鹽脅迫環境下植物促生菌的作用機制研究進展
2020-05-13 14:57:02紀超王曉輝劉訓理
生物技術通報 2020年4期
關鍵詞:植物
紀超 王曉輝 劉訓理
(1. 山東農業大學林學院 黃河下游森林培育國家林業和草原局重點實驗室,泰安 271018;2. 山東農業大學生命科學學院,泰安 271018)
土壤鹽漬化是全球土地退化的主要表現形式之一,目前,全世界至少20%的耕地遭受不同程度的鹽漬化威脅[1]。雖然植物能夠在鹽漬土生境中有一定的產量,但是多數作物和樹種耐鹽性不高,其中,小麥、玉米、水稻、大麥等主要糧食作物受到鹽脅迫減產可達到70%[2]。鹽脅迫對植物的生長發育具有多方面的作用,包括復雜的分子、基因、生物化學、生理學和形態學影響[3]。脅迫持續的時間與嚴重性能夠造成植物不同程度的損傷[4-5],如初期的滲透脅迫、氧化脅迫和離子毒性可能導致植物細胞膜結構破壞、營養失衡、活性氧(Reactive oxygen species,ROS)消除能力下降、抗氧化酶活性下降、光合作用減弱等,隨著大量的Na+和Cl-的攝入,細胞離子平衡和生理平衡被打破,植物根系吸水能力降低,葉片中含水率下降[6],植物難以吸收K+,逐漸造成植物營養虧缺、生產力下降甚至死亡[4]。與其他脅迫類似,植物通過調控復雜的信號網絡來應對鹽脅迫,包括脅迫傳感蛋白、信號傳導因子、轉錄因子、應激反應基因和代謝物等[7]。
土壤鹽漬化降低了植物根際土壤水的能量狀態,使土壤溶液滲透壓超過了植物細胞液的正常滲透壓,導致植物根系長度、密度和結構改變,阻礙了植物對土壤水分和養分的攝取,使植物處于生理干旱狀態,生長速度變得緩慢,甚至萎蔫或死亡[8]。滲透脅迫和離子脅迫相互作用產生次級氧化脅迫[9],打破了植物體內活性氧產生與清除間的動態平衡,破壞細胞膜系統的完整性,影響了植物的光合作用[10]。
植物促生菌(Plant growth-promoting bacteria,PGPB)能與植物建立共生關系,并在正常狀態下或植物面臨脅迫時促進其生長[11]。合理使用PGPB 是減輕鹽脅迫危害的一種重要途徑[12]。不具有耐鹽性的PGPB 在自然條件下隨鹽度增加逐漸失去PGP(Plant growth-promoting)特征,開發利用耐鹽PGPB是鹽脅迫下增加作物產量的可行措施。研究發現PGPB 具有產植物激素、鐵載體、ACC 脫氨酶,溶磷,固氮,解鉀,上調抗氧化酶(如SOD、POD 等),調節離子轉運蛋白(如HKT)表達,分泌胞外多糖(EPS),改變根系結構和形態、導水率和激素狀態,釋放與脅迫相關的揮發性化合物,積累滲透物(谷氨酸、脯氨酸、肽等),抑殺病原菌等特性,以上機制與提高植物耐鹽性、誘導植物產生鹽脅迫系統抗性(Induced system tolerance,IST)有關,對促進植物根、莖和葉片的發育及生長、防治病害、調節植物根際微生物群落結構有顯著效果[1,3,13-23]。此外,蛋白質組學分析顯示,PGPB 影響參與植物光合作用、抗氧化過程、跨膜轉運和發病相關的多種蛋白質的表達[24]。本文綜述了PGPB 在鹽脅迫中提高植物耐鹽性的作用機制研究進展,旨在為篩選能夠改良鹽堿地的耐鹽促生菌株奠定理論基礎。
1 PGPB 產生ACC 脫氨酶
乙烯是幾乎所有植物都會產生的植物生長調節劑和應激激素[25],能夠引起植物產生生理、分子水平上的變化。鹽脅迫能夠激發植物產生過量的乙烯,結節中的乙烯水平升高將降低固氮量,從而抑制植物根及植株的進一步生長,降低脅迫誘導的乙烯水平將減輕作物的脅迫損傷。分泌ACC 脫氨酶(ACC deaminase,ACCD)的PGPB 通過代謝植物生產乙烯的前體ACC 來降低乙烯水平,還可以通過改變編碼乙烯合成酶、ACC 合酶和ACC 氧化酶基因來影響植物乙烯內源穩態,促進根伸長[15,26]。
