不同種屬酵母菌共培發(fā)酵桑葚酒的工藝優(yōu)化
2020-05-11 02:00:22葉片劉建黃均嚴(yán)樂晉周榮清
食品與發(fā)酵工業(yè) 2020年8期
關(guān)鍵詞:酵母菌
葉片,劉建,黃均,嚴(yán)樂晉,周榮清,2*
1(四川大學(xué) 輕工科學(xué)與工程學(xué)院,四川 成都,610065)2(制革清潔技術(shù)國家工程實(shí)驗(yàn)室,四川 成都,610065)
桑葚是桑科植物桑樹的果實(shí),富含多糖和白藜蘆醇等功能組分,有增強(qiáng)人體免疫力、降血脂血糖和防癌抗變等作用[1]。桑葚采摘期僅在每年的4~6月,且不耐壓、易腐爛,不宜在室溫下儲(chǔ)存和運(yùn)輸。除以鮮食外,深加工主要是生產(chǎn)濃縮糖漿、果醬、醋和果酒等[2],其中生產(chǎn)桑葚酒是提高附加值的重要途徑之一[3]。目前桑葚酒技術(shù)開發(fā)關(guān)注的是原料加工性能、菌株篩選和特征及功能組分的剖析等[4-6]。
酵母菌的性質(zhì)對(duì)果酒風(fēng)味特征影響顯著,基于釀酒酵母菌和非釀酒酵母菌的共培發(fā)酵技術(shù)不僅可改善果酒的風(fēng)味特征,還使之高度發(fā)酵,是生產(chǎn)高品質(zhì)果酒常用的技術(shù)之一[7-8]。Torulasporadelbrueckii具有產(chǎn)異戊醇、苯乙醇、乙酯類和萜烯等物質(zhì)并賦予其風(fēng)味特色組分的能力[9-10],可采用其與Saccharomyces酵母菌共培養(yǎng)發(fā)酵方式改善果酒的品質(zhì)。
本文報(bào)道了不同類型桑葚原料、預(yù)處理方式對(duì)桑葚酒品質(zhì)及風(fēng)格影響的研究結(jié)果。同時(shí)探討了酵母屬的S.cerevisiae和S.bayanus分別與T.delbrueckii共培及不同溫度發(fā)酵對(duì)桑葚酒風(fēng)味特征的影響,旨在開發(fā)生產(chǎn)高質(zhì)量桑葚酒的生產(chǎn)技術(shù)。
1 材料與方法
1.1 材料與試劑
1.1.1 材料
新鮮桑葚果,購于本地農(nóng)貿(mào)市場(chǎng);冷凍桑葚,成都世煌生物科技有限責(zé)任公司饋贈(zèng)。白砂糖(一級(jí)),購于本地副食品商店。
S.cerevisiaeY1(CCTCC M 2019521)、S.bayanusY4(CCTCC M 2019522)、T.delbrueckiiY7 (CCTCC M 2019523),本實(shí)驗(yàn)室從果酒廠的發(fā)酵醪中分離,經(jīng)菌落及菌體細(xì)胞形態(tài)觀察、生理生化試驗(yàn)結(jié)果[11]初步鑒定和ITS測(cè)序確認(rèn),送中國典型培養(yǎng)物保藏中心保藏。
