升溫與硝酸鹽加富對鹿角杯形珊瑚幼蟲的生理影響
2020-05-21 13:15:02孫有方俞曉磊
應用海洋學學報 2020年2期
張 芳,孫有方,俞曉磊,江 雷,黃 暉,4*
(1.中國科學院南海海洋研究所、中國科學院熱帶海洋生物資源與生態重點實驗室,廣東 廣州 510301; 2.中國科學院南海生態環境工程創新研究院,廣東 廣州 510301; 3.中國科學院大學,北京 100049; 4.中國科學院海南熱帶海洋生物實驗站,海南 三亞 572000)
珊瑚礁生態系統正面臨全球氣候變化與人類活動影響的巨大威脅,而日益升高的表層海水溫度與營養鹽富集是其中的關鍵驅動因子[1]。與工業革命前相比,全球平均海水表層溫度已經升高了將近0.74 ℃[2],根據聯合國政府間氣候變化專門委員會(Intergovernmental Panel on Climate Change,IPCC)模型預測截止到本世紀末海水表層溫度將持續上升2~3 ℃[3],日益升高的表層海水溫度對珊瑚的威脅將更加劇烈。在珊瑚礁破壞、紅樹林砍伐、近岸污染物排放以及生活污水的暗排等人類活動的嚴重干擾下,珊瑚礁區海域的無機營養鹽負荷將嚴重超載[4]。近年來,越來越多的學者對近岸珊瑚礁生態系統所面臨的表層海水溫度升高和營養鹽加富這兩大主要環境壓力進行關注,但主要集中于成體珊瑚。研究表明,造礁石珊瑚的熱耐受性在水質環境不佳的情況下有所降低[5-8],表層海水溫度升高和營養鹽加富的疊加或協同作用可能會導致珊瑚覆蓋率的顯著下降[9]。但是,也有研究表明無機氮加富可以增加珊瑚共生體色素濃度[10],增強固氮細菌固氮能力[11],減緩熱應激對珊瑚的負面影響[12]。與珊瑚成體相比,珊瑚幼蟲的浮游期較長、營養物質來源單一、細胞防御機制不成熟、生物量及相對表面積小,對環境壓力更加敏感[13]。目前為止,大多數有關于珊瑚早期發育的研究都聚焦于海水升溫對珊瑚存活率、傳播距離和附著率等生理影響[14-20],僅有少數研究關注于升溫與營養鹽加富對珊瑚幼蟲的共同作用[21-23],結果表明不同種類珊瑚幼蟲對升溫和營養鹽加富的響應與幼蟲是否含有蟲黃藻有關,營養鹽加富對不含蟲黃藻的排卵型珊瑚幼蟲影響較小[23]。同時,也有研究表明珊瑚幼蟲對營養鹽加富具有一定的耐受性。雖然,該過程涉及的機制尚不清晰,但可能與谷氨酰胺合成酶或谷氨酰胺脫氫酶對過量氨的解毒能力有關[24]。進一步研究升溫與營養鹽加富對珊瑚幼蟲的影響有助于預測和闡明兩者對珊瑚種群補充的潛在影響。
