雨生紅球藻HaeDGAT2-3 的分子克隆和生物信息學分析
2020-05-19 03:19:30張宏江趙春超許文鑫崔紅利李潤植
山西農(nóng)業(yè)科學 2020年5期
關鍵詞:分析
張宏江,趙春超,許文鑫,杭 偉,崔紅利,李潤植
(山西農(nóng)業(yè)大學分子農(nóng)業(yè)與生物能源研究所,山西太谷030801)
化石能源的過度開采以及煤炭、石油等的大量使用是造成化石能源枯竭、全球氣候變暖的主要原因。溫室效應的“罪魁禍首”CO2是植物光合作用所必需的底物材料,光合作用過程中,綠色植物可以在光能作用下,將所吸收的CO2和空氣中的水分轉化成碳水化合物并且釋放出O2。微藻是一種誕生于億萬年前并且普遍存在于地球上的光合自養(yǎng)綠色植物[1],它可以利用光合作用高效吸收CO2,并將其轉化為對人類有益的高附加值活性成分,如高蛋白、多糖、蝦青素和生物柴油等[2-4]。近年來,微藻由于其具有光合效率高、繁殖速度快、生長周期短等優(yōu)點,被認為是解決能源危機和治理環(huán)境污染問題的最佳選擇。
三酰甘油(TAGs)是真核生物能量儲存的主要形式,同時也是生物柴油原材料的理想來源[5]。植物體內(nèi)的TAGs 合成主要有2 種途徑:一種是乙酰輔酶A(acyl-CoA)依賴型,也稱肯尼迪途徑;另一種是不依賴于乙酰輔酶A 的代謝途徑[6]。其中,肯尼迪途徑中TAGs 的合成主要是在3 種酰基轉移酶的作用下將底物acyl-CoA 順序酰基化到甘油分子(Glycerol-3-phosphate)sn-1、sn-2、sn-3 位點的過程[7]。這3 種酰基轉移酶依次為甘油三磷酸酰基轉移酶(Glycerol-3-phosphateacyltransferase,GPAT)、溶血磷脂酸酰基轉移酶(Lyso-phosphatidic acidacyltransferase,LPAT)和二酰甘油酰基轉移酶(Diacylglycerol acyltransferase,DGAT)[8]。其中,DGAT 將酰基CoA 上的酰基部分轉移到二酰基甘油(DAG)的sn-3 位置,從而完成TAGs 的最終合成,是TAGs 合成過程中的唯一關鍵限速酶[9-10]。目前,根據(jù)DGAT功能域、亞細胞定位等差異將其分為4 種類型:DGAT1、DGAT2、DGAT3 和WS/DGAT[11-12]。其中,DGAT1 和DGAT2 是結合在內(nèi)質網(wǎng)上并編碼內(nèi)嵌于膜酯雙分子層的微粒體酶,二者具有不同的拓撲結構;而DGAG3 和WS/DGAT 是最近剛發(fā)現(xiàn)的具有DGAT 活性的酶,DGAT3 是一種可溶性酶,WS/DGAT 除了具有DGAT 活性外還具有催化臘酯(Wax ester)合成的活性[12-13]。
雨生紅球藻(Haematococcus pluvialis)是一種單細胞真核綠藻,在脅迫條件下(如高溫、高光和N脅迫等),雨生紅球藻細胞形態(tài)由快速生長的綠色營養(yǎng)細胞轉變?yōu)榧t色厚壁孢子狀態(tài)[14-15],此時蝦青素在藻細胞內(nèi)大量積累以抵御不良環(huán)境。天然蝦青素具有極強的抗氧化性,抗氧化能力是維生素C 的65 倍,是β- 胡蘿卜素的54 倍[16]。相較于其他產(chǎn)蝦青素來源(紅法夫酵母、南極磷蝦等),雨生紅球藻具有生長速度快、蝦青素產(chǎn)量高(達細胞干質量的4%)等優(yōu)點,是公認的天然蝦青素的理想來源[17];此外,其還可以有效清除氧自由基[17],具有延緩衰老的功效,目前已被廣泛應用于化妝品、食品等行業(yè)。