1.1.2 試劑
標(biāo)品草酸、檸檬酸、酒石酸、L-蘋果酸、乳酸、乙酸和丙酸,Sigma(CP,上海,中國);Lallzyme EX-V復(fù)配果膠酶制劑P1,康禧食品飲業(yè)有限公司(上海,中國)。
1.1.3 儀器與設(shè)備
Trace GC Ultra-DSQⅡ氣相色譜質(zhì)譜聯(lián)用儀,Thermo Fisher Electron;HP-INNOWAX毛細(xì)管色譜柱(30 m×0.25 mm×0.25 μm)、1260高效液相色譜儀,美國Aglient;Alltech OA-1000有機(jī)酸色譜柱(300 mm×6.5 mm),美國GRACE;SPE小柱,成都SWELL;CAR/PDMS/DVB萃取頭(50/30 μm),美國Supelco。
1.2 試驗(yàn)方法
桑葚破碎后,先加入NaHSO3(120 mg/L),隨即加入果膠酶(70 mg/L),室溫靜置10 h。然后添加(NH4)2HPO3(150 mg/L),加蔗糖調(diào)整至23.5 Brix。
原料的影響:S.bayanusY4的菌懸液接種到預(yù)處理后的原料中,使其菌體初始濃度為5×106CFU/mL,17 ℃下發(fā)酵12 d,取發(fā)酵清液分析。這些原料包括鮮全果(FG)、冷凍全果(RG1,貯存1年)、冷凍全果汁(RZ)和冷凍全果(RG2,貯存2個(gè)月)等4種不同類型。
不同種屬酵母共培養(yǎng)的影響:預(yù)處理的鮮桑葚全果汁分為6份,其中4份的操作步驟為分別接種S.cerevisiae和T.delbrueckii菌懸液(1∶1,5×106CFU/mL,Y1&Y7);S.bayanus和T.delbrueckii菌懸液(1∶1,5×106CFU/mL,Y4&Y7),接種S.cerevisiae(Y1)和S.bayanus(Y4)為對(duì)照,菌體初始濃度和過程參數(shù)同前。置于(25±2) ℃條件下,發(fā)酵6 d時(shí),補(bǔ)糖4%調(diào)整糖度,繼續(xù)發(fā)酵4 d,過濾得原酒液。
溫度的影響:剩余2份操作步驟分別與Y1&Y7和Y4&Y7相同,僅發(fā)酵的溫度控制在(20±2)℃以探討溫度影響。樣品簡(jiǎn)稱為LY1&Y7和LY4&Y7。
1.3 分析檢測(cè)方法
1.3.1 理化指標(biāo)的測(cè)定
原料及原酒的主要理化指標(biāo)按照GB/T15038—2006所述的方法檢測(cè)。
1.3.2 有機(jī)酸的測(cè)定