三亞旅游業的快速發展導致人口數量急劇增加使得陸源輸入對三亞灣的影響顯著增強,迫使鹿回頭珊瑚礁區的營養鹽含量升高,存在富營養化的隱患。而硝酸鹽作為無機氮的主要存在形式,其含量的增加是導致海洋富營養化的主要因素之一。鹿角杯形珊瑚(Pocilloporadamicornis)繁殖方式為孵幼型,是鹿回頭海域的優勢種和印度太平洋海區的廣布種。在三亞灣海域受氣候變化與陸源氮源污染影響愈來愈重的雙重環境背景下,本研究通過室內培養實驗探究了升溫與硝酸鹽加富對鹿角杯形珊瑚幼蟲的附著、共生蟲黃藻熒光指數及代謝速率的影響,以了解鹿角杯形珊瑚幼蟲對升溫與海水富營養化共同作用的響應。
1 材料與方法
1.1 珊瑚幼蟲收集
2018年9月14日于中國科學院海南熱帶海洋生物實驗站旁鹿回頭(18°12.7′N,109°28.5′E)岸礁2~3 m水深處采集10株大小相近(直徑15 cm左右),顏色相似的成熟鹿角杯形珊瑚母體,在實驗站珊瑚養殖缸中暫養24 h后,將10株母體珊瑚分別置于10個配有幼蟲收集器的流水式塑料桶(18 dm3)中,9月16日早上7時將各收集器中的幼蟲收集于燒杯中,混合均勻后隨機挑選用于實驗。
1.2 實驗設計
本實驗中,溫度設常溫(29 ℃)與升溫(31 ℃)兩個水平,硝酸鹽濃度設置0、5、20 μmol/dm33個梯度,分別表示不加富、低濃度加富和高濃度加富。29 ℃是鹿回頭海域夏季月均溫[25-26],而31 ℃高于鹿回頭珊瑚群落白化閾值的1~2 ℃,同時也在IPCC預測的升溫范圍內[25]。雙因子全交互設計下共有6個實驗組,編號為1、2、3、4、5、6,分別代表常溫未加富組、常溫低濃度加富組、常溫高濃度加富組、升溫未加富組、升溫低濃度加富組和升溫高濃度加富組。1號對照組為0.5 μm過濾海水,硝酸鹽加富組為分析純KNO3配置的相應濃度的海水。實驗在一個44 cm×66 cm×24 cm的塑料實驗缸中進行,光照強度約為300 μmol photons/(m2·s),12 h照明(6:30至18:30)。數顯式溫度控制器和鈦合金加熱棒控制溫度,小型潛水泵維持實驗缸水體流動,60 cm3玻璃瓶和800 cm3塑料瓶培養珊瑚幼蟲。每個溫度設3個重復實驗缸,每個實驗缸設3個硝酸鹽濃度,每個濃度2個重復,因此每個處理共設置了6個重復,實驗周期為5 d。使用AA3連續流動分析儀檢測實驗用水的硝酸鹽濃度,各實驗組溫度和硝酸鹽濃度以實際測量值為準(表1)。