近年來,大量研究證明,具有ACC 脫氨酶活性的菌株能夠在實際生產中改善植物的生長狀態。例如,熒光假單胞菌能夠使玉米在電導率為9 dS/m 的鹽堿土中根長增加330%,惡臭假單胞菌能夠使玉米在12 dS/m 的鹽堿土中株高增加230%[27]。巴西固氮螺菌FP2 可以使小麥植株ACC 氧化酶表達量降低,促進根的生長[28]。耐鹽PGPBMicrococcus yunnanensis、Planococcus rifietoensis和Variovorax paradoxus接種到甜菜(Beta vulgarisL.)后,提高了種子發芽率和植株生物量,增強了光合作用,脅迫誘導的乙烯含量也顯著降低[29]。以上研究表明,能夠產生ACC 脫氨酶的PGPB 可以減輕鹽脅迫對作物的影響,并將乙烯降低到抑制生長的水平以下。多種植物內生菌也被發現具有ACC 脫氨酶活性[30],通過對比從各種生境(包括植物的不同組織)中分離菌株的ACC 脫氨酶活性,發現內生菌比從根際土以及非根際土等生境分離的PGPB 具有更高的ACC脫氨酶的活力[31]。目前,具有ACC 脫氨酶活性的PGPB 已成為鹽土改良的重要生物資源。
2 PGPB 調節植物內源激素水平
2.1 PGPB產生IAA
通過對受到鹽和干旱脅迫的擬南芥和水稻等植物的多種信號傳導途徑和組分研究,發現植物激素互相協調配合可以提高植物滲透調節水平[32]。植物生長素(auxin,IAA)對植物細胞、組織和新生器官的特異性等有重要作用,作物在受到鹽脅迫后體內IAA 濃度顯著降低,如番茄可降低75%左右[33],IAA 信號轉導途徑與植物幼苗的發育過程關系緊密[34],鹽脅迫通過改變植物IAA 的積累和再分配重塑了根系結構,影響了植株的生長速度[35]。用IAA處理小麥種子可以減輕鹽脅迫對小麥種子萌發和幼苗發育造成的不利影響[36]。
很多PGPB 能夠產生IAA[37]。研究發現,小麥接種產生IAA 的中度嗜鹽菌后,根中IAA 含量顯著提高,其生長量顯著增加[38]。Sharma 等[39]將產生IAA的菌 株Klebsiella、Pseudomonas、Agrobacterium和Ochrobactrum接種到Arthrocnemum indicum后,植物生物量顯著增加,耐鹽性增強。從鹽角草根際分離的產IAA 耐鹽菌株B. endophyticus、B. tequilensis、Planococcus重新接種到鹽角草后,發芽率增加7%-11%,莖長增加13%-22%,根長增加44%-57%,鮮重增 加21%-54%[40]。在25 mmol/L NaCl鹽脅迫下,茄子幼苗接種產IAA 的菌株Xanthobacter autotrophicusBM13、Enterobacter aerogenesBM10 和Bacillus brevisFK2 后,幼枝鮮重較對照組顯著增加12.13%-19.58%,產量也得到提高[41]。以上研究表明,通過PGPB 調節鹽生植物和甜土植物的IAA 生成量可能是增強作物耐鹽性的重要因素。
2.2 PGPB產生ABA
脫落酸(Abscisic acid,ABA)是一種重要的應激激素,可在植物響應非生物脅迫后合成,并激活抗逆基因的表達[42],是調節植物全生育期活動以及刺激植物產生對廣泛非生物脅迫適應性反應的重要信號介質。ABA 作為內源信號分子能夠通過調節氣孔活動影響植物的光合作用,進而減輕鹽脅迫對植株的損傷并提高植物的耐鹽性和存活率[3,43-45]。研究發現,許多PGPB 能夠在鹽脅迫下產生ABA,改變植物內源ABA 水平,使RAB18、ABA 反應元件RD29A和RD29B調節子、脫水反應元件(DRE)等基因表達上調,提高植物耐鹽性[21,46-47]。