按照參考文獻(xiàn)[12]所述方法,應(yīng)用HPLC檢測(cè)。主要操作步驟為3 mL離心上清液加載到活化的SPE小柱,洗脫液經(jīng)0.22 μm濾膜過濾后,高效液相色譜法檢測(cè),檢測(cè)波長為210 nm。以檸檬酸,酒石酸,L-蘋果酸,琥珀酸,乳酸和乙酸標(biāo)準(zhǔn)品的保留時(shí)間鑒定樣品中有機(jī)酸,并采用外標(biāo)法定量。
1.3.3 揮發(fā)性組分的測(cè)定及分析
按照參考文獻(xiàn)[13]所述應(yīng)用頂空固相微萃取-氣相質(zhì)譜方法檢測(cè)。主要操作步驟為精確取0.5 mL樣品置于頂空瓶中,用NaCl飽和,加入混合內(nèi)標(biāo)2-辛醇(69 mg/L,GC級(jí))和辛酸甲酯(71 mg/L,GC級(jí)) 10 μL,60 ℃平衡15 min,插入萃取頭,萃取40 min, 然后GC進(jìn)樣口中解吸析3 min。通過待測(cè)樣品組分質(zhì)譜數(shù)據(jù)與標(biāo)準(zhǔn)譜庫(NIST2005)比對(duì),匹配度>800(最大為1 000)的物質(zhì)予以報(bào)道,采用內(nèi)標(biāo)法進(jìn)行半定量計(jì)算。
1.4 數(shù)據(jù)處理
每個(gè)樣品3次平行樣品檢測(cè)結(jié)果,采用SPSS 19.0軟件對(duì)測(cè)得各項(xiàng)指標(biāo)分析其單因素方差(Dancan’s test,P<0.05)。
2 結(jié)果與分析
2.1 原料類型及預(yù)處理方式對(duì)理化性質(zhì)的影響
如表1所示,冷凍桑葚因其內(nèi)源酶或原料表面棲息的微生物的作用使有機(jī)酸、總酸及揮發(fā)酸較其余類型的原料高,且酸類的含量也與貯存周期有關(guān),如RZ的總酸和揮發(fā)酸含量高于RG2的。各樣品間的殘?zhí)呛鸵掖紳舛葻o顯著區(qū)別。
如表2所示,琥珀酸、蘋果酸和草酸等是原料中優(yōu)勢(shì)有機(jī)酸[14]。冷凍貯存過程中因物理除酸作用,尤其是酒酒球菌的作用[15-16]誘導(dǎo)了蘋果酸—乳酸發(fā)酵。檸檬酸和酒石酸的含量降低了,從而導(dǎo)致酒體失去溫和爽快和后苦時(shí)間短的新鮮口感[17]。蘋果酸部分被轉(zhuǎn)換為琥珀酸使RG2中琥珀酸含量顯著高于其他樣品,賦予了原酒鮮味和酸味[18],而蘋果酸含量降低則減弱了果酒的酸澀及粗糙感。此外乳酸含量適量提高使之有更柔和的口感[5],但乳酸及乙酸含量偏高,不僅酸度高,還致使果酒出現(xiàn)酸刺激味等諸多欠愉悅的缺陷,冷凍或濃縮的桑葚不適合做釀酒原料。
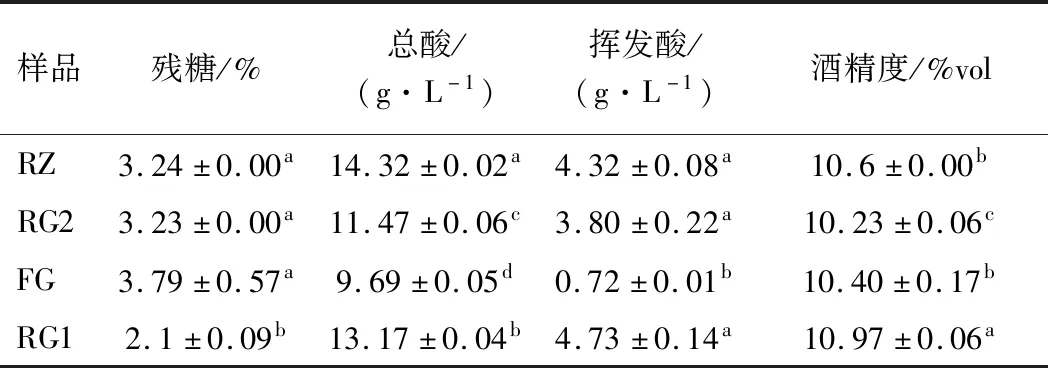
表1 不同原料及預(yù)處理對(duì)桑葚酒主要理化性質(zhì)的影響Table 1 Effect of different raw material and pretreatment pattern on major physicochemical properties