雨生紅球藻中蝦青素主要以蝦青素單酯和蝦青素雙酯的形式存在,少量以游離形式存在。有研究表明,雨生紅球藻受到外界脅迫時,油脂含量會隨之增加[18]。超微結構顯示,雨生紅球藻產(chǎn)生的蝦青素酯在蝦青素積累期儲存于富含TAGs 的油體中[18]。有研究表明[19],DGAT 可能是催化蝦青素酯化的關鍵酶。此外,萊茵衣藻、三角褐指藻和小球藻來源的DGAT 都具有恢復DGAT 缺陷型酵母H1246菌株合成三酰甘油能力的活性[20-21]。然而,雨生紅球藻中的DGAT 目前尚未見報道。
本研究以同源克隆的方法獲得雨生紅球藻中DGAT 基因,并利用生物信息學手段對其理化性質、高級結構和系統(tǒng)進化關系進行分析,以期為雨生紅球藻蝦青素酯化問題研究提供理論依據(jù)。
1 材料和方法
1.1 試驗材料
1.1.1 試驗藻種及培養(yǎng)條件 試驗所用藻種為雨生紅球藻Flotow 1844(購自Dunstaffnage Marine Laboratory),現(xiàn)保存于山西農(nóng)業(yè)大學分子農(nóng)業(yè)與生物能源研究所。所用藻種培養(yǎng)基為BG11 培養(yǎng)基,藻種培養(yǎng)于人工氣候培養(yǎng)箱,培養(yǎng)條件設置為溫度(23±1)℃、光/ 暗周期12 h/12 h、白天光照強度1 400 lx,并且每8 h 搖晃一次。
1.1.2 試驗設備 研究所用儀器為:高壓蒸汽滅菌鍋Panasonic MLS-3781L(日本松下電器產(chǎn)業(yè)株氏會社);超凈工作臺Boxun BJ-CD(上海博迅實業(yè)有限公司);人工氣候培養(yǎng)箱BIC-400(上海博訊實業(yè)有限公司);電泳儀JY-SPCT(北京君意東方電泳設備有限公司);凝膠成像儀、PCR 儀BIO-RAD(伯樂生命醫(yī)學產(chǎn)品(上海)有限公司);分光光度計Nano Drop 2000(美國賽默飛世爾科技公司)。
1.2 試驗方法
1.2.1 總RNA 的提取及cDNA 的獲得 雨生紅球藻總RNA 通過TRizol 法提取[22],并根據(jù)所提取的雨生紅球藻RNA 的濃度利用反轉錄試劑盒(Prime-ScriptTMRT)獲得cDNA 模板。為避免實驗過程中樣品污染,操作人員應佩戴口罩及頭套,所用離心管均為提取RNA 專用離心管。另外,將所提取的RNA保存于-80 ℃冰箱。
1.2.2 同源克隆及RACE 擴增 對來自萊茵衣藻CC3491、小球藻ATCC 30412 以及三角褐指藻的DGAT2 進行序列比對,從而獲得高度保守序列;在此基礎上,利用CODEHOP 軟件進行設計雨生紅球藻DGAT2 的2 段同源克隆引物(F1、R1 和F2、R2)。以獲得的雨生紅球藻cDNA 為模板,按照TaKaRa LA TaqR擴增體系進行PCR 擴增,從而獲得雨生紅球藻HaeDGAT2-3 的同源克隆片段。RACE cDNA 模板的制備按照試劑盒(SMARTerTMRACE cDNA Amplication Kit)說明書進行。