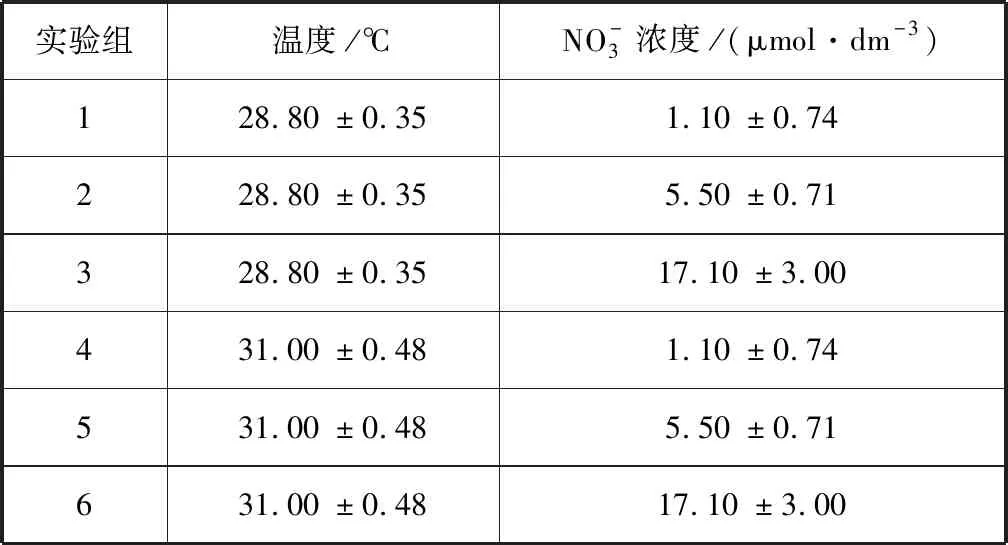
表1 各實驗組溫度和硝酸鹽濃度Tab.1 Nitrate concentration and temperature for the treatments
注:數值以平均值±標準差表示。
1.3 珊瑚幼蟲存活率、附著率和葉綠素熒光指數(Fv/Fm)
用于統計存活率的珊瑚幼蟲在60 cm3透明玻璃管中單獨培養,每根玻璃管20個幼蟲,每個處理4個重復,每天觀察珊瑚幼蟲狀態,記錄管中幼蟲數目(n=4)。實驗第5天,分別從塑料瓶中隨機選取60個幼蟲,加入聚丙乙烯六孔板中,每孔10個幼蟲、1片珊瑚藻和15 cm3相應處理的海水,12 h后統計成功附著珊瑚幼蟲個數(n=6)。使用水下葉綠素調制熒光儀(Diving-PAM)對暗適應1 h后珊瑚幼蟲進行葉綠素熒光指數(Fv/Fm)的測定(n=6),測量方法參照文獻[16]。
1.4 代謝速率
實驗第5天進行幼蟲凈光合速率(Net Photosynthesis,Pnet)和暗呼吸速率(Dark Respiration,RD)測定(n=4)。從塑料瓶中隨機挑選20個幼蟲和一個3 mm×5 mm的轉子放入2 cm3內壁貼有氧氣光學感應貼片的玻璃瓶,封瓶后置于相同實驗環境,測量前校正。連接溶氧測量儀(Presens,OXY-4 mini),打開攪拌裝置,轉速約為300 r/min。測定光照條件下10 min內的氧氣含量變化,利用線性擬合計算單位時間內每個幼蟲的產氧速率,即為珊瑚幼蟲的凈光合速率(Pnet)。珊瑚幼蟲暗呼吸速率(RD)于暗適應2 h后測量,方法與凈光合速率相同。珊瑚幼蟲總光合速率(Gross Photosynthesis,PG)等于凈光合速率與暗呼吸速率之和。光合呼吸速率比即PG和RD之間的比值(PG/RD)可以反映珊瑚幼蟲有機碳固定能力,當PG/RD≥2時,說明共生藻光合作用固定的凈有機碳完全可以支撐黑暗條件下呼吸作用的消耗,即共生體有機碳凈累積;當PG/RD<2時,則說明幼蟲暗呼吸消耗的有機碳大于光照下共生藻光合作用累積的有機碳,即幼蟲的內源能量物質出現凈消耗[27]。
1.5 數據處理與分析
實驗數據以平均值±標準差(mean±SD)表示,用SPSS 21.0數據處理軟件進行雙因素方差分析和Duncan多重比較,實驗結果用SigmaPlot 11.0作圖。
2 結果與討論
2.1 珊瑚幼蟲存活率和共生蟲黃藻熒光指數