注:表中不同小寫字母表示差異顯著(P<0.05)(下同)

表2 不同原料及預(yù)處理對(duì)桑葚酒有機(jī)酸含量的影響 單位:g/LTable 2 Effect of different materials and pretreatment pattern on organic acid contents
2.2 不同類型酵母共培的桑葚果酒生產(chǎn)工藝
2.2.1 共培發(fā)酵溫度對(duì)理化指標(biāo)的影響
S.cerevisiae常用于新世紀(jì)葡萄酒的釀造[19]。在其過程中接種S.bayanus,可提高其甘油、乳酸和琥珀酸含量,使乙酸和乙醇的含量略低[20]。如表3所示,二者用于鮮全桑葚汁釀酒時(shí),乙醇、殘?zhí)羌翱偹岷坎]有顯著差異,僅降低發(fā)酵溫度時(shí),殘?zhí)呛柯愿叨掖紳舛嚷缘汀?/p>
2.2.2 共培養(yǎng)發(fā)酵對(duì)揮發(fā)組分的影響
同系Saccharomyces的S.cerevisiae和S.bayanus所釀桑葚原酒的主要理化指標(biāo)相同,但揮發(fā)性組分含量差異顯著。這2株酵母菌分別與T.delbrueckii共培時(shí),對(duì)揮發(fā)性組分的貢獻(xiàn)因種屬不同而異。S.cerevisiae與T.delbrueckii共培,揮發(fā)性組分總量略增,其中酚和酮類的含量顯著提高,但酯和酸類含量降低,比例或增或減。S.bayanus與T.delbrueckii共培,其總量及各組分比例均略增高。
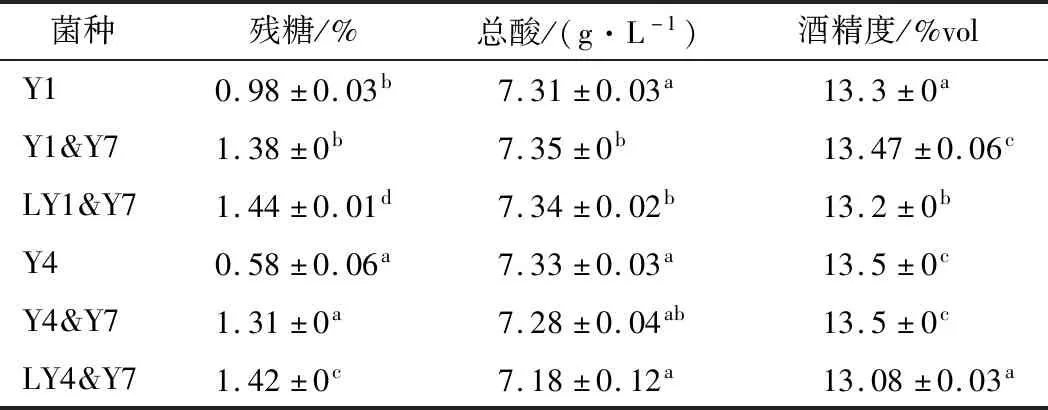
表3 酵母菌間共培及溫度對(duì)主要理化指標(biāo)的影響Table 3 Effect of co-culture by different yeast strain and fermentation temperature on major physicochemical properties of mulberry wine
如圖1所示,6種酒樣中共檢出了73種揮發(fā)性組分,包括酯(32)、醇(10)、酸(4)、醛(6)、酮(1)、酚(6)、萜烯類(7)和其他(7)等不同類型。Y4樣品55種,Y1樣品54種,分別與T.delbrueckii共培,使組分?jǐn)?shù)量增多。Y1&Y7和 Y4&Y7分別檢出了64種和59種,降低發(fā)酵溫度,前者減少了1種,而后者增加了6種。樣品間含量差異顯著的是酯、酚和醇3類組分,這些組分占總揮發(fā)組分總含量的92.95%~95.73%。
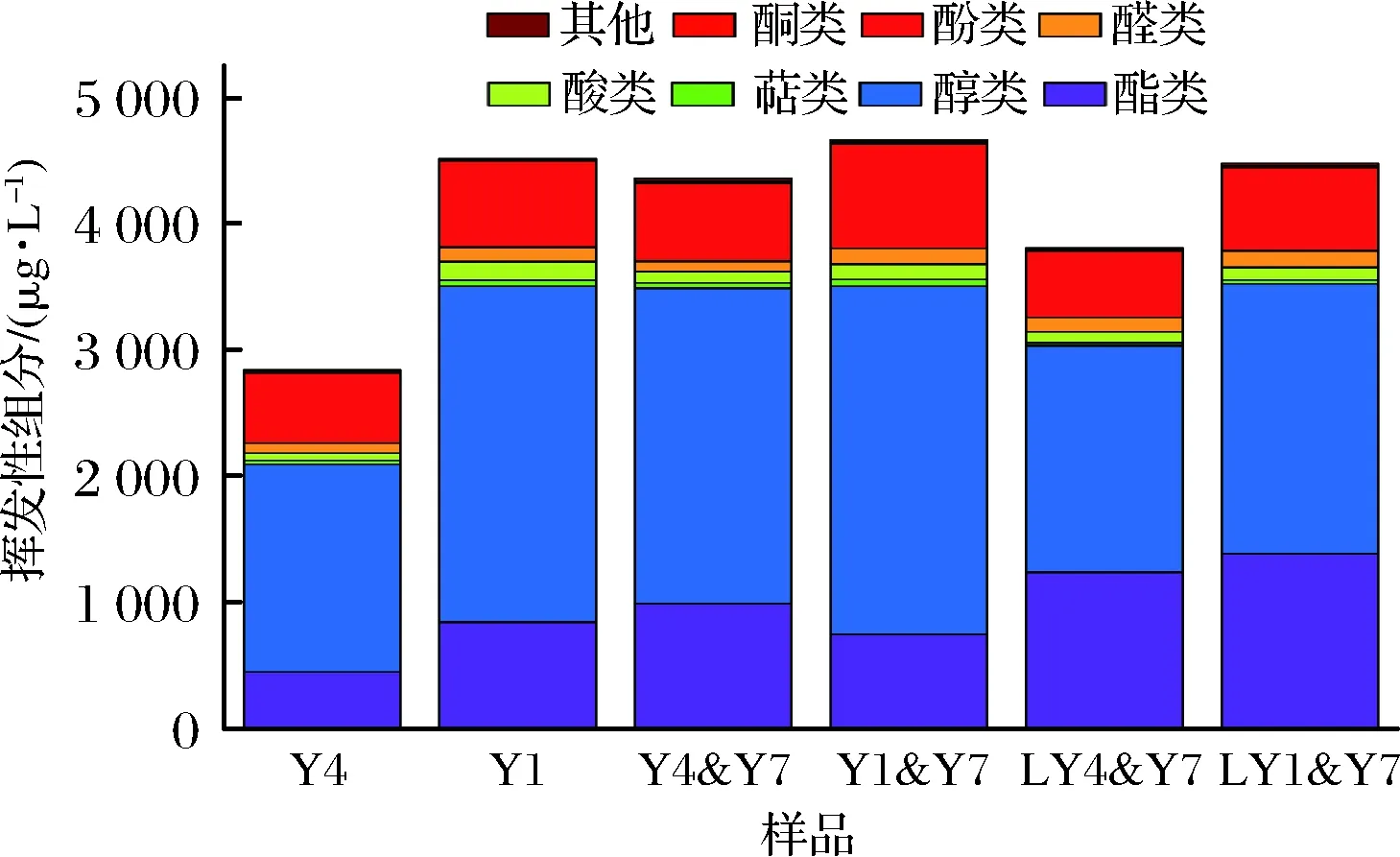