在同源克隆片段的基礎上獲得5′端和3′端引物(5′RACE R3、R4 和3′RACE F3、F4)。以RACE cDNA 為PCR擴增模板,以5′端和3′端引物為PCR 擴增引物獲得目的基因兩端(5′端和3′端)的片段。通過DNAStar SeqMan 軟件對中間以及5′端和3′端RACE 片段進行拼接獲得雨生紅球藻HaeDGAT 基因cDNA 序列全長,并據(jù)此設計表達框引物(F5、R5)。所用引物信息如表1 所示,均由上海生工生物工程有限公司合成。

表1 引物信息
1.3 生物信息學分析
雨生紅球藻HaeDGAT2-3 氨基酸序列是通過序列處理在線工具包(SMS)(http://www.bio-soft.net/sms/),將獲得的HaeDGAT2-3 的核苷酸翻譯而來,開放閱讀框的查找是通過在線ORFfinder 軟件(https://www.ncbi.nlm.nih.gov/orffinder/)對目的基因進行;通過在線軟件ProtParam(http ://web.expasy.org/protparam/)對雨生紅球藻HaeDGAT2-3 的理化性質進行分析;跨膜域及疏水區(qū)的預測是通過在線軟件TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)及ProtScale(http ://web.expasy.org/protscale/)進行;雨生紅球藻HaeDGAT2-3 及其他物種來源的DGAT2 蛋白保守功能域通過NCBI 的CCD 在線軟件(https://www.ncbi.nlm.nih.gov/cdd/)進行預測;二級結構及三級結構的預測分別通過SOPMA(http://metadatabase.org/wiki/SOPMA) 和 SWISS-MODEL(http://swissmodel.expasy.org/)進行;多序列比對是通過本地軟件BioEdit 進行分析;系統(tǒng)進化樹是通過Clustal W 軟件進行序列比對,再通過本地軟件MEGA 7.0 進行繪制,最終通過在線軟件EvolView(https://www.evolgenius.info/)進行修飾。
2 結果與分析
2.1 總RNA 的提取和基因克隆結果分析
由圖1 可知,泳道1 從上往下依次是28SrRNA、18S rRNA 和5S rRNA 等3 個條帶,其中,28S rRNA、18S rRNA 條帶帶型清晰,5S rRNA 條帶較模糊,說明所提RNA 質量較好;NanoDorp 2000 測定總RNA濃度是850 ng/μL,A260/A280及A260/A230值分別為1.8~2.0 和1.9~2.0,說明所提RNA 質量較好,可進行后續(xù)試驗;泳道2 和3 大小分別為482、773 bp;泳道4和5 大小分別為667、1 149 bp。
2.2 雨生紅球藻HaeDGAT2-3 蛋白cDNA 序列分析