豌豆(Pisum sativum)接種產生ABA 的爭論貪噬菌5C-2 降低了根中ABA 的水平,并影響韌皮部中芽-根ABA 轉運的長距離信號傳導和木質部中的根-莖ABA 轉運過程[48]。葡萄與擬南芥接種產ABA 的地衣芽孢桿菌、熒光假單胞菌、巴西固氮螺菌后,內源ABA 含量增加并且具有更高的滲透脅迫耐受性[49-50]。大量研究表明,PGPB 誘導的植物ABA 水平變化可能會降低植物對滲透脅迫的敏感性,對提高植物非生物脅迫耐受性具有顯著作用。
2.3 PGPB產生GA
赤霉素(Gibberellin,GA)參與植物的多個生長發育階段,如種子萌發、葉片膨大、光形態建成、莖伸長、花器的發育和果實的成熟等,對細胞分裂和伸長,下胚軸、根和葉分生組織大小的調節有積極作用[51-53]。GA3是一種具有生物活性的GA,在低鹽脅迫下可以顯著提高種子的發芽率和植物的水分利用率[54-55],在高鹽脅迫下仍能顯著改善植物的生長狀況[56]。
PGPB 可以影響植物內源GA 水平[57]。例如,接種產生GA 的蠟狀芽孢桿菌 MJ-1 的辣椒,莖鮮重增加138%,根鮮重增加128%,具有更高的內源GA 水平,表現出更好的定殖和促生效果[58]。同樣,Leifsonia soliSE134 菌株產生的GA 對水稻、黃瓜、番茄和 蘿卜有促生作用[59]。Bacillussp. 和Azospirillumsp.產生的GA 增加了植物氮的吸收[60]。接種產GA3的解淀粉芽孢桿菌,水稻內源GA1、GA4、GA7和GA9被上調,水楊酸含量增加,促生效果顯著[61]。接種產生GA 的PGPB 能夠通過影響植物內源GA3、水楊酸、脫落酸等激素水平促進作物生長[62]。
2.4 PGPB產生JA和SA
茉莉酸(Jasmonic acid,JA)和水楊酸(Salicylic acid,SA)廣泛存在于植物體內[63]。JA 作為信號分子參與植物對水、鹽的響應,也是植物各發育過程和防御反應的重要信號分子[64]。近年來,分子生物學研究證實了JA 與植物耐鹽性的相關性。SA 在植物細胞膜疏水區積累,可以減少細胞內電解質外滲,減輕膜脂過氧化,保護蛋白質及膜結構的完整性,對植物種子萌發、離子吸收與轉運、細胞膜透性、光合作用及生長速率等生理生化過程產生影響,進而減輕鹽脅迫對植物的傷害[65-66]。
多種能夠合成并向植物提供JA、SA 的PGPB能夠促進植物生長,提高植物耐鹽性、抗病能力[67-68]。產生SA 的解淀粉芽孢桿菌LJ02 接種到黃瓜幼苗后,植株SOD、POX、CAT、PPO、PAL 顯著提高,葉片中游離SA 含量和抗病基因PR-1的表達明顯升高,植株抗病能力增 強[69]。Burkholderia phytofirmansPsJN 作用于葡萄后也使植株內源SA 增加,激活并上調了葡萄的防御基因[70]。研究表明,產生JA 和SA 的PGPB 促進了植物內源SA 的積累,同時能夠提高植物的抗病和抗氧化活性,對植物的耐鹽性產生了積極影響。
2.5 PGPB產生CTK
嘌呤類化合物參與根愈傷組織分化和芽形成的過程[71]。細胞分裂素(Cytokinins,CTK)主要調節植物細胞分裂、頂端優勢、葉綠體生物發生、養分調節、葉片衰老、管組織分化、光信號傳導發育、芽分化和花青素的產生等生命活動,參與植物對生物和非生物脅迫抗性的形成[72-73]。多種PGPB能夠產生CTK 并改變植物內源CTK 濃度[37,74],其中,AHK3、AHK2、CRE1/AHK4/WOL 三種受體能夠以CTK 依賴的方式促進調節因子的表達和信號傳 導[75-76],對促進植物生長有積極作用[77]。例如側柏接種產CTK的枯草芽孢桿菌,芽中CTK 含量增加,對滲透脅迫的抗性更強[78]。枯草芽孢桿菌接種到萵苣根際,通過調節植株根-芽CTK 信號傳導來促進作物生長[79]。PGPB 合成CTK 并改變植物CTK內穩態的能力凸顯了PGPB 在刺激植物生長和增加植物耐鹽性上的重要性。