圖1 不同酵母菌共培及發(fā)酵溫度的桑葚酒揮發(fā)組分輪廓的差異Fig.1 Difference of volatiles profile among mulberry wine brewed by different co-cultured yeast and fermentation temperature
此外,這2株酵母菌分別與T.delbrueckii共培,也使桑葚酒的多種組分含量差異顯著。T.delbrueckii的貢獻(xiàn)主要是提高脂肪酸乙酯的含量,如辛酸乙酯和十二烷酸乙酯[21-22]。共培養(yǎng)增加了乳酸、辛酸、己酸和苯甲酸的乙酯類衍生物、十二烷酸甲酯和丁二酸二乙酯6種組分的含量,使己酸丁酯等4種組分的含量略降,且新檢出了肉桂酸乙酯和十四酸乙酯(圖2-a)。S.bayanus和S.cerevisiae因代謝途徑及生理生化性質(zhì)不同,與T.delbrueckii組合共培使己酸乙酯等8種酯類組分含量的增減幅度不同(圖2-b)。S.bayanus與T.delbrueckii共培,酯類組分略增,而S.cerevisiae與T.delbrueckii共培,酯類組分略降,新檢出了苯甲酸乙酯、磷酸三丁酯和丙位癸內(nèi)酯3種組分。共培發(fā)酵使異戊醇、1-己醇、異丁醇和十一醇4種醇類組分的含量增高,苯甲醇和辛醇含量降低(圖2-d),可能與T.delbrueckii中β-葡萄糖苷酶的活性有關(guān)[23]。1-癸醇、3-甲硫基丙醇和正庚醇等3種組分僅在S.cerevisiae和T.delbrueckii共培養(yǎng)時(shí)才被檢出。T.delbrueckii合成芳樟醇和α-萜品醇的能力較強(qiáng),橙花醇是芳樟醇的重要前體物之一[10],所以前者的含量減少,而后者略增。此外,萜類及除癸酸外的酸類組分含量略增(圖2-e)。
溫度影響T.delbrueckii的代謝速率,低溫降低了其合成苯乙醇的能力,使醇類組分的含量減少[24]。己酸、辛酸、十二烷酸、苯乙酸和肉桂酸的乙酯衍生物和4-羥基丁酸乙酰酯的含量減少,而甲酸辛酯、丁二酸二乙酯和戊二酸二乙酯的含量較高(圖2-c)。T.delbrueckii與S.bayanus共培使苯甲酸、苯乙酸、苯丙酸、軟脂酸的乙酯衍生物的含量增高,與S.cerevisiae共培則使這些組分的含量減少。
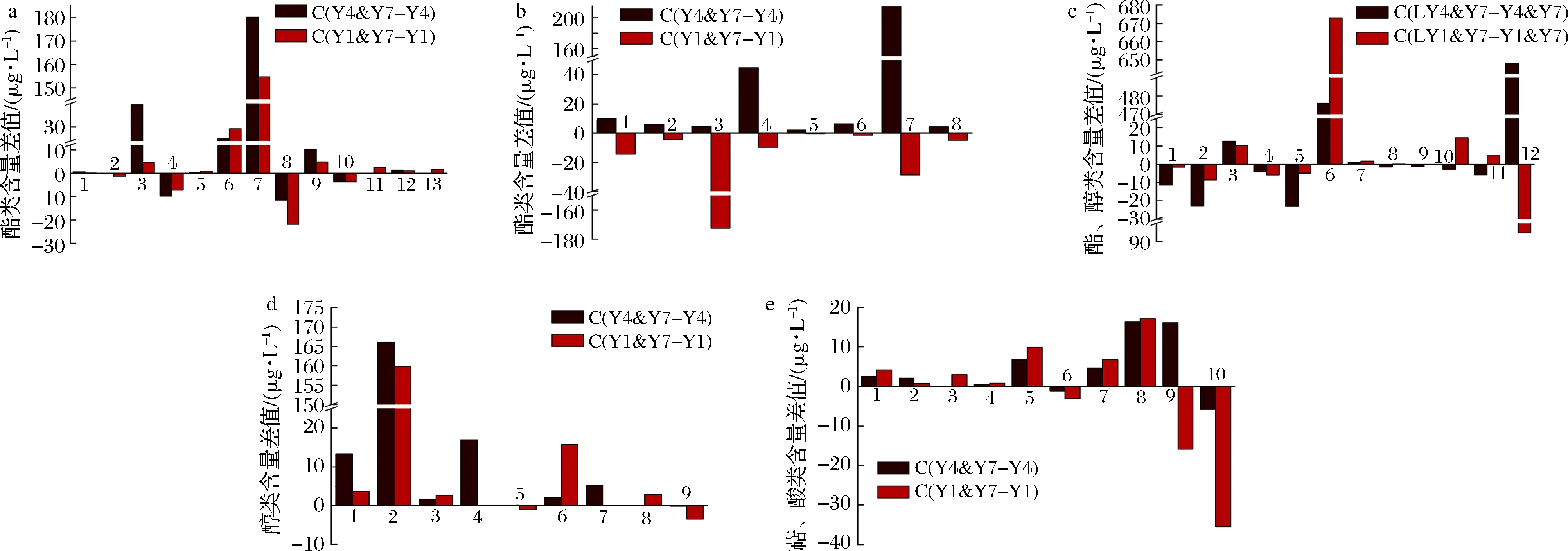