雨生紅球藻2 型二酰甘油酰基轉移酶DGAT2(HaeDGAT2-3)的cDNA 序列全長是在同源克隆的基礎上利用RACE (rapid-amplification of cDNA ends)技術獲得,NCBI 注冊號為MN073496。如圖2所示,HaeDGAT2-3 序列全長為1 446 bp,編碼區(qū)從248~1 396 bp 共1 149 bp,編碼383 個氨基酸,其中,5′端非編碼區(qū)(5′-UTR)和3′端非編碼區(qū)(3′-UTR) 長度分別為247、49 bp。利用NCBI 中的BLASTp 程序對HaeDGAT2-3 氨基酸序列進行比對分析,結果發(fā)現(xiàn),HaeDGAT2-3 基因與衣藻來源的DGAT2 基因(NCBI 檢索號為GAX79066.1)相似性最大(54.02%),與小球藻、月牙藻來源的DGAT2 基因(NCBI 檢索號分別為QBG05555.1、GBF91693.1)相似性分別達45.08%和41.82%。
2.3 雨生紅球藻HaeDGAT2-3 蛋白理化性質分析
雨生紅球藻HaeDGAT2-3 蛋白基本理化性質通過在線軟件ExPASY 中的ProtParam 工具進行分析,結果顯示,該蛋白分子式為C1844H2824N486O489S17,分子質量為40.17 ku,理論等電點為9.36,屬于堿性蛋白。編碼HaeDGAT2-3 蛋白的20 種氨基酸中丙氨酸(Ala)含量最高(13.3%);其次為亮氨酸(Leu)和絲氨酸(Ser),含量分別為10.6%和7.6%;而色氨酸(Trp)和半胱氨酸(Cys)含量偏低,均為1.6%。說明該蛋白屬于不穩(wěn)定蛋白。在線分析軟件ProtScale 分析結果顯示,HaeDGAT2-3 蛋白存在至少3 個疏水區(qū),屬于疏水蛋白。
2.4 雨生紅球藻HaeDGAT2-3 蛋白的高級結構分析
雨生紅球藻HaeDGAT2-3 蛋白的二級結構通過在線軟件SOPMA 進行分析,結果顯示(圖3-A),目的蛋白中無規(guī)則卷曲所占比例最高(44.87%),其次為α- 螺旋(32.88%),延伸鏈和β- 折疊分別占18.75%和5.71%。圖3-B 為雨生紅球藻DGAT2 的三級結構預測結果,圖3-C 為三級結構所參照模型。
雨生紅球藻HaeDGAT2-3 與其他不同來源DGAT2 蛋白的保守域通過NCBI 的在線工具進行預測,結果顯示(圖4-A),同其他來源的DGAT2 相同,HaeDGAT2-3 基因也有DGAT 基因的典型LPLAT 家族,暗示HaeDGAT2-3 具有與其他來源DGAT2 蛋白相似的功能;利用在線分析軟件TMHMMServer 對雨生紅球藻HaeDGAT2-3 與其他不同來源DGAT2 蛋白的跨膜域進行預測,結果顯示(圖4-B),HaeDGAT2-3 蛋白在40—90 位氨基酸區(qū)間內(nèi)有2 個明顯的跨膜域;對其他來源DGAT2 蛋白的跨膜域進行分析表明,DGAT2 蛋白有1~3 個跨膜域。此外,信號肽分析結果顯示, 雨生紅球藻HaeDGAT2-3 中無信號肽存在。圖4 中DGAT2 序列相關信息如表2 所示。
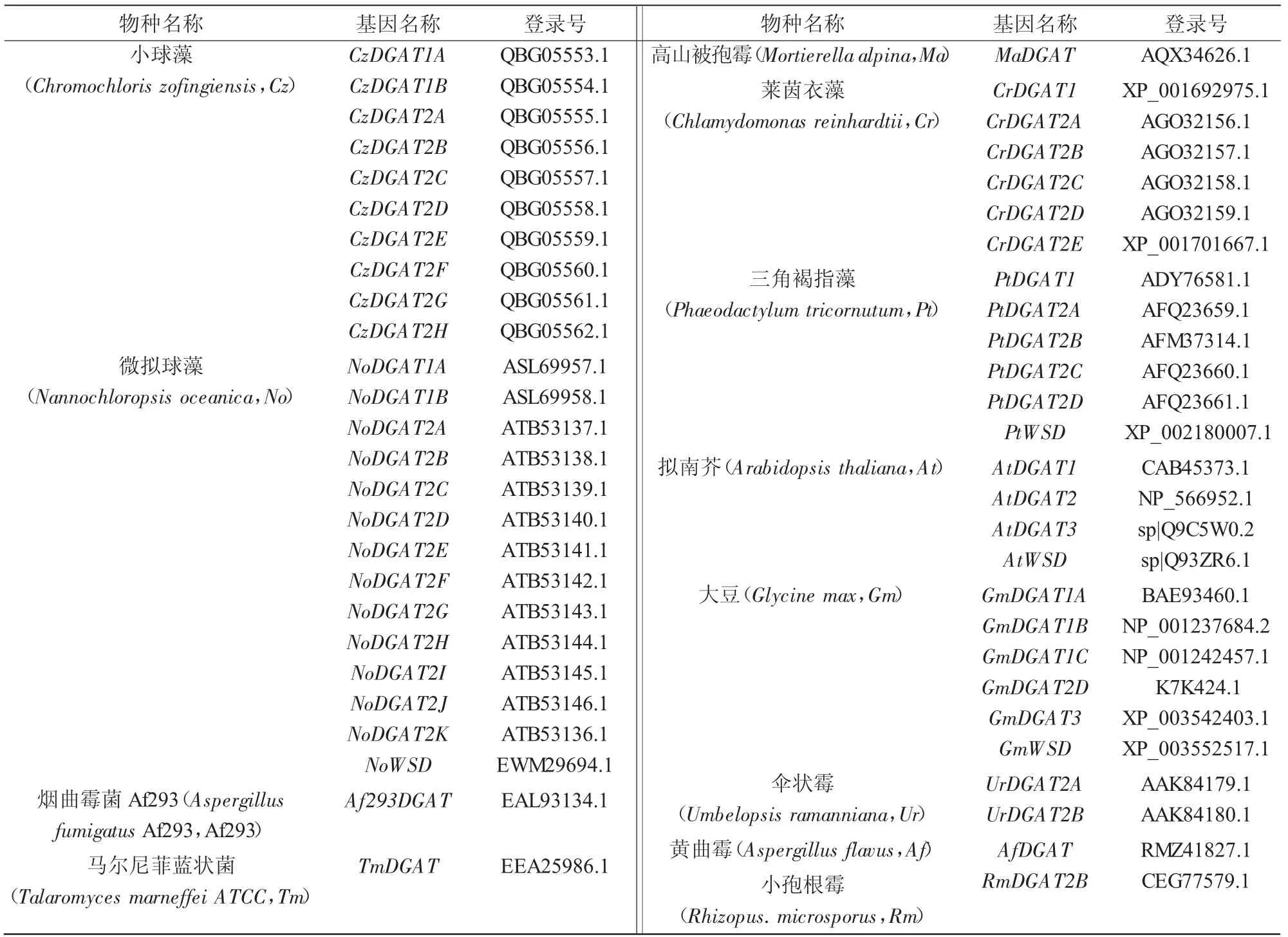
表2 DGAT 基因名稱及其登錄號
2.5 雨生紅球藻HaeDGAT2-3 多序列比對分析