3 PGPB 促進植物營養吸收
3.1 PGPB的溶磷作用
磷(P)是植物的主要常量營養素之一。土壤中P 含量約0.05%(W/W),其中只有0.1%的P 為有效磷,嚴重限制了P 的利用率[80]。鹽堿地降低了植物對磷的吸收,抑制了植物生長和代謝,而施用無機磷肥料及鹽水灌溉會增加土壤鹽分[16]。溶磷菌(Phosphate-solubilizing bacteria,PSB)能夠分泌低分子量有機酸,通過螯合、離子交換和酸化等各種機制將不溶性磷酸鹽轉化為HPO42-和H2PO4-,進而提高植物組織中磷含量,改善植物長勢并減輕鹽脅迫損傷[81]。
從白骨壤根際篩選鑒定出129 種能夠溶解磷酸鹽巖的細菌菌株,其中,Oceanobacillus picturae能夠產生多種有機酸以及酸性、堿性磷酸酶,能夠溶解97%磷酸鹽巖,增加木質部導管數量,促進根和莖養分吸收,增強光合氣體交換,顯著促進幼苗根和芽的生長[82]。鹽脅迫下,番茄接種Achromobacter piechaudii后植株磷含量和水分利用效率提高[83]。部 分植物內生菌,包括Arthrobacter、Bacillus和O. picturae等能夠溶解Ca3(PO4)2、AlPO4和FePO4,可增加植株高度、干重、側枝數量和穗大小[82,84-85]。慢生根瘤菌和假單胞菌通過改變大豆根系結構,促進了氮、磷的吸收,并提高了其耐鹽性[86]。綜上所述,在鹽脅迫環境中,PSB 的應用可促進植物磷素吸收而不會加劇土壤鹽度。
3.2 PGPB產生鐵載體
鐵(Fe)是植物生長所需的一種微量營養素,是植物呼吸、光合作用等反應中關鍵酶的組成部分[87]。鹽脅迫抑制了土壤中包括Fe 在內的大量微量營養素的可利用性,嚴重抑制了植物生長[88]。鐵載體是PGPB 分泌的一種水溶性、小分子物質,可以專屬、高親和力地與Fe3+結合成可被吸收的有機螯合物形式[89]。產鐵載體的PGPB 對鐵的親和力高于真菌病原體,因此,可以有效抑制病原體的增殖。鐵載體能夠增加鐵在根際的供給量,植物則可以利用鐵載體螯合的鐵來改善植物營養、防止植物缺鐵失綠癥的發生[90]。目前,已經報道的產鐵載體的PGPB 有B. thuringiensis、Gracilibacillus saliphilu、Arthrobacter pascens等,能夠在鹽漬土中促進Arthrocnemum、macrostachyum、Atriplex leucoclada等作物生長[91-92]。鹽堿土壤中鐵的可用性非常低[88],研究表明,接種產生鐵載體的PGPB 對提高植物的抗病性和鐵元素攝取量具有積極作用[93-94]。
3.3 PGPB的固氮作用
氮素(N)是陸地生態系統中最重要、最易匱乏的養分之一,土壤中缺乏有效氮會限制甜土植物及鹽生植物的生產力,鹽脅迫可以干擾植物氮代謝,抑制銨和硝酸鹽的吸收和同化[95],降低植物組織的N 含量[96]。過量使用無機肥將增加土壤鹽度、破壞土壤結構、改變土壤微生物的組成、干擾植物N 吸收,反而降低植物組織的N 含量[96-97]。有研究表明,在鹽土中施無機氮肥后,鹽度增加導致的危害多于氮素增加帶來的益處[98]。
