基于psbA-trnH 序列對藥用黃芪及其混偽品的鑒別
2020-05-19 03:19:28耿雅萍馬艷珠解瀟冬張鵬飛劉亞令
山西農業科學 2020年5期
耿雅萍,馬艷珠,王 芳,解瀟冬,張鵬飛,劉亞令
(1.山西農業大學生命科學學院,山西太谷030801;2.山西農業大學園藝學院,山西太谷030801)
黃芪原名黃耆,屬多年生草本植物,始載于《神農本草經》,其藥用歷史悠久,是傳統中藥體系中的重要藥材[1]。《中華人民共和國藥典》(2015 版第一部)中規定,正品藥用黃芪為豆科黃芪屬植物蒙古黃芪(Astragalus membranaceus(Fisch.)Bge.var.mongholicus(Bge.)Hsiao)或膜莢黃芪(Astragalus membranaceus(Fisch.)Bge.)的干燥根[2]。近年來,隨著藥用黃芪野生量的迅速減少,該種被列為漸危種[3]。目前,市場上多以栽培黃芪為主,但較為混雜,除了正品的藥用黃芪外,還存在著許多混偽品,常見的有紫花苜蓿(Medicago sativa L.)、蜀葵(Althaea rosea(Linn.)Cavan.)、錦雞兒(Caragana sinica(Buc'hoz)Rehder.)、藍花棘豆(Oxytropis coerulea(Pall.)DC.)等。這些植物藥用部位的外形特征與藥用黃芪十分相似,但在臨床應用方面,其藥效相差甚遠,其中部分混偽品甚至還具有一定的毒性,嚴重影響中醫藥的療效與用藥安全[4-5]。因此,急需一種簡便高效的手段對黃芪及其混偽品進行準確鑒別,以確保藥品質量和臨床用藥安全。
傳統的植物鑒定方法是對其表型如株高、葉片大小、數量等進行觀察,但由于形態觀察極易混淆,且極易受到人為主觀因素、實際經驗以及外界環境、季節性的影響,因此,不能準確充分地鑒定出物種間的進化關系[6-7]。而相較于傳統的鑒定方法,DNA條形碼(DNA barcoding)是目前發展較快的能夠簡便有效地鑒定動植物物種的一項分子生物學技術,自2003 年由加拿大分類學家HEBERT 等[8]首次提出后,受到了各國分類學家的廣泛關注。其原理是利用一個或幾個變異程度高、系統進化信息豐富、引物設計區保守、擴增容易、標準規范且相對較短的DNA 片段作為標記對目標物種進行有效鑒定,具有通用性強、操作簡便、鑒定準確、用時少等優點[9-11]。目前,在植物物種鑒定方面,常用的DNA 條形碼為葉綠體序列psbA-trnH、trnL-trnF、matK、rbcL 以及核糖體序列ITS、ITS2 等。這些DNA 條形碼能夠簡便有效地鑒定中藥材基原物種,并為中藥材的溯源、種質資源的評價、保護以及可持續開發利用提供數據支撐和物質基礎。其中,psbA-trnH 序列是位于編碼光合系統Ⅱ反應中心DI 蛋白的psbA 基因和編碼tRNA 組氨酸的trnH 基因之間的一段非編碼序列[12]。葉綠體基因組中psbA-trnH 序列在非編碼區中功能上限制少,顯示的變異位點多,因此,易設計通用引物。同時,與編碼區相比,該片段可以提供更多的系統發育信息[13]。
本試驗以藥用黃芪蒙古黃芪和膜莢黃芪為研究對象,利用psbA-trnH 通用引物對其進行序列分析及對各序列進行差異性分析,觀察其變異位點,探討其遺傳變異程度,并從GenBank 上下載的藥用黃芪混偽品(紫花苜蓿、蜀葵、錦雞兒)的psbA-trnH序列為對照,構建其系統發育樹,研究其對藥用黃芪及其混偽品的鑒定能力,從而為藥用黃芪優良種質資源的選擇、保護、評價以及分類提供一定的科學依據,為黃芪的產業化發展及可持續性開發利用提供物質基礎。
1 材料和方法
1.1 試驗材料
供試材料為來自山西渾源和內蒙古包頭的蒙古黃芪各5 份以及來自山西渾源和吉林新興林場的膜莢黃芪各5 份(表1)。另外從GenBank 數據庫中下載了6 份黃芪混偽品,分別為2 份紫花苜蓿、2 份蜀葵、2 份錦雞兒(表2)。