a-共培前后變化趨勢(shì)相似的酯類物質(zhì)的差異;b-共培前后變化相反的酯類物質(zhì)的差異;c-不同溫度共培酯類和醇類物質(zhì)的差異;d-共培前后醇類物質(zhì)的差異;e-共培前后萜類和酸類物質(zhì)的差異圖2 不同酵母菌共培及發(fā)酵溫度的桑葚酒主要揮發(fā)組分之間的差異Fig.2 Difference of major volatile among mulberries wine brewed by different co-cultured yeast and fermentation temperature注:a:1-乳酸乙酯;2-己酸丁酯;3-辛酸乙酯;4-甲酸辛酯;5-十二烷酸甲酯;6-苯甲酸乙酯;7-丁二酸二乙酯;8-乙酸苯乙酯;9十四酸乙酯;10-磷酸二乙基壬酯;11-磷酸三丁酯;12-肉桂酸乙酯;13-丙位癸內(nèi)酯;b:1-己酸乙酯;2-苯甲酰甲酸酯;3-4-羥基丁酸乙酰酯;4-十二烷酸乙酯;5-苯乙酸乙酯;6-苯丙酸乙酯;7-軟脂酸乙酯;8-十八烷酸乙酯;c:1-己酸乙酯;2-辛酸乙酯;3-甲酸辛酯;4-4-羥基丁酸乙酰酯;5-十二烷酸乙酯;6-丁二酸二乙酯;7-戊二酸二乙酯;8-苯乙酸乙酯;9-肉桂酸乙酯;10-苯甲酸乙酯;11-苯丙酸乙酯;12-苯乙醇;13-丙位葵內(nèi)酯;d:1-異丁醇;2-異戊醇;3-1-己醇;4-正庚醇;5-1-辛醇;6-十一醇;7-3-甲硫基丙醇;8-1-癸醇;9-苯甲醇;e:1-芳樟醇;2-(-)-4-萜品醇;3-松油醇;4-香茅醇;5-(+)-3-蒈烯,10-(乙酰基甲基);6-反式-橙花叔醇;7-2-甲基乙酸;8-己酸;9-辛酸;10-癸酸
對(duì)桑葚酒風(fēng)味貢獻(xiàn)度大的7種揮發(fā)組分(OAVs>1)的輪廓圖如圖3所示。癸醛氣味活度值(odor activity value,OAV)值最高,S.bayanus與T.delbrueckii共培養(yǎng)使之顯著降低,顯著減弱了酒樣中青草味、桔皮味和不愉快的脂肪味。S.cerevisiae與T.delbrueckii共培養(yǎng)使己酸乙酯、苯丙酸乙酯和苯乙醇升高,多種愉悅的芳香的強(qiáng)度增大。(20±2) ℃發(fā)酵時(shí),LY4&Y7中己酸乙酯、苯丙酸乙酯和苯乙醇的OAV降低,癸醛、癸酸和苯乙醛的OAV上升,且未檢出肉桂酸乙酯。除苯丙酸乙酯的OAV顯著上升和癸醛、苯乙醛的OAV略微下降外,LY1&Y7中呈現(xiàn)了類似的結(jié)果。S.bayanusy和S.cerevisiae分別與T.delbrueckii共培,降低溫度共培發(fā)酵,對(duì)果香、花香等果酒的特色芳香呈負(fù)貢獻(xiàn),使樣品中不愉悅的脂肪味和腐敗味及青草氣味增濃。

圖3 不同溫度及酵母菌共培樣品的OAVs輪廓Fig.3 OAVs profiles of mulberry wine samples with different co-cultured yeast and co-cultured temperature注:揮發(fā)性物質(zhì)感覺閾值參考文獻(xiàn)[25-27]
2.2.3 共培發(fā)酵對(duì)有機(jī)酸的影響