固氮菌是鹽漬土中植物有效氮的重要來源,同時,固氮PGPB 可以通過產生滲透因子來維持植物正常的細胞膨壓和新陳代謝[99]。例如,假產堿假單胞菌接種到海蓬子后,植株葉綠素含量和N 含量顯著增加[100];固氮菌Klebsiella pneumoniae顯著促進鹽角草早期幼苗生長,發芽率、鮮重、干重和根長均顯著高于未接菌處理的植株[101];Jha 等[98]研究表明,與對照相比,鹽脅迫條件下S. brachiate接種B. saurashtrense和Pseudomonas后生物量顯著增加。可見,固氮PGPB 對植物及環境的潛在益處突出了其作為鹽土農業發展潛在生物資源的重要性。
4 PGPB 調節植物抗氧化酶活性
鹽脅迫會破壞植物體內ROS 產生與清除間的動態平衡,引起生物大分子的氧化損傷甚至細胞死亡[102]。過量的自由基一方面生成過量丙二醛等物質造成膜質過氧化,損害生物膜及其功能[103];另一方面造成植物體內負責光合色素合成的特異性酶活性下降,葉綠體基粒片層膨脹松散甚至片層解體,光合系統中的超微結構遭到破壞[104]。PGPB 可以通過上調關鍵酶的活性來激活植物抗氧化防御機制來防止過量ROS 導致的膜質變質,所述關鍵酶包括超氧化物歧化酶、抗壞血酸氧化酶、過氧化物酶、過氧化氫酶和谷胱甘肽還原酶,同時PGPB 也可以產生半胱氨酸、谷胱甘肽和抗壞血酸等非酶組分以清除過量的活性氧物質[105]。
5 PGPB 調節植物離子平衡
5.1 PGPB產生滲透調節因子
維持植物的滲透平衡對提高其抗鹽性具有重要作用。PGPB 能夠促進植物積累相容性溶質以維持細胞內滲透平衡,如氨基酸及其衍生物(如谷氨酸、脯氨酸、肽和N-乙酰化氨基酸等)、季胺(如甘氨酸甜菜堿和脯氨酸甜菜堿等)、糖(如葡萄糖苷、蔗糖和海藻糖等)、四氫嘧啶、羥基四氫嘧啶以及乙酰二氨基酸(乙酰鳥氨酸、乙酰賴氨酸)等[106-107],其表現出的多種與脅迫相關的特性可以使作物在高鹽度下受到保護[39]。其中,四氫嘧啶類物質是目前發現的在細菌界分布最廣泛的相容性溶質,兼具滲透壓調節、誘導滲透壓基因表達、抗逆保護、分子伴侶等諸多作用。在鹽脅迫下,脯氨酸通過穩定細胞膜和蛋白質等亞細胞結構,清除自由基并緩沖細胞氧化還原電勢來減輕植物鹽脅迫損傷[108]。甘氨酸甜菜堿(GB)和脯氨酸是植物中積累的兩種主要的抵抗鹽、干旱脅迫的有機滲透物,兩種化合物對酶和膜完整性具有積極作用[109]。蘿卜在鹽脅迫下接種PGPB 后,光合色素、脯氨酸、總游離氨基酸和可溶性蛋白含量增加,同時檢測到IAA、GA3、N、P、K+、Ca2+和Mg2+的含量均有提高[110]。Zarea 等[111]研究表明,PGPB 定殖于小麥根際后可通過增加植物脯氨酸積累促進植物生長。Upadhyay 等[112]發現PGPB 處理的小麥中脯氨酸和可溶性糖含量的增加是提高其耐鹽性的關鍵。在鹽脅迫下,滲透調節物質能夠保護生物大分子的結構和功能,防止酶失活,保障光合作用等生理活動進行,增加作物的鹽脅迫系統耐受性,對鹽土農業利用甚至未來農業發展具有十分重要的意義[113]。
5.2 PGPB產生胞外多糖
生物膜建立在植物根和土壤顆粒表面,對提高作物產量、改善土壤理化性質和減少病原體侵染植物根部有重要作用[114-115]。產生胞外多糖(Exopoly saccharides,EPS)的PGPB 能夠促進生物膜的形成,進而提高土壤肥力、保持種子周圍水分充足、促進植物養分吸收和減輕植物Na+損傷。生物膜和EPS 的增加有助于土壤顆粒與植物根系的聯系[114]。EPS 中羥基、巰基、羧基和磷酰基官能團的結合特性會促進土壤顆粒粘結并加強團聚體的形成,提高植物在鹽脅迫下生物量積累[116]。目前,報道較多的產EPS 的耐鹽菌株主要有Aeromonas、hydrophila、caviae、Bacillussp.、Planococcus rifietoensis、Halomonas variabilis、Burkholderia、Enterobacter、Microbacterium和Paenibacillus等[117-120]。