培養5 d后,各處理組珊瑚幼蟲的存活率均維持在90%左右[圖1(a)],共生蟲黃藻的熒光指數(Fv/Fm)在0.65左右[圖1(b)],各處理組之間無顯著差異(p>0.05)。結果表明,在本實驗中珊瑚幼蟲存活率和共生蟲黃藻熒光指數(Fv/Fm)既不受硝酸鹽濃度也不受溫度的影響,且溫度與硝酸鹽濃度之間無交互作用(p>0.05)。
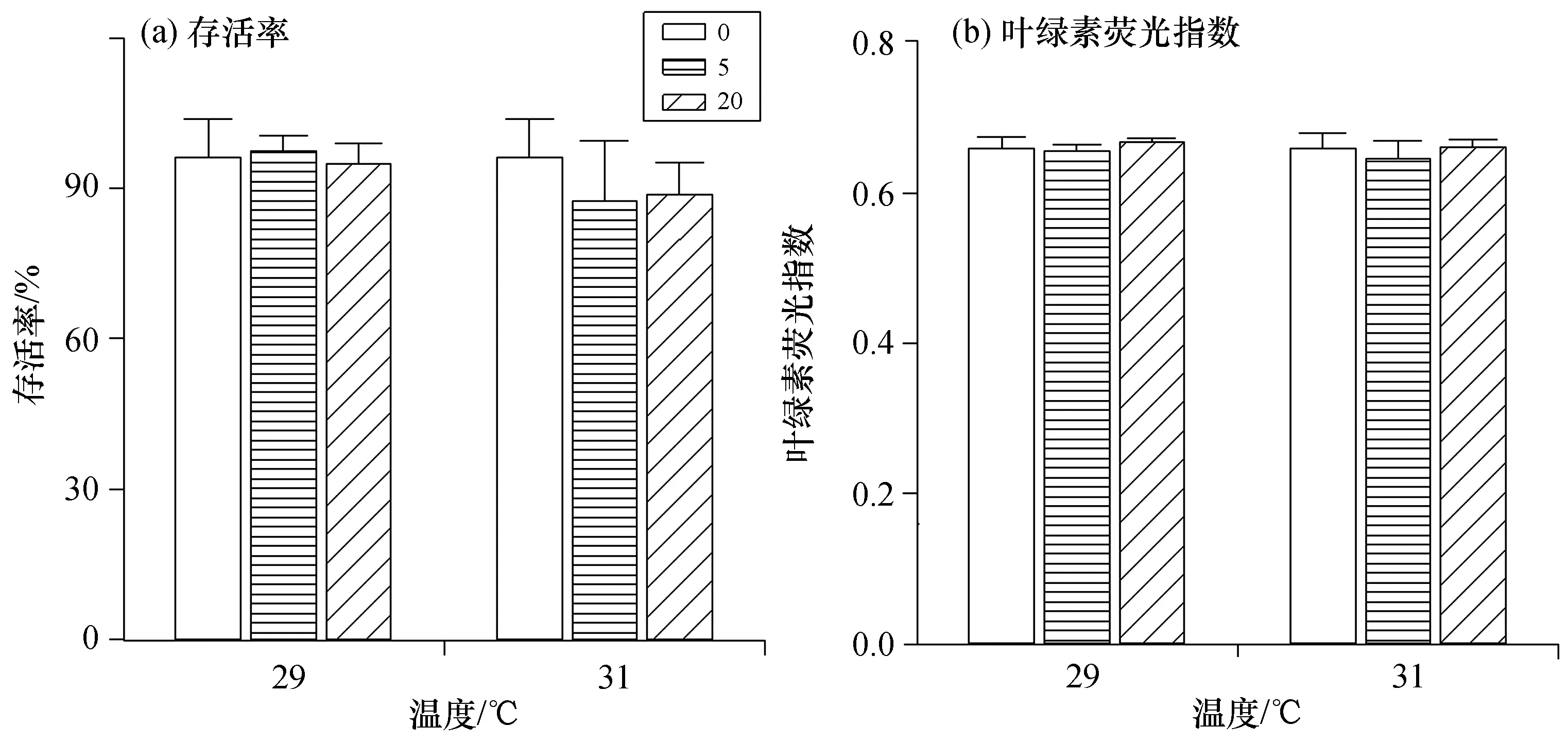
圖1 鹿角杯形珊瑚幼蟲存活率與共生蟲黃藻熒光指數Fig.1 Survival rate and Fv/Fm of Pocillopora damicornis larvae in 5 days by the treatments 圖例數值0、5、20分別表示相應的硝酸鹽濃度,單位為μmol/dm3。
2.2 珊瑚幼蟲附著率
珊瑚幼蟲的附著率在升溫時顯著降低(p<0.05)。與實驗組1、2、3相比較,實驗組4、5、6的附著率在31 ℃時分別顯著下降了14.3%、23.3% 和48.2%(p<0.05)。常溫條件下幼蟲附著率不受硝酸鹽濃度的影響(p>0.05);但升溫條件下附著率隨硝酸鹽濃度的增加而降低,且6號實驗組顯著降低(p<0.05)。硝酸鹽濃度與溫度對幼蟲附著率的影響表現為明顯的交互作用(p<0.05),即硝酸鹽加富加重了升溫對珊瑚幼蟲附著率的負面影響(圖2)。
2.3 珊瑚幼蟲代謝速率
溫度與硝酸鹽濃度對鹿角杯形珊瑚幼蟲代謝的影響如圖3所示。溫度和硝酸鹽濃度對珊瑚幼蟲Pnet的影響存在著顯著交互作用(p<0.05)。升溫條件下,實驗組4珊瑚幼蟲Pnet顯著高于實驗組1(p<0.05),但實驗組5和6的Pnet不受溫度影響(p>0.05)。硝酸鹽加富對珊瑚幼蟲Pnet的影響在不同溫度條件下有所差異,5 μmol/dm3的硝酸鹽加富顯著促進了珊瑚幼蟲在常溫條件下的Pnet,增長率113.3%(p<0.05);而20 μmol/dm3的硝酸鹽加富顯著抑制了珊瑚幼蟲在升溫條件下的Pnet,下降率33.3%(p<0.05)。同樣,珊瑚幼蟲的RD在溫度與硝酸鹽濃度之間具有顯著交互作用(p<0.05)。升溫時,實驗組4和6增加率分別為82.7%和45.1%(p<0.05),而實驗組5的RD無顯著變化(p>0.05)。與對照組相比,實驗組2的RD顯著升高,但實驗組5顯著降低(p<0.05);實驗組3的RD顯著升高,但實驗組6無顯著變化(p>0.05);與珊瑚幼蟲RD相同,0、20 μmol/dm3硝酸鹽加富組珊瑚幼蟲PG對海水升溫具有顯著正面影響(p<0.05),而5 μmol/dm3加富組不受溫度影響[圖3(c)]。