通過本地軟件BioEdit 對HaeDGAT2-3 氨基酸序列和其他來源的DGAT2 進行比對分析,結果顯示(圖5),HaeDGAT2-3 氨基酸序列中共包含7 個保守的功能基序,分別為YFP、PH、PR、GGE、RGFA、VPFG 和G 基序。在YFP 基序中,所有序列前2 個氨基酸皆為YF,而只有HaeDGAT2-3、AtDGAT2、CzDGAT2D 等7 個序列在第3 個氨基酸位置上為苯丙氨酸(Phe);在PH 基序中,只有CzDGAT2D、CrDGAT2D、AtDGAT2 和GmDGAT2 等4 個序列第3 個氨基酸位置上的甘氨酸(Gly,G)被絲氨酸(Ser,S)所取代,暗示這4 個氨基酸序列在功能上相似;在PR 基序中,雨生紅球藻來源的HaeDGAT2-3 氨基酸序列同高等植物來源的DGAT(AtDGAT2 和GmDGAT2)在第1 個位置上的氨基酸保持一致,第2 個位置上的氨基酸在所測試序列中散亂分布,只有氨基酸序列相對特殊的微擬球藻來源的NoDGAT2K 在第4 個位置上的氨基酸是甘氨酸(Gly,G),其余全都是精氨酸(Arg,R);在GGE 基序中,所有序列都含有完整的GGE 基序,但是在2 個位置上的氨基酸并不保守,而只有CzDGAT2D、CrDGAT2D、AtDGAT2 和GmDGAT2 這4 個氨基酸序列在這2 個位置上的氨基酸保持一致,再一次暗示其功能相似;在基序RGFA 中,雨生紅球藻HaeDGAT2-3 異亮氨酸(Ile,I)替換了苯丙氨酸(Phe,F(xiàn)),這可能是造成功能差異的關鍵原因;VPFG 基序相比其他基序有很大的差異性,相對不保守;在基序G 中,所有氨基酸序列都有完整的G 基序,相對保守。從圖5 的分析結果可以看出,HaeDGAT2- 3 與CrDGAT2A、CzDGAT2A和CzDGAT2B可能具有相同的功能。
2.6 雨生紅球藻HaeDGAT2-3 的系統(tǒng)進化分析
為進一步研究雨生紅球藻來源的HaeDGAT2-3與其他來源DGAT 的進化關系,從NCBI 數(shù)據(jù)庫以及已發(fā)表的文獻中搜集到53 條DGAT 序列,包括微藻、菌類和高等植物來源的DGAT1s、DGAT2s、DGAT3s 和WS/DGATs 等4 種不同類型的二酰甘油酰基轉移酶,在序列比對的基礎上對其進行系統(tǒng)進化分析,結果顯示(圖6),4 種不同類型的DGATs分別聚為一支,而且微藻、菌類和高等植物來源的DGAT 在各自的類群中明顯分開;值得注意的是,HaeDGAT2-3 與CrDGAT2A、CzDGAT2A和CzDGAT 2B聚為一個小支,暗示它們具有相同的功能,進一步驗證了多序列比對的預測結果。圖6 進化樹所有序列的相關信息列于表2。
3 結論與討論
雨生紅球藻是一種單細胞綠色微藻,被認為是天然蝦青素的理想來源[5],并且其產(chǎn)生的蝦青素以蝦青素單酯和蝦青素雙酯的形式存儲于富三酰甘油(TAGs)的油體中。二酰甘油酰基轉移酶(DGAT)是依賴于乙酰輔酶A(acyl-CoA)合成TAG 的肯尼迪途徑中催化TAG 生物合成最后一步的關鍵限速酶[9-10],目前,已有大量來源于不同物種(包括高等植物、微藻和真菌)的DGAT 被用來探究DGAT 的表達與脂肪酸含量及成分的變化關系,研究發(fā)現(xiàn),高等植物中續(xù)隨子來源的EiDGAT2 基因可以提高煙草葉片中的總脂含量[23],紫蘇來源的PfDGAT1 被發(fā)現(xiàn)可以恢復TAG 缺陷型酵母H1246 重新合成油脂的能力[24];高山被包霉來源的MaDGAT 基因以及毛霉菌來源的McDGAT 基因被證實具有恢復H1246合成油脂的能力,但它們有著不同的底物偏好性[25-26];此外,萊茵衣藻、微擬球藻、三角褐指藻等微藻來源的DGAT 也相繼被發(fā)現(xiàn)具有催化TAG 合成的能力[5,21,27]。然而,雨生紅球藻來源的DGAT2 并未見報道。