5.3 PGPB參與轉運蛋白調控
水通道蛋白是存在于植物細胞的等離子體和細胞內膜中的蛋白,作為水的主要轉運蛋白,使植物繼續從高鹽度土壤中吸收水分,將有助于植物應對干旱和鹽脅迫[121],鹽脅迫條件下,玉米和豌豆等接種變形菌和巨大芽孢桿菌后,根部PIP2和ZmPIP1-1上調,表明接種PGPB 有助于提高植物的根際導水率[14]。
高親和K+轉運載體(HKT)是與植物耐鹽性有關的一類Na+或K+轉運體或Na+-K+共轉運體[122]。研究表明,枯草芽孢桿菌可以通過下調植物根中HKT基因的表達來減少根對Na+的過量吸收,促進芽- 根Na+再循 環[14,123]。Islam 等[13]的研究 表明,鹽脅迫下接種PGPB 可能通過調節離子轉運蛋白的表達來提高植物吸收選擇離子的效率以保持高K+/Na+比,減少Na+和Cl-等離子的積累,調節大量營養素和微量營養素的平衡。
6 PGPB 誘導植物系統抗性
鹽脅迫引起的滲透脅迫、離子毒害、氧化脅迫和植物病害等復雜因素能夠破壞植物細胞、組織和器官,引起植物生理生化代謝紊亂,甚至造成植物死亡(圖1)。誘導系統抗性(Induced system tolerance,IST)即微生物在植物根際或內部定殖后,植物形成的抵抗廣泛的生物與非生物因子的防御能力[124]。研究表明,微生物及其組分,如脂多糖、鞭毛、鐵載體、環脂肽、2,4-二乙酰基間苯三酚、高絲氨酸內酯和某些揮發物作為誘導系統抗性的誘導劑保障了植物的健康生長和發育[125],使其能夠抵抗多種細菌、真菌和病毒性植物病害[126]。
揮發性有機化合物(Volatile organic compounds,VOCs)是分子量較低、疏水性、常溫常壓下容易揮發的化合物,可在大氣和土壤中擴散[127]。微生物產生的VOCs 和N-acyl-L-homoserine lactones(AHLs)等多種信號在植物形態發生過程中起重要作用[20]。Pseudomonas產生的VOCs 可以顯著上調營養貯藏蛋白、γ-谷氨酰水解酶和RuBisCo大蛋白,是在鹽脅迫下誘導大豆產生系統耐受性的關鍵[128]。Alcaligenes faecalisJBCS1294 通過散發己二酸和丁酸,改變生長素和赤霉素合成途徑來提高植物耐鹽性[23]。Paraburkholderia phytofirmansPsJN 產生的揮發性有機物2-十一酮、7-己醇、3-甲基丁醇和二甲基二硫等也被驗證具有促進擬南芥生長并誘導其產生脅迫耐受性的作用[129]。
HCN、吩嗪、硝吡咯菌素、2,4-二乙酰基間苯三酚、藤黃綠膿菌素和張力蛋白等是PGPB 能夠產生的抗真菌代謝物[23]。PGPB 還能夠產生真菌細胞壁降解酶,如能降解一些真菌細胞壁脂類的脂肪酶、降解真菌細胞壁碳水化合物的β-1,3-葡聚糖酶、降解真菌細胞壁幾丁質的幾丁質酶、降解細胞壁蛋白的蛋白酶等[130-131]。Egamberdieva 等[132]報道,當用Pseudomonas aeruginosa處理鹽堿土中黃瓜和番茄時,可抑制Fusarium solani引起的根腐病。研究證實,即使在鹽脅迫下,PGPB 仍然能夠介導植物產生抵抗多種植物病害的系統抗性[133]。
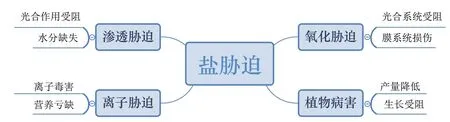
圖1 鹽脅迫對植物的影響
基于鹽漬化土壤的理化性質和鹽脅迫下植物的受損機制,結合上述研究成果,我們認為,PGPB 能夠從多方面通過多種途徑改良土壤,依靠不同特性提高植物滲透調節能力、離子轉運能力、激素調節能力、抗氧化能力和養分吸收能力(圖2)。
7 結論與展望