29 ℃實驗組2中PG最高,實驗組3次之,對照組最低,兩兩之間存在顯著差異(p<0.05);31 ℃實驗組4顯著高于實驗組5和實驗組6(p<0.05)。雖然升溫對珊瑚幼蟲PG/RD無顯著影響(p>0.05),但實驗組2和4的PG/RD均大于2,且分別顯著高于實驗組3和6(p<0.05),表明珊瑚幼蟲共生體固定的有機碳在硝酸鹽濃度為5 μmol/dm3時凈累積。
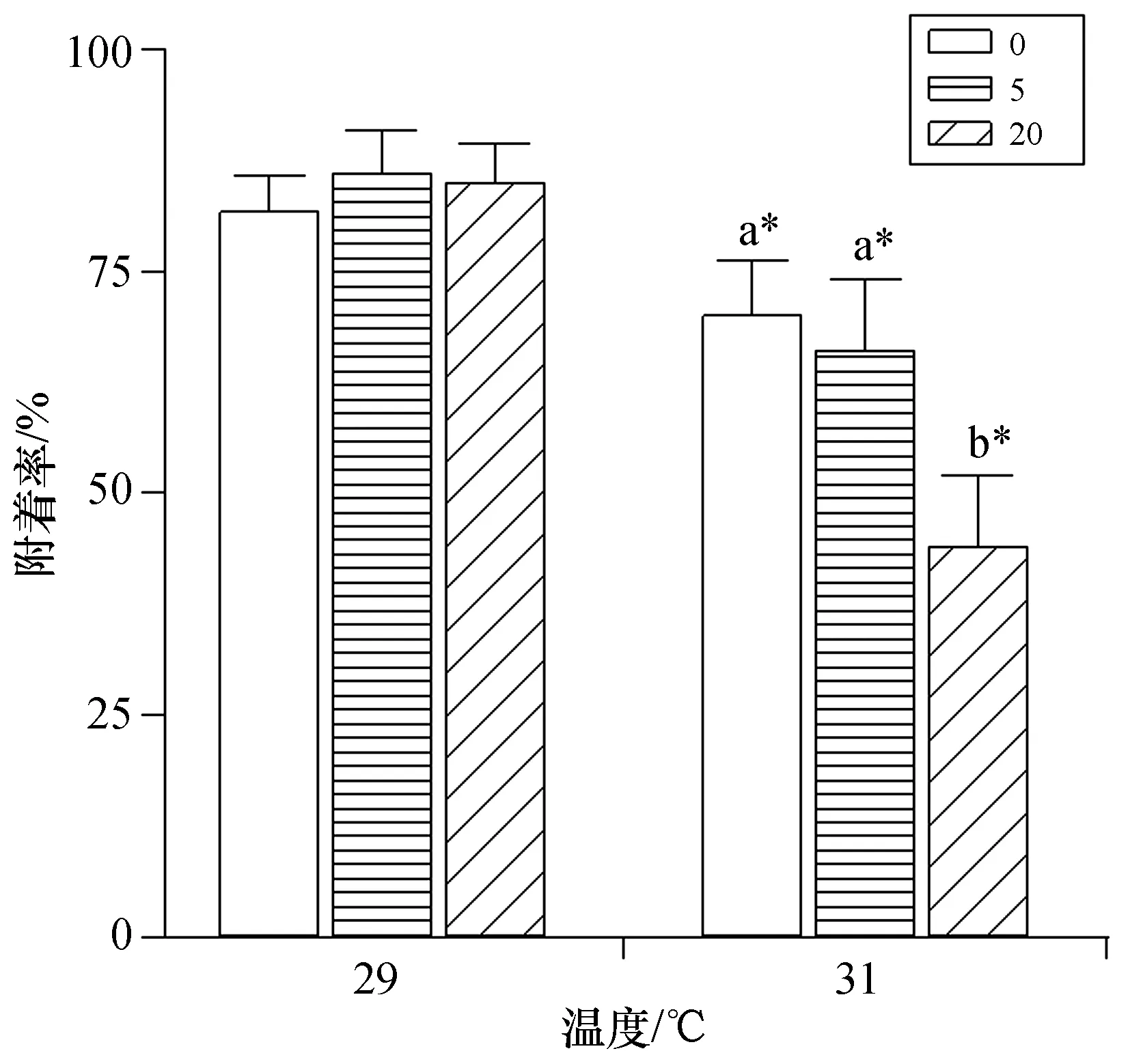
圖2 鹿角杯形珊瑚幼蟲附著率Fig.2 Settlement rates of Pocillopora damicornis larvae by the treatments 圖例數值0、5、20分別表示相應的硝酸鹽濃度,單位為μmol/dm3;不同的小寫字母表示相同溫度下,不同硝酸鹽 濃度處理組之間具有顯著性差異(p<0.05),“*”表示相同硝酸鹽濃度下,不同溫度處理組之間具有顯著性差異(p<0.05)。
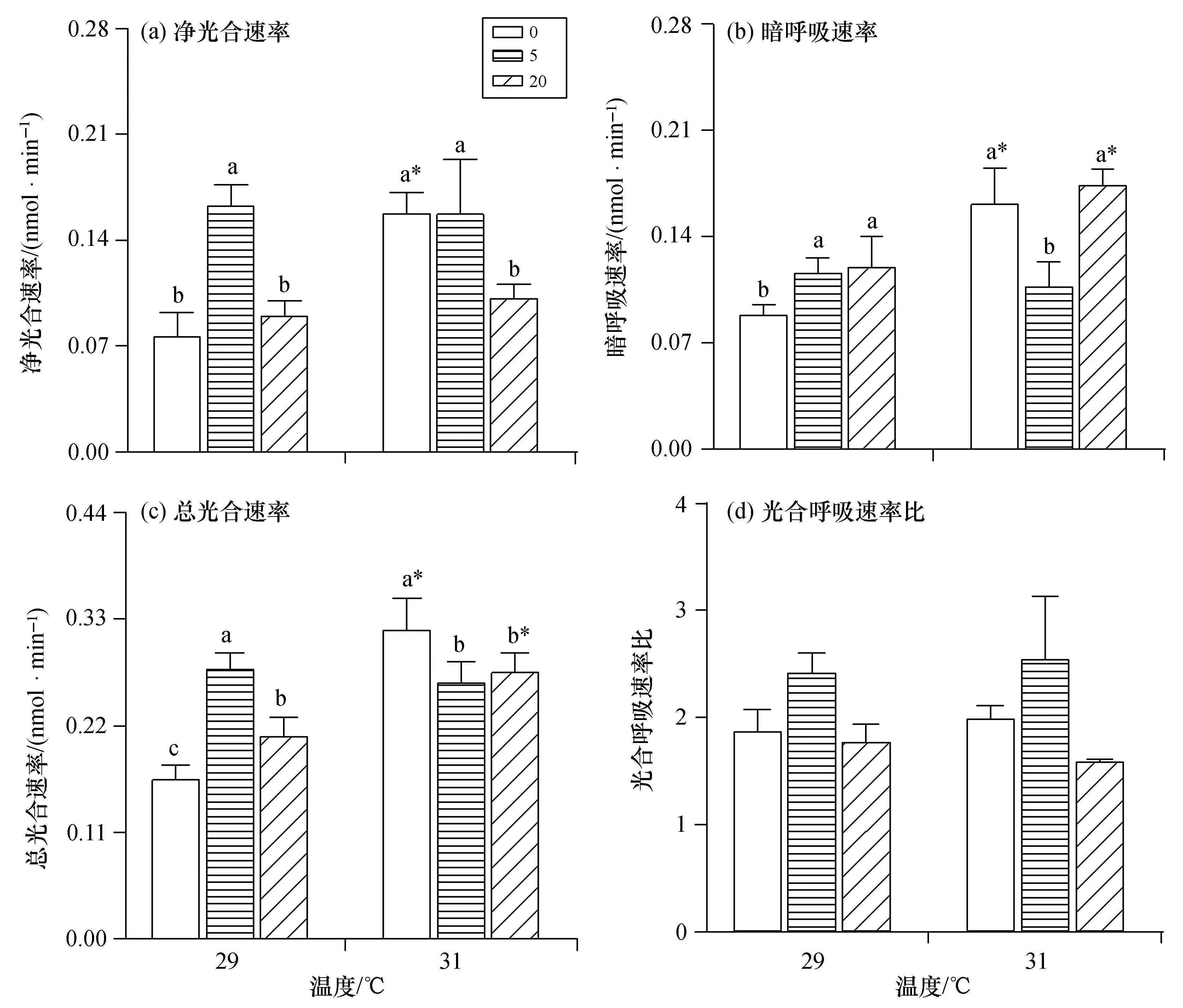
圖3 鹿角杯形珊瑚幼蟲的凈光合速率、暗呼吸速率、總光合速率及光合呼吸速率比Fig.3 Net photosynthesis,dark respiration,gross photosynthesis and photosynthesis ratios to the respiration of Pocillopora damicornis larvae by the treatments 圖例數值0、5、20分別表示相應的硝酸鹽濃度,單位為μmol/dm3;圖(a)、(b)、(c)的縱坐標單位表示每個幼蟲每 分鐘產生或消耗氧氣的物質的量;不同的小寫字母表示相同溫度下,不同硝酸鹽濃度處理組之間具有顯著差異 (p<0.05),“*”表示相同硝酸鹽濃度下,不同溫度處理組之間具有顯著差異(p<0.05)。
2.4 討論
