的太陽能值總量(sej),T1為初級生產(chǎn)者的能值轉(zhuǎn)換率(sej/J)。
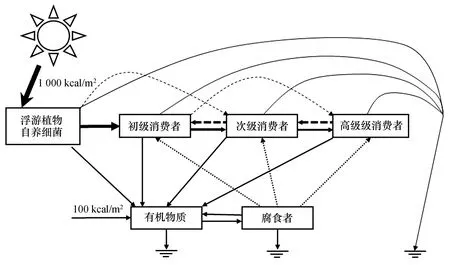
圖1 食物網(wǎng)信息能流圖Fig.1 Energy flow chart in a food web
由公式(2~4)建立生物營養(yǎng)級與其能值轉(zhuǎn)換率之間的經(jīng)驗關(guān)系,推導(dǎo)出海洋生物能值轉(zhuǎn)換率的經(jīng)驗公式如下:
根據(jù)公式(5),由初級生產(chǎn)者的太陽能值轉(zhuǎn)換率、林德曼效率和營養(yǎng)級數(shù)據(jù),可估算出某一物種的太陽能值轉(zhuǎn)換率。前人的研究方法需要建立食物網(wǎng)模型,計算系統(tǒng)每一成分間流動的能量,而經(jīng)驗公式簡化了能值轉(zhuǎn)換率的計算過程。
2.3 案例應(yīng)用
根據(jù)生物能值轉(zhuǎn)換率的相關(guān)研究,同時考慮數(shù)據(jù)的可獲得性,選取表2中的生物進行案例研究,以Brown等的研究結(jié)果[31]為參考,其初級生產(chǎn)者的能值轉(zhuǎn)換率為1.1×104sei/J,林德曼效率EL的取值則為10%。
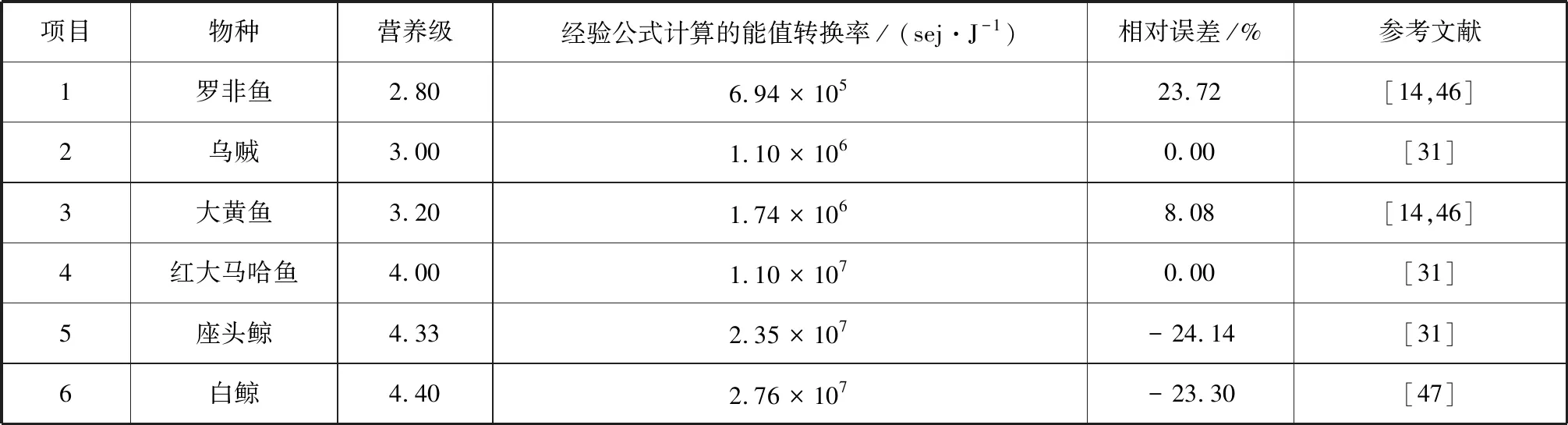
表2 經(jīng)驗公式使用案例Tab.2 A case of using empirical formula
根據(jù)本研究提出的經(jīng)驗公式,計算得到表2中生物能值轉(zhuǎn)換率,與表1中對應(yīng)值相比,其相對誤差的絕對值在0%~25%之間。海洋生態(tài)系統(tǒng)的平均生態(tài)效率通常比陸地的高,植食性動物生態(tài)效率是20%左右,較高營養(yǎng)級之間的生態(tài)效率可能在10%~15%范圍內(nèi)[45]。由于部分海區(qū)缺乏調(diào)查研究,參考現(xiàn)有相關(guān)營養(yǎng)級數(shù)據(jù),從而會產(chǎn)生一定誤差。Campbell等(2005)認為不同方法計算出的能值轉(zhuǎn)換率保持在同一數(shù)量級即可,能值分析的最終目標是將計算結(jié)果與實際值的誤差控制在10%以內(nèi)[48],表明本研究經(jīng)驗公式的適用性。基于海洋生物營養(yǎng)級相關(guān)研究[14,46]、能值轉(zhuǎn)換率的計算結(jié)果(表2)以及本研究提出的經(jīng)驗公式,可得到海洋食物網(wǎng)不同食性生物所處營養(yǎng)級及能值轉(zhuǎn)換率(表3),為海洋能值研究提供一定參考。其中,海洋生物營養(yǎng)位置變化與其生長發(fā)育階段[49]、生態(tài)系統(tǒng)類型、捕食策略等因素有關(guān)[50],全球氣候變化、極端氣候事件[51]和人類活動也會對其產(chǎn)生一定影響。具體應(yīng)用時根據(jù)實際調(diào)查數(shù)據(jù)確定海洋生物的營養(yǎng)級,在缺乏數(shù)據(jù)時選擇鄰近且海洋生態(tài)系統(tǒng)相似海區(qū)的數(shù)據(jù)替代,同時相應(yīng)調(diào)整林德曼效率,以提高計算結(jié)果的科學(xué)性。
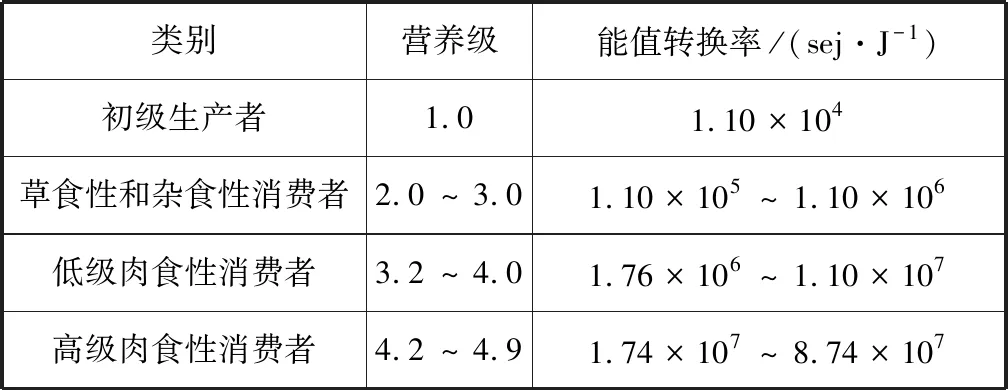