鹽脅迫使植物同時面臨離子脅迫、滲透脅迫、氧化脅迫和植物病害等威脅,PGPB 的多種特性能夠協助植物減輕脅迫損傷,其中,調節植物內源激素水平、促進營養吸收等特性對植物生長具有重要促進作用。本課題組依托于黃河下游森林培育國家林業和草原局重點實驗室,在研究中證實,耐鹽菌株能夠在鹽脅迫環境中保持更高的生物活性,顯著提高小麥和棗樹等作物的滲透調節、抗氧化和養分吸收能力,具有改變根系結構,提高生物量積累的作用效果。為了進一步探究植物-土壤-PGPB 的作用關系、篩選得到更多具有實效的PGPB 菌株、加快高效耐鹽PGPB 的產業化應用、減少鹽漬化土壤快速擴張帶來的生產損失,建議在以下方面做進一步研究:
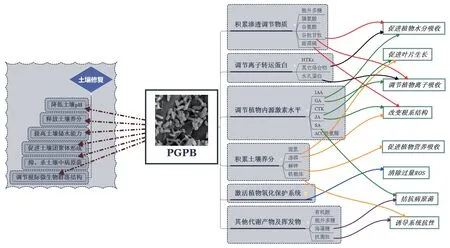
圖2 鹽脅迫環境下PGPB 對土壤與植物的作用機制
(1)植物激素是植物用于調節各種應激反應的必要因子,分子生物學的突破增加了對植物激素信號通路的認識。鹽脅迫使植物產生多種信號傳導途徑,而IAA、GA、ABA、CTK、JA 和SA 等在多種信號傳導中存在相互促進或抑制的情況[113],不同激素在植株內的協同作用仍有待闡明。此方向的研究有利于更全面地認識PGPB 對植物激素的調控機制,可為最終獲得可靠的PGPB 菌種提供理論依據。
(2)目前,關于鹽脅迫下PGPB 的研究大多集中在其對宿主植物的促生效應,而PGPB 與根際其他微生物對鹽漬土的協同作用的研究則鮮有涉及。同時,PGPB 能夠對植物的根際土、甚至內生菌微生物群落結構、多樣性、穩定性產生影響,通過高通量測序等技術手段分析PGPB 對其作用規律,找到群落的發生、演替規律,發現穩定存在于根際和作物內部的有益菌群,分析優勢菌群的功能特性,了解其對植物生長、耐鹽性的影響,將為更好地揭示PGPB 在鹽漬土改良中的生態學功能提供理論依據。
(3)PGPB 對環境條件存在依賴性,部分菌株在脅迫環境中活性降低甚至失活。研究發現,從鹽、堿、酸、干旱、水淹、高溫及低溫等環境分離的PGPB 可以對相應農業條件下的作物提供有效保護,因此,鹽生植物與鹽土中PGPB 資源的開發將加快菌劑篩選進程,另外,通過基因工程手段構建生物活力高、試驗效果好、活性穩定的菌株也是重要的獲得方式。
(4)不同PGPB 在不同鹽分類型、不同脅迫強度和不同脅迫時間的土壤中定殖能力不同,對不同植物及同一植物不同部位的影響也存在差異。很難用一個標準來界定PGPB 對植物的促生效果。若能針對不同鹽漬土壤類型、植物種類和組織部位確定適當的評價標準,將更有利于評價PGPB 對植物和土壤的影響,也將為篩選高效耐鹽的PGPB 菌種及其應用提供理論和技術依據。
(5)獲得某種具有特定功能的菌株并非難事,但獲得生物活力高、可在植物根際或組織內部穩定定殖、發酵性狀良好、生物安全性符合要求、分類地位清晰、作用機理明確、大田試驗效果好的菌株仍需大量的研究工作。目前,多數研究工作仍局限在人工模擬條件,作物培養方式多為短周期水培、盆栽等,而對在自然生境下土壤與植物間的營養分配和水鹽傳導規律的認識仍存在不足,植物-土壤-PGPB 的作用方式、信號傳遞途徑、干擾因素等研究尚不完善,菌劑在自然環境中最佳的施用條件、適用作物、用法用量等仍需完整的試驗驗證,該工作的研究要求增加在自然環境和生態系統尺度上的試驗。
(6)鹽堿土改良對改善植物長勢至關重要,綜合利用物理、化學、水利、生物等土壤改良措施具有重要意義。目前,通過小規模研究發現PGPB 與其他改良劑聯合使用能夠更好地改善植物長勢,增加土壤養分,提高改良水平。科學地結合不同改良劑并創新試驗方法對提高鹽土改良效果、減少修復成本、維持生態系統穩態具有更大的優勢和更高的推廣價值。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13