2.4.1 鹿角杯形珊瑚幼蟲存活率和共生蟲黃藻熒光指數 目前,僅有Serrano等(2018)報道了升溫與營養鹽加富共同作用對孵幼型珊瑚幼蟲存活率的影響,結果顯示升溫顯著降低了濱珊瑚(Poritesastreoides)的存活率,但硝酸鹽濃度對幼蟲存活率無影響[23]。同時,Humanes 等(2016)在升溫與營養鹽加富共同作用對排卵型珊瑚幼蟲存活率影響的研究中也顯示柔枝鹿角珊瑚(Acroporatenuis)的存活率在32 ℃顯著降低,但在有機碳加富組無顯著變化[22]。而本研究中,升溫(31 ℃)與硝酸鹽加富對鹿角杯形珊瑚幼蟲的存活率均無顯著影響。實驗結果不一致的原因可能是不同珊瑚種群所處的溫度和營養鹽環境背景差異較大,因此珊瑚幼蟲對升溫和營養鹽加富的適應能力不同。此外,Serrano等在實驗中所用的營養鹽為硝酸鹽[23],而Humanes 等實驗中的營養加富物質包括有機成分與無機成分,組成復雜[22]。實驗中所用的營養鹽成分不同也是珊瑚幼蟲對環境脅迫產生不同響應的原因之一。在升溫對珊瑚幼蟲存活率的影響研究中,結果顯示升溫會對多孔鹿角珊瑚(A.millepora)的存活產生負面影響,但對日本刺星珊瑚(Cyphastreajaponica)、角蜂巢珊瑚(Favitesstylifera)、壯實鹿角珊瑚(A.robusta)和鹿角杯形珊瑚(P.damicornis)的幼蟲存活并沒有顯著影響[20,28-29],而Randall等(2009)對蜂巢珊瑚的研究則表明升溫會顯著降低幼蟲存活率[18]。由此可見,珊瑚幼蟲對溫度的響應確實存在一定的種屬特異性。營養鹽加富對珊瑚幼蟲存活率影響的研究不多,本研究實驗結果與Cox等(2002)[30]、Serrano等[23]和Lam等(2015)[24]的研究結果一致,即營養鹽加富對珊瑚幼蟲存活率無影響,這可能是因為珊瑚幼蟲對營養鹽加富具有一定的耐受性。
共生蟲黃藻熒光指數即Fv/Fm是光系統Ⅱ(PSⅡ)的最大光化學量子產量,反映出PSⅡ反應中心捕獲激發能的效率,若Fv/Fm降低則表示PSⅡ潛在活性中心受損,發生光抑制現象[31]。本研究中,硝酸鹽加富與溫度對珊瑚幼蟲的共生蟲黃藻熒光指數并無顯著影響,說明升溫和硝酸鹽加富不會對共生蟲黃藻的光合系統造成損傷。但是,先前的研究表明最大光化學量子產量(Fv/Fm)對環境脅迫的響應相對滯后[29],并不能良好地預測與評價珊瑚共生體的健康狀況[22]。
2.4.2 鹿角杯形珊瑚幼蟲附著率 本研究首次發現硝酸鹽濃度的增加會促進高溫對鹿角杯形珊瑚幼蟲附著率的負面影響,與Humanes等和Serrano等的研究結果均有所不同[22-23]。Humanes等人發現溫度是促進柔枝鹿角珊瑚幼蟲附著的關鍵因素,而營養鹽加富對幼蟲附著并無影響[22];但Serrano等則認為營養鹽加富是促進濱珊瑚幼蟲附著的主要因子,而溫度對幼蟲附著并無影響[23]。