表1 黃芪樣品信息

表2 混偽品樣品信息
1.2 試驗方法
1.2.1 黃芪樣本的DNA 提取 試驗采用CTAB 法提取黃芪的DNA,所得DNA 用1%的瓊脂糖凝膠電泳進行質量檢測,并用核酸蛋白測定儀進行純度檢測后,放于-20 ℃冰箱中保存備用。
1.2.2 PCR 擴增及測序 本試驗擴增體系為30 μL,其中,1 μL DNA 模板,1 μL psbA 引物(5′-GTTATG CATGAACGTAATGCTC-3′),1 μL trnH 引物(5′-CGCGCATGGTGGATTCACAATCC-3′),15 μL Taq PCR MasterMix(+dye),加ddH2O 至30 μL。PCR 反應擴增程序為:95 ℃預變性5 min;94 ℃變性45 s,55 ℃復性40 s,72 ℃延伸1 min,33 個循環;72 ℃補平1 min,4 ℃保存。利用1%瓊脂糖凝膠電泳法檢測PCR 產物,后將擴增成功的PCR 產物送至上海美吉常規測序公司進行雙向測序。
1.3 數據分析
利用CExpress 序列拼接軟件對測序結果進行人工拼接校對,并去除兩端引物區及低質量區,得到一條完整的psbA-trnH 序列;利用EditSeq 軟件對20 個樣本psbA-trnH 序列的堿基含量及序列特征進行統計分析;通過DNAman 軟件對本試驗所有樣品的20 條psbA-trnH 序列進行差異性比對,并分析其變異位點;通過MEGA 7[14]軟件基于K2-P(Kimura 2-Parameter)距離模型,計算黃芪種內、居群間的遺傳距離,構建Neighbor-Joining(NJ)系統發育樹,并用bootstrap1000 次檢驗各分支置信度,評價分支系統發育關系的可靠性。
2 結果與分析
2.1 PCR 產物檢測
利用1%瓊脂糖凝膠電泳對PCR 擴增產物進行檢測,從圖1 可以看出,擴增的條帶亮且清晰,無其他雜帶,蒙古黃芪與膜莢黃芪樣品psbA-trnH 序列的PCR 產物長度在500 bp 附近,符合測序要求,可用于后續分析。
2.2 黃芪psbA-trnH 序列特征及差異性分析
利用DNAstar 軟件計算黃芪psbA-trnH 序列中的G+C 含量及總長度(表3),結果表明,G+C含量為26.10%~27.88%,序列長度在345~378 bp,其中,M-JX5 的psbA-trnH 序列最短,為345 bp;M-JX1 的psbA-trnH 序列最長,為378 bp。不同居群藥用黃芪psbA-trnH 序列的G+C 含量不同,居群G-SH、G-NB、M-SH、M-JX 的G+C 平均含量分別27.35%、26.76%、26.85%、27.48%。其中,G-NB(26.76%)與M-JX(27.48%)的G+C 含量相差最大,表明這2 個居群之間的親緣關系較遠。對20 份藥用黃芪psbA-trnH 序列進行比對差異分析,結果表明,20 份藥用黃芪樣品中有337 個保守位點,保守率為95.74%;有13 個變異位點,變異率為3.69%(表4)。

表3 黃芪psbA-trnH 序列中G+C 含量及序列長度

表4 黃芪psbA-trnH 序列變異位點
2.3 黃芪psbA-trnH 序列的遺傳距離分析
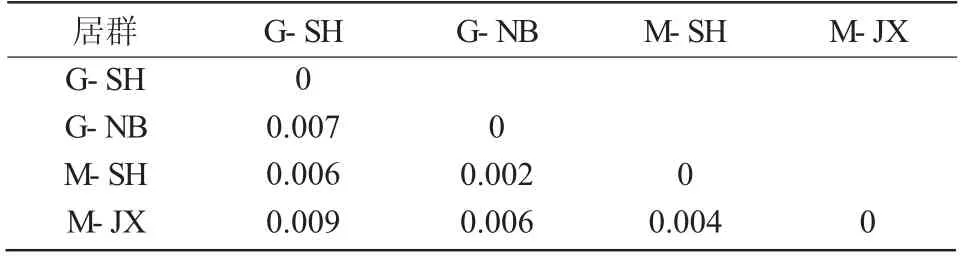
表5 黃芪各居群之間的遺傳距離
利用MEGA 7 軟件計算黃芪各居群之間的遺傳距離和黃芪種內個體間的遺傳距離(表5、圖2)。黃芪各居群之間的遺傳距離為0.002~0.009,其中,居群G-SH 與M-JX 之間遺傳距離最大,為0.009;居群G-NB 與M-SH 之間遺傳距離最小,為0.002。黃芪種內遺傳距離范圍為0.000~0.023,其中,個體M-JX1 與G-SH5 之間遺傳距離最大,為0.023。