T.delbrueckii是與S.cerevisiae共培改善葡萄酒感官特征的非釀酒酵母之一,能有效提高葡萄酒的感官特征,發(fā)酵高糖度葡萄汁,降低乙酸及揮發(fā)性酸含量[28]。如圖4所示,Y1和Y1&Y7聚為一簇,而Y4和Y4&Y7聚為一簇,Saccharomyces酵母菌影響桑葚酒的有機(jī)酸含量,且S.cerevisia的原酒8種有機(jī)酸的含量較S.bayanus的略低。S.cerevisiae和S.bayanus分別與T.delbrueckii共培,乙酸含量略降。S.bayanus與T.delbrueckii共培,檸檬酸含量略降;S.cerevisiae與T.delbrueckii共培,琥珀酸含量略低,其他有機(jī)酸無顯著差異。

圖4 基于8種主要的有機(jī)酸不同酵母共培桑葚酒樣品的熱圖分析Fig.4 Heatmap analysis based on the 8 major organic acids of mulberry wine samples with different co-cultured yeast
3 結(jié)論
桑葚原料的類型、預(yù)處理方式、不同種屬的酵母菌共培養(yǎng)發(fā)酵及溫度顯著影響桑葚酒的品質(zhì),尤其是有機(jī)酸和關(guān)鍵揮發(fā)組分。冷凍后的桑葚用于釀酒,揮發(fā)酸和乳酸含量增高,不適合釀酒。鮮果破碎直接釀酒,揮發(fā)酸含量低,酸澀及粗糙感減弱。S.cerevisiae和S.bayanus分別與T.delbrueckii共培養(yǎng),改變了檢出的8種有機(jī)酸的比例,乙酸含量減少,也使檢出的揮發(fā)組分?jǐn)?shù)量增多,含量和比例與相應(yīng)單一種屬菌株不同,致使果酒的果香、花香等特色芳香及特征風(fēng)味增強(qiáng)。發(fā)酵溫度降至(20±2) ℃則使不愉悅氣味強(qiáng)度增大。
猜你喜歡
發(fā)明與創(chuàng)新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學(xué)少年(2021年1期)2021-02-04 08:03:45
當(dāng)代水產(chǎn)(2019年3期)2019-05-14 05:43:24
天然產(chǎn)物研究與開發(fā)(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學(xué)與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學(xué)低年級(jí)版)(2015年8期)2015-05-30 10:48:04
食品工業(yè)科技(2014年9期)2014-03-11 18:15:31