研究結果不同的原因包括兩點,一是因為濱珊瑚在1—9月都可以排放珊瑚幼蟲,使幼蟲可以耐受相對較寬的溫度范圍,對溫度的耐受能力較強;二是因為濱珊瑚幼蟲體內含有母體垂直傳遞而來的蟲黃藻,對營養鹽加富響應相對敏感[22]。與Humanes等觀點一致,我們的結果也表明溫度是影響珊瑚幼蟲附著的主要因素,但其影響表現為顯著降低,這與先前的研究結果吻合[18,32]。珊瑚幼蟲在附著時會發生形態學與生理學兩方面的改變,需要消耗大量的能量[33]。同時,附著與基因表達、細胞分化與重排等細胞生物學過程相關,對高溫脅迫更加敏感脆弱[29]。此外,珊瑚幼蟲附著與殼狀珊瑚藻密切相關[34]。珊瑚幼蟲附著率在高溫條件下顯著降低也可能是因為升溫降低了殼狀珊瑚藻釋放誘導珊瑚附著的化學信號的能力與活力,從而影響珊瑚幼蟲附著行為[18,35]。本實驗中,高濃度硝酸鹽加劇了高溫對珊瑚幼蟲附著率的抑制作用,可能是因為20 μmol/dm3的硝酸鹽對珊瑚幼蟲產生毒害作用,幼蟲為適應高強度的環境脅迫消耗了部分能量,進而導致用于附著的能量比例減少,使珊瑚幼蟲附著率顯著降低。附著率的降低意味著珊瑚幼蟲的浮游期增加,幼蟲可能會隨水流與海浪擴散到與母體珊瑚距離較遠的珊瑚礁區,有利于珊瑚礁之間的基因交流,但對本地珊瑚礁的種群補充可能產生不利的影響[36]。總體來看,高溫與營養鹽加富對鹿角杯形珊瑚的本地種群補充具有負面影響。
2.4.3 鹿角杯形珊瑚幼蟲代謝速率 研究表明,釋放日期、浮游時間和物種間差異均會影響珊瑚幼蟲呼吸速率對環境脅迫的響應[14,37-39]。Cumbo等(2013)發現鹿角杯形珊瑚幼蟲呼吸速率在1~5 d的升溫脅迫會顯著提高21%[37],同樣Rivest等(2015)也發現6 h升溫會顯著提高鹿角杯形珊瑚幼蟲的呼吸速率[40],與本實驗的升溫促進珊瑚幼蟲呼吸速率的結果相同,但Putnam等(2013)則發現9 d的升溫會顯著降低鹿角杯形珊瑚幼蟲的呼吸速率[41]。以上結果表明,同一種珊瑚幼蟲的代謝作用對高溫的響應與脅迫時間相關。Edmunds等(2011)認為,雖然珊瑚幼蟲在適應高溫脅迫時會加速能量消耗,但是代謝速率的改變可以滿足珊瑚幼蟲在高溫環境下的能量需求[42],而代謝的提升可能和升溫下甘油三酯的分解功能有關[40]。除了幼蟲自身的能量物質以外,共生蟲黃藻也可以通過光合作用為珊瑚宿主代謝提供能量[43]。