2.4 基于psbA-trnH 序列的黃芪及其外類群系統發育樹分析
本試驗以紫花苜蓿、蜀葵、錦雞兒作為外類群,采用鄰接法構建系統發育樹對蒙古黃芪和膜莢黃芪樣品以及外類群進行聚類分析(圖3)。結果顯示,將其分成了三大支,其中,蒙古黃芪與膜莢黃芪為一大支,紫花苜蓿和錦雞兒為一大支,蜀葵單獨為一大支,其自展支持率各達100%。在藥用黃芪的一大支中,內蒙古包頭的蒙古黃芪(G-NB1~5)、山西渾源的8 個樣品(4 個蒙古黃芪與4 個膜莢黃芪)以及遼寧新興林場的膜莢黃芪M-JX2、M-JX3 聚為一支,自展支持率達64%,山西渾源的G-SH5、M-SH1 與吉林新興林場的M-JX1、M-JX5 聚為一支,自展支持率達61%。
3 結論與討論
近年來,DNA 條形碼技術已成為國內外研究人員在生物物種鑒定及遺傳多樣性分析方面的重要手段之一。在藥用植物鑒定方面,psbA-trnH 被作為ITS2 序列的輔助序列,可有效用于藥用植物的鑒定[15]。萬如等[16]通過對21 份材料進行psbA-trnH序列分析發現,枸杞屬植物的航天誘變種與普通栽培種各聚為一支,得出psbA-trnH 可以用于親緣關系較遠的枸杞屬植物研究。韓正洲等[17]通過對蘆根及其混偽品進行psbA-trnH 序列分析發現,蘆根及其混偽品分成了兩大支,同時各分支均有較高的自展率。吳海燕[18]研究表明,利用葉綠體psbA-trnH 基因在序列和結構上相對保守,能夠有效并鑒別甘肅各地的黃芪和紅芪,有利于保障黃芪的用藥安全和臨床療效。徐子欣等[19]通過對不同居群的巴戟天進行psbA-trnH 和rDNA ITS 序列分析,得出psbAtrnH 基因能夠廣泛應用于地理居群較近的物種之間的研究。張宏意等[20]對不同產地不同個體的何首烏進行了psbA-trnH 序列分析,結果顯示,各居群何首烏的親緣關系大體上符合地域差異且其遺傳變異顯著。本試驗利用葉綠體基因組psbA-trnH 序列分析山西渾源的蒙古黃芪和膜莢黃芪、內蒙古包頭的蒙古黃芪、吉林新興林場的膜莢黃芪,以黃芪混偽品(紫花苜蓿、蜀葵、錦雞兒)為對照,利用NJ法構建系統發育樹。結果表明,藥用黃芪及其混偽品之間存在有較大程度的變異。藥用黃芪聚為一大支,紫花苜蓿與錦雞兒聚為一支,蜀葵單獨聚為一支,且自展支持率各達100%,表明利用psbA-trnH序列可以有效鑒別黃芪及其混偽品。此外,根據聚類結果得到藥用黃芪與紫花苜蓿、錦雞兒的親緣關系較近,與蜀葵的親緣關系較遠。此結果與崔占虎等[4]利用ITS 序列對黃芪與其混偽品的鑒定結果一致。
不同物種psbA-trnH 序列的G+C 不同。G+C含量的差異是反映物種親緣關系遠近的遺傳性特征,親緣關系越近,G+C 含量差異較小;親緣關系越遠,G+C 含量差異較大[21]。王麗芝等[22]對8 種川芎種質及藁本、遼藁本之間G+C 含量進行了比較分析,結果表明,藁本、遼藁本與川芎間G+C 含量相近,表明彼此間親緣關系較近。周文才等[23]基于ITS 序列鑒定三葉木通與白木通,結果表明,二者具有相似的G+C 含量,驗證了其很近的親緣關系。本試驗通過計算各居群蒙古黃芪與膜莢黃芪psbAtrnH 序列的G+C 含量,得出來自吉林的膜莢黃芪M-JX(27.48%) 與來自內蒙古包頭的蒙古黃芪G-NB(26.76%)之間的含量差異最大,因此,這2 個居群之間的親緣關系較遠,與其居群間的遺傳距離結果一致,即居群M-JX 與G-NB 的遺傳距離最大,為0.023。
本試驗通過分析藥用黃芪psbA-trnH 序列的G+C 含量、遺傳距離、保守率、變異率等指標,以混偽品為對照構建系統發育樹,表明可以利用psbAtrnH 序列鑒別藥用黃芪及其混偽品。此結果為藥用黃芪及其混偽品的鑒別提供了分子水平依據,為藥用黃芪優良種質資源的保護、評價以及分類提供了一定的數據支撐。