表3 食物網(wǎng)營養(yǎng)關(guān)系及能值轉(zhuǎn)換率Tab.3 Nutritional relationship in a food web and emergy transformity
2.4 討論與應(yīng)用前景
自然資源的價值評估具有復(fù)雜性且涉及許多假設(shè),目前的研究大都以人類視角對生態(tài)系統(tǒng)服務(wù)功能價值進行評估,而生態(tài)系統(tǒng)內(nèi)在價值評估方法和定量化的研究不足[52]。基于該現(xiàn)狀,本研究構(gòu)建海洋生物能值轉(zhuǎn)換率計算的經(jīng)驗公式以期精確量化海洋生態(tài)系統(tǒng)內(nèi)在價值,該公式主要由3部分組成,一是初級生產(chǎn)者的能值轉(zhuǎn)換率,二是物種所處的營養(yǎng)級,三是林德曼效率。由于缺乏營養(yǎng)級間能量流傳遞效率、食物鏈的反饋機制等相關(guān)數(shù)據(jù)和研究,本研究在經(jīng)驗公式中取林德曼效率為10%,該值目前已廣泛應(yīng)用于海洋漁業(yè)生產(chǎn)[53]、水產(chǎn)品網(wǎng)箱養(yǎng)殖產(chǎn)量估算等方面。
然而海洋食物網(wǎng)是復(fù)雜的、動態(tài)的,需要進一步考慮林德曼效率的適用性[54]及明確物種所處的營養(yǎng)級[55]。海洋物種在食物網(wǎng)中的營養(yǎng)位置,最初是通過體型大小判斷、胃含物分析法[56]等方法確定其營養(yǎng)級,而后碳氮穩(wěn)定同位素分析法提供了估算營養(yǎng)級的新方法。目前同位素分析法廣泛應(yīng)用于陸地、鹽沼濕地[57]、河口、海灣[58]、近海[59]及大洋生態(tài)系統(tǒng)食物網(wǎng)和營養(yǎng)級生態(tài)位變化研究。隨著DNA測序技術(shù)的發(fā)展,DNA條形碼法[60]也已應(yīng)用于海洋生物食性分析。但上述方法都存在一定的局限性,今后可運用綜合性研究方法確定營養(yǎng)級關(guān)系以及能量流動,同時可將能值分析理論與地理信息系統(tǒng)(GIS)等工具相結(jié)合,有助于客觀理解海洋生態(tài)系統(tǒng)能量代謝的動態(tài)變化過程[61]。還可借鑒國際魚類數(shù)據(jù)庫(Fishbase)的經(jīng)驗,構(gòu)建信息全面、實時更新、開放共享的全球海洋生物多樣性數(shù)據(jù)庫[62],完善海洋食物網(wǎng)模型和食物網(wǎng)拓撲結(jié)構(gòu),便于開展能值轉(zhuǎn)換率的精細化研究。
3 結(jié)論
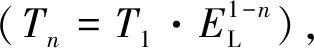
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農(nóng)業(yè)(2021年11期)2021-08-13 08:53:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
甘肅教育(2020年14期)2020-09-11 07:57:42
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
海峽姐妹(2016年5期)2016-02-27 15:20:20
時代英語·高二(2015年1期)2015-03-16 00:08:11