目前,升溫或營養鹽加富對孵幼型珊瑚幼蟲代謝速率影響的相關研究多數聚焦于呼吸作用,忽視了蟲黃藻光合作用在幼蟲代謝中的作用。Ceh等(2013)的研究結果顯示珊瑚共生體可以快速吸收無機氮[44],且共生藻可以固定比其珊瑚宿主細胞多14~23倍的氮[45]。Serrano等分別對排卵型與孵幼型珊瑚幼蟲的呼吸速率進行測量,結果表明硝酸鹽加富對排卵型珊瑚幼蟲的呼吸速率無影響,但卻顯著促進了孵幼型珊瑚幼蟲的呼吸速率[23],作者猜測可能是因為孵幼型珊瑚內蟲黃藻的代謝速率較高所致。本研究中,常溫條件下珊瑚幼蟲的Pnet和PG/RD在5 μmol/dm3硝酸鹽加富組顯著增加且大于2,表明共生蟲黃藻通過光合作用為宿主提供的有機碳足以維系珊瑚幼蟲共生體的呼吸作用與其他生命活動,有機碳有所累積[27]。這可能是因為適量的硝酸鹽加富促進了共生蟲黃藻的生長,提高了共生蟲黃藻的固碳能力,從而為珊瑚幼蟲宿主提供更多的有機碳源,以補償因環境脅迫而消耗的能量,緩解了高溫對珊瑚幼蟲呼吸作用的負面影響。Zhou等(2017)在對鹿角杯形珊瑚成體的研究中也發現,營養鹽增加可以減弱高溫脅迫對珊瑚的負面影響[12]。一方面可能是因為營養鹽促進了珊瑚幼蟲共生蟲黃藻的分裂進而在一定程度上抵消了高溫脅迫下共生蟲黃藻的損失,而適宜的蟲黃藻密度可以為珊瑚宿主提供足夠的有機營養和氧氣,在一定程度上抑制珊瑚的熱脅迫反應[46];另一方面是因為營養鹽濃度的升高可以抑制珊瑚細胞在高溫脅迫時的凋亡與死亡[12]。總體上來看,升溫促進了鹿角杯形珊瑚幼蟲的代謝速率,而5 μmol/dm3的硝酸鹽加富促進了珊瑚幼蟲的有機碳凈累積,減緩了升溫對珊瑚幼蟲呼吸速率的促進作用。
3 結論
本研究以鹿角杯形珊瑚幼蟲為研究對象,探究了升溫與硝酸鹽加富對造礁石珊瑚早期生活史的生理影響,初步得到以下結論:升溫和硝酸鹽加富對鹿角杯形珊瑚幼蟲的存活率與共生蟲黃藻熒光指數無顯著影響,說明升溫與硝酸鹽加富不會對共生蟲黃藻的光合系統造成損傷,珊瑚幼蟲具有良好的存活能力。但是,升溫與硝酸鹽加富對珊瑚幼蟲附著率的影響表現為明顯的協同作用,即高濃度硝酸鹽加富促進了升溫對珊瑚幼蟲附著率的負面影響,可見鹿角杯形珊瑚的本地擴充在升溫與營養鹽加富條件下有所降低。同時,鹿角杯形珊瑚幼蟲的呼吸速率在升溫條件下顯著上升,但5 μmol/dm3卻不受高溫影響,且珊瑚幼蟲共生體有機碳固定在5 μmol/dm3硝酸鹽加富組有所升高,表明適量的硝酸鹽加富可以緩解升溫對鹿角杯形珊瑚幼蟲代謝的不利影響。