本研究通過同源克隆方法獲得了雨生紅球藻DGAT2 的同源片段及同源克隆引物,并在此基礎上通過RACE 擴增技術獲得雨生紅球藻HaeDGAT2-3的cDNA 序列全長1 446 bp,編碼383 個氨基酸,其中,丙氨酸(Ala)含量最高,為13.3%。此外,雨生紅球藻HaeDGAT2-3 蛋白分子質量為40.17 ku,理論等電點為9.36。二級結構分析結果顯示,無規(guī)則卷曲所占比例最高,為44.87%。根據(jù)NCBI中的BLASTp分析結果顯示,雨生紅球藻來源的DGAT2 與萊茵衣藻、小球藻和月牙藻來源的DGAT2 相似性均在40%以上,分別達54.02%、45.08%和41.82%,說明其功能具有相似性。DGAT1s 和DGAT2s 雖然在分子結構上有很大的差異,屬于2 個不同的基因家族,但在功能水平上卻是一致的。DGAT1s 一般具有8~9 個跨膜域,而DGAT2s 中跨膜域相對較少,一般為1~3 個。比如,紫蘇、小球藻來源的DGAT1 都有9 個跨膜螺旋區(qū)[27-28],續(xù)隨子來源的DGAT2 有2 個跨膜區(qū)[24]。本研究發(fā)現(xiàn),HaeDGAT2-3 蛋白具有2 個跨膜區(qū),符合DGAT2 家族序列特點。保守域分析結果顯示,雨生紅球藻來源的DGAT2 具有同其他DGAT2來源相同的LPLAT 家族。
此外,本研究對HaeDGAT2-3 與微藻、高等植物和細菌來源的DGAT 進行多序列比對,結果表明,同其他物種來源的DGAT2 相同,HaeDGAT2-3也具有7 個功能域;在這7 個功能域中,YFP、RGFA和VPFG 基序的保守性較低。同時,CzDGAT2D、CrDGAT2D、AtDGAT2 和GmDGAT2 這4 個氨基酸序列具有很高的相似性。值得注意的是,在第5 個基序RGFA 中,HaeDGAT2-3 蛋白中異亮氨酸(Ile,I)替換了苯丙氨酸(Phe,F(xiàn)),暗示HaeDGAT2-3 基因可能具有不同的功能。系統(tǒng)進化分析結果表明,HaeDGAT2-3 基因確實屬于DGAT2 家族,并且與CrDGAT2A、CzDGAT2A 和CzDGAT2B 聚為同一個小支,暗示其具有相同的功能。
本研究在同源克隆的基礎上利用RACE 擴增技術首次獲得了雨生紅球藻中2 型二酰甘油酰基轉移酶DGAT2 的cDNA 序列全長,并利用生物信息學方法對其進行分析,結果顯示,HaeDGAT2-3 蛋白分子式為C1844H2824N486O489S17,分子質量為40.17 ku,編碼區(qū)全長1 149 bp,共編碼383 個氨基酸。功能域分析表明,其可能具有催化TAG 合成的功能。同其他來源的DGAT2 相同,HaeDGAT2-3 也具有7 個保守的功能域,而在RGFA 基序中,HaeDGAT2-3表現(xiàn)出一定的特異性,暗示其功能可能不盡相同。進化樹分析結果表明,其與已經(jīng)報道的具有TAG合成功能的DGAT2 相似性極高,暗示其可能亦具有合成TAG 的功能。本試驗結果可為進一步了解雨生紅球藻中蝦青素酯化機制提供